
V.P. Gourineni
Nutritional Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama A and M University, Normal, AL, USA
M. Verghese
Nutritional Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama A and M University, Normal, AL, USA
J. Boateng
Nutritional Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama A and M University, Normal, AL, USA
International Journal of Cancer Research
Year: 2010 | Volume: 6 | Issue: 4 | Page No.: 220-233







ABSTRACT
Protective role of prebiotics such as inulin type fructans (Synergy 1®) in colon cancer prevention was demonstrated using animal models. In vitro and in vivo studies have shown the inhibitory effects of dietary fiber and isoflavones such as genistein against the development of cancer. However, the underlying mechanisms of these phytonutrients remain elusive. The aim of this study was to examine the anticancer effects of Synergy1® and soybean extracts in combinations on colon cancer cell line (Caco-2). Cells were incubated (12 and 24 h) with specific concentrations of Syn1®, SM, Syn1®+SM (25-200 μg mL-1), isoflavones (0.125-0.8 μg mL-1) and butyric acid (0.1-0.8 μg mL-1). Lactate dehydrogenase (LDH) release (%) was higher in cells at all concentrations following 24 h incubation with a mixture of Syn1® + SM compared to the cells treated with the extracts alone. LDH release (%) was increased by two fold in cells after incubation with genistein for 24 h compared to the LDH release after 12 h. Caspase-3 activity; a measure of apoptosis seems to be proportional to incubation time in cells treated with Syn1® and SM extracts. Histone associated DNA fragmentation and release of oligonucleotides was higher in cells incubated with a mixture of Syn1®+SM and butyric acid extracts. These results suggests anti-neoplastic properties of Synergy1® and Soybean in combinations by inducing cell toxicity and apoptosis (Caspase-3 and histone associated DNA fragmentation) as a result of synergism between bioactive components present in these functional ingredients. Phytochemical combinations tested at lowest and safer levels in this study may be effective in colon cancer prevention.
PDF Abstract XML References Citation
Received: April 17, 2010;
Accepted: September 28, 2010;
Published: November 04, 2010
How to cite this article
V.P. Gourineni, M. Verghese and J. Boateng, 2010. Anticancer Effects of Prebiotics Synergy1® and Soybean Extracts: Possible Synergistic Mechanisms in Caco-2 Cells. International Journal of Cancer Research, 6: 220-233.
DOI: 10.3923/ijcr.2010.220.233
URL: https://scialert.net/abstract/?doi=ijcr.2010.220.233
DOI: 10.3923/ijcr.2010.220.233
URL: https://scialert.net/abstract/?doi=ijcr.2010.220.233
INTRODUCTION
Cancer deaths worldwide are projected to rise with an estimated 12 million deaths by 2030 (WHO, 2009). One cancer that is most commonly diagnosed in populations of economically developed nations is colon cancer, the third most common cause of mortalities among men and women in the US accounting for 25,240 estimated deaths in the present year (ACS, 2009). Previous research reported a relationship between dietary consumption patterns and prevention of chronic diseases (Potter, 1993; Verghese et al., 2002). Populations consuming diets containing phytonutrients such as fish (n-3 fatty acids), fresh fruits, vegetables (vitamin and minerals), whole grains, legumes (protein), nuts (vitamin E), olive oil, antioxidants (resveratrol from wine) and dairy (probiotics) have been strongly associated with a reduction in chronic diseases such as cancer (Willett, 1995) and considered as a desirable approach than treatment (Liu, 2003).
Legumes such as soybeans were utilized extensively in human foods traditionally in many regions of the world. In contrast, western diets have lower legume consumption patterns. Among legumes, Soybean (Glycine max) is unique with phytochemicals specifically isoflavones and others such as saponins, phytates, protease inhibitors, phenolic acids, lecithin, dietary fiber, phytosterols and omega-3-fatty acids (Messina, 1999; Omoni and Aluko, 2005). Epidemiological, cell culture and animal studies revealed health benefits of soybean isoflavones by modulation of metabolism (Spector et al., 2003). Evidence suggests the anti-proliferative effects of Soybean extracts, genistein and daidzein in colon cancer cell lines in vitro (Zhu et al., 2002; Kuo, 1996; Guo et al., 2004).
Interventions and role of dietary fiber in human health has been a point of interest for epidemiologists, nutritionists and the entire fraternity of research scientists for more than three decades. Prebiotic properties of Synergy1® (Inulin: Oligofructose = 1:1) (soluble dietary fiber) in stimulation of probiotics such as Bifidobacteria and Lactobacillus sp., provide a basis for its role in the prevention of colon cancer by modulating colonic microenvironment (Lim et al., 2005).
The anti-cancer effect of bioactive compounds of plant origin (Phytochemicals) using colon cancer cell lines such as Caco-2, HT-29 and DLD-1 are well studied (Zessner et al., 2008; Duessel et al., 2008; Dong et al., 2007). Animal experiments provide information about the effects of dietary non-nutritive compounds at safer doses. However, in vitro cell culture studies elucidate the key molecular mechanisms involved in the balance of cell cycle through up-regulation and or down-regulation of critical genes. Inhibition of inflammatory cytokines (COX-2), stimulation of antioxidative (Catalase, glutathione peroxidase, Superoxide-dismutase) and specific detoxification enzymes (Glutathione-S-transferase), apoptosis responsive factors, cell differentiation, inhibition of cell proliferation, anti-angiogenesis, histone-deacetylase inhibition causing growth arrest in colon cancer cells (Archer et al., 2005) are some of the mechanisms exerted by phytonutrients. Isoflavones present in soybean demonstrated different mechanisms in colon cancer cell lines at transcription and post translational levels (Radzikowski et al., 2004; Messina et al., 1994; Messina and Bennink, 1998).
Single dietary components found in plant foods are not exclusively responsible for the health benefits of food rather; dietary plant compounds have a synergistic effect in disease prevention (Gerber, 2001). Food synergy has been defined as additive or more than additive influences of foods and food constituents on health. Food synergy attempts to identify the food components with relevant health effects (Jacobs and Steffen, 2003). Liu (2004) suggested that additive and synergistic effects of phytochemicals in whole foods improve health benefits when compared to an isolated purified phytochemical extract or a dietary supplement, which may be responsible for their anti-carcinogenic properties (Liu, 2004).
Recently, research is focused on identifying specific combinations of phytochemicals or foods offering greater chemopreventive potential. Understanding the influence of various bioactive compounds on molecular interactions and immuno-modulatory responses led to the emerging strategy of combinational chemoprevention (De Kok et al., 2008). Thus, using combinations of dietary ingredients provides a promising solution in controlling cancer by either halting or inhibiting carcinogenesis.
Based on present results from animal experiments (Fisher 344 male rats), more than 70% reductions in colon tumors were observed in rats fed combinations of Synergy1® and Soybean meal. This initiated to conduct mechanistic study using human colon cancer caco-2 cell lines. The objective of this study was to elucidate the inhibitory mechanisms of Synergy1® and Soybean meal extracts in combination on human colon cancer cell line Caco-2.
MATERIALS AND METHODS
Chemicals
Individual isoflavones (genistein, daidzein, glycetin and equol) and butyric acid were purchased from Sigma Aldrich, St. Louis, MO.
Preparation of Synergy1® and Soybean Extracts
Synergy1® (BeneoTM) was obtained from Orafti, Belgium and yellow Soybean Meal (SM) was obtained from the local market (Garden Cove, Huntsville, AL). Synergy1® is a white crystalline powder (inulin:oligofructose = 1:1) soluble in aqueous solution. Twenty five milligrams of Synergy1® was dissolved in one hundred milliliters of autoclaved water and filtered using a Whatman No.1 paper. Soybean meal was weighed (25 mg) and added to 100 mL of cold 80% methanol and phenolic compounds were extracted overnight under restricted light. The extract was defatted using a SoxtecTM 2043 extraction unit (FOSS, Hoganas Sweden). The extracts were then filtered using a Whatman No.1 paper and the filtrate was collected, concentrated using a rotary evaporator (Buchi Rotavapor R-205 equipped with self cleaning dry vacuum systemTM Model 2025) at 40°C for 15 min and stored at-20°C. Before treating the cells, soybean extract and individual isoflavones were dissolved in dimethyl sulfoxide (DMSO).
General Procedures
Caco-2 human colon cancer cells (ATCC HTB-37) were obtained from the American Type Culture Collection. Cells were maintained in Dulbecco’s modified Eagles Media (DMEM) with 10% fetal bovine serum. For assay, 5x104 Caco-2 cells well-1 were seeded to a 24 well culture plate and incubated at 37°C and 7% CO2 until development of a monolayer. After the development of a monolayer, 400 μL of fresh media (DMEM-serum free) was added to the wells, selected concentrations of Syn1®, SM (25, 50 and 100 μg mL-1) singly and in combinations, isoflavone extracts (0.25, 0.5 and 1 μg mL-1) and butyric acid (0.1, 0.2 and 0.4 μg mL-1) made up to 100 μL with saline were added and incubated for 12 h and 24 h.
Determination of Lactate Dehydrogenase (LDH)
The cytotoxic effects of Syn1® and SM extracts (25-100 μg mL-1) were measured by the release of lactate dehydrogenase (LDH) from the Caco-2 cells (12 and 24 h incubation) into the culture supernatant. A 10% trition-X solution was used as a positive control. LDH was quantified by a colorimetric cytotoxicity detection kit (LDH) (Roche Diagnostics, Indianapolis, IN) and the absorbance was read at 490/600 nm. Percentage LDH release was calculated based on control or untreated caco-2 cells.
Gluthaione-S-Transferase Activity in Cell Lysates
The supernatant was collected from the incubated cells (12 and 24 h) and centrifuged at 10,000x g for 10 min in PBS buffer (pH 7). Glutathione-s-transferase was determined using a microplate reader as described by Habig et al. (1974) and kinetic absorbances were read at 340 nm at five time points.
Apoptosis
Induction of apoptosis by Syn1®, SM, isoflavones and butyric acid extracts was analyzed by (1) Caspase-3 assay (2) DNA fragmentation and (3) morphological changes.
General Procedure
To determine the effect of Syn1®, SM and butyric acid in induction of apoptosis, 1x105 Caco-2 cells were treated with Syn1®, SM extracts (25-100 μg mL-1) and butyric acid (0.1-0.4 μg mL-1) for 12 and 24 h. Incubated cells were extracted with PBS (pH 7) and centrifuged at 10,000 g for 15 min. Cell lysis buffer was added to the pellet and incubated for 10 min. Later, cells were centrifuged at 15,000 g for 15 min.
Caspase-3 Activity
Caspase-3 activity was determined using a Caspase kit (Promega Corporation, Madison, WI).
DNA Fragmentation
Apoptosis was determined by measuring DNA fragmentation by using a cell death detection ELISA kit (Cell Death Detection ELISA, Roche Molecular Biochemicals, Indianapolis, IN). Spectrophotometric analysis was carried out using an ELISA plate reader at 405 nm for the histone related DNA fragmentation.
Morphological Changes
Caco-2 cells grown in slide flaskets were incubated for 24 h with Syn1®, SM extracts and butyric acid and stained with Giemsa stain (0.1% in PBS) for 5 min. Cells were washed three times with PBS and photographs were taken using a Nikon CT 120 microscope (Nikon Instruments Inc., Melville, NY) and SPOT 2.0 Digital Camera (Verghese et al., 2006).
Statistical Analysis
Statistical analysis of data was conducted (SAS 9.1, 2007) (SAS Institute Inc., Cary, NC) using analysis of variance (One-Way ANOVA) to determine any significant differences among the groups. When significant, means were separated using Tukey’s Studentized Range Test. The level of significance was set at p<0.05.
RESULTS
Cytotoxic Effects of Bioactive Compounds: Lactate Dehydrogenase (LDH) Assay
Percentage LDH was calculated based on LDH release in control or untreated caco-2 cells. Cells incubated with Syn1® and SM extracts singly and in combinations for 12 h had higher LDH release relative to the untreated cells (Fig. 1). Following 24 h incubation of cells with Syn1® and SM extracts showed cytotoxicity at all concentrations compared to the control (Fig. 2). However, LDH release was higher after incubation (24 h) of cells with Syn1® and SM extracts compared to 12 h of incubation. Cells incubated with a mixture of Syn1® and SM extracts had over 50% LDH release and ranged from 47.0-75.2% after 12 h and 57.7-84.3% after a 24 h incubation period. Following 24 h of incubation of cells with higher concentrations (200 and 400 μg mL-1) of Syn1® and SM, similar LDH release was seen.
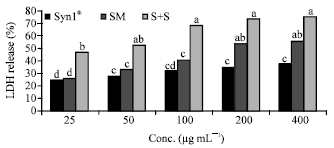 | |
| Fig. 1: | Effect of Syn1® and SM extracts on LDH release (12 h). Syn1®: Synergy1®, SM: Soybean meal, S+S: Synergy1®+Soybean. abcdeBars with the same letter are not significantly different using Tukey’s studentized range test (p≤0.05) |
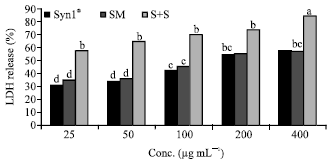 | |
| Fig. 2: | Effects of Syn1® and SM extracts on LDH release (24 h). Syn1®: Synergy1®, SM: Soybean meal, S+S: Synergy1®+Soybean. Bars with the same letter are not significantly different using Tukey’s studentized range test (p≤0.05) |
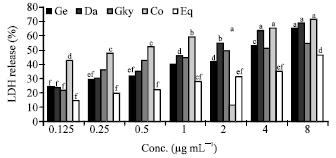 | |
| Fig. 3: | Effects of Isoflavone extracts and Equol on LDH release (12 h). Ge: Genistein, Da: Daidzein, Gly: Glycitein, Co: Combo, Eq: Equol. Bars with the same letter are not significantly different using Tukey’s studentized range test (p≤0.05) |
Cells incubated (12 h) with equal combination of the three isoflavones (genistien, daidzein and glycitein) had higher LDH release (%) ranging from 42.8%-71.5% (Fig. 3). Daidzein was effective at higher concentrations in inducing cytotoxicity in Caco-2 cells after a 12 h incubation period. Equol, a metabolite of daidzein resulted in lower LDH release (%) in cells incubated for 12 (15-46.5%) and 24 h (17-47.8%) (Fig. 3, 4). Genistein induced higher LDH release (%) ranging from 47.5% at 0.125 μg mL-1 to 81.8% at 8 μg mL-1 after 24 incubation of cells (Fig. 4). Cells treated with combinations of isoflavones (combo) showed a decrease in LDH release after 24 h of incubation at lower concentrations and gradually increased at concentrations >2 μg mL-1.
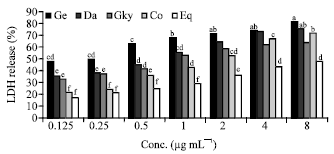 | |
| Fig. 4: | Effects of Isoflavone extracts and Equol on LDH release (24 h). Ge: Genistein, Da: Daidzein, Gly: Glycitein, Co: Combo, Eq: Equol. Bars with the same letter are not significantly different using Tukey’s studentized range test (p≤0.05) |
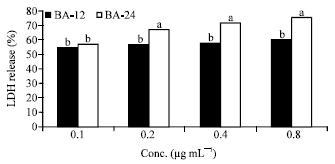 | |
| Fig. 5: | Effects of Butyric acid on LDH release (12 and 24 h). BA: Butyric acid. Bars with the same letter are not significantly different using Tukey’s studentized range test (p≤0.05) |
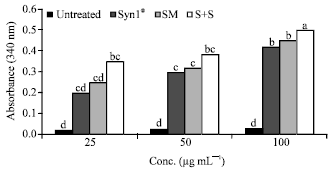 | |
| Fig. 6: | Effects of Syn1® and SM extracts on Gluathione-s-transferase (GST) activity in cell lysates. Syn1®: Synergy1®, SM: Soybean meal, S+S: Synergy1®+Soybean. Bars with the same letter are not significantly different using Tukey’s studentized range test (p≤0.05) |
Figure 5 shows the effects of butyric acid extracts (0.1-0.8 μg mL-1) on cytotoxicity as measured by LDH (%) release in Caco-2 cells ranged from 54.5-60% after 12 h and 57.1-75.4% following 24 h incubation.
GST Activity in Cell Lysates
Following 24 h incubation, cells treated with combinations of Syn1®+SM showed higher GST activity (nmol/min/mg protein) compared to the cells treated with the extracts alone and ranged from a low of 0.35 (25 μg mL-1) to a high of 0.5 (100 μg mL-1) (Fig. 6). Caco-2 cells incubated with butyric acid for 24 h showed significantly (p<0.05) higher GST activity (nmol/min/mg protein) compared to the untreated cells (Fig. 7). A dramatic increase in GST activity was seen in cells treated with butyric acid at 0.4 μg mL-1 (0.7) and with the activity seen highest at 0.8 μg mL-1 (0.85).
Caspase-3 Activity
Caspase-3 activity was significantly (p<0.05) higher in Caco-2 cells incubated with all treatments compared to the untreated cells (Control). Time dependent effect on Caspase-3 activity (fold increase) was apparent in cells that were incubated with Syn1® (Fig. 8), SM (Fig. 9) and Syn1® + SM extracts (Fig. 10) for 12 and 24 h with concentrations 25, 50 and 100 μg mL-1. Although, Syn1® extract alone had no dose response Caspase-3 activity on cells incubated for 12 and 24 h (Fig. 8).
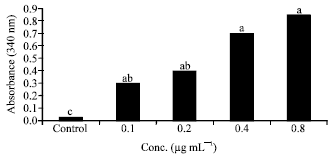 | |
| Fig. 7: | Effects of butyric acid on GST activity in cell lysates. BA: Butyric acid, GST: Glutathione-s-transferase. Bars with the same letter are not significantly different using Tukey’s studentized range test (p≤0.05) |
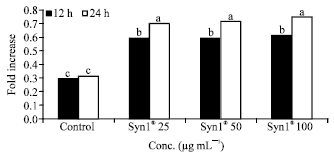 | |
| Fig. 8: | Effect of Syn1® extracts on Caspase-3 activity. Syn1®: Synergy1®. Bars with the same letter are not significantly different using Tukey’s studentized range test (p≤0.05) |
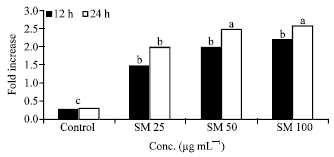 | |
| Fig. 9: | Effect of SM extracts on Caspase-3 activity. SM: Soybean meal. Bars with the same letter are not significantly different using Tukey’s studentized range test (p≤0.05) |
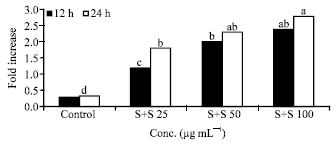 | |
| Fig. 10: | Effect of Syn1® and SM (S+S) extracts on Caspase-3 activity. Syn1®: Synergy1®, SM: Soybean meal. Bars with the same letter are not significantly different using Tukey’s studentized range test (p≤0.05) |
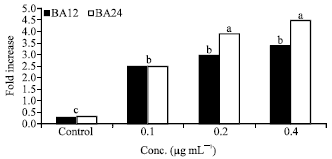 | |
| Fig. 11: | Effect of butyric acid on Caspase-3 activity. BA: Butyric acid. Bars with the same letter are not significantly different using Tukey’s studentized range test (p≤0.05) |
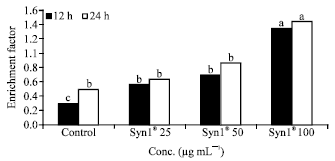 | |
| Fig. 12: | Effect of Syn1® extracts on Histone-DNA fragmentation (12 and 24 h). Syn1®: Synergy1®. Bars with the same letter are not significantly different using Tukey’s studentized range test (p≤0.05) |
Caco-2 cells incubated for 12 and 24 h with SM extracts had higher Caspase-3 activity at all doses (25-100 μg mL-1) compared to the cells treated with Syn1® extracts, with an increase of 6 fold in the cells treated with SM extract at 100 μg mL-1 compared to the control (Fig. 9). Highest (over 4 fold increase) Caspase-3 activity was observed after 12 and 24 h incubation of cells with Syn1® + SM at 100 μg mL-1 (Fig. 10). The effect of incubation time seems to be greater at the higher (0.4 μg mL-1) compared to the lower concentrations (0.2 μg mL-1) of butyric acid in Caco-2 cells (Fig. 11).
Histone-Associated DNA Fragmentation
The release of oligonucleotides into the cytoplasm was higher in Caco-2 cells treated with SM ranging between 0.6-1.2 and 0.7-1.52 following 12 and 24 h respectively (Fig. 13).
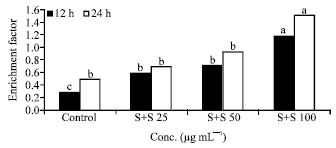 | |
| Fig. 13: | Effect of SM extract on Histone-DNA fragmentation (12 and 24 h). SM: Soybean meal. Bars with the same letter are not significantly different using Tukey’s studentized range test (p≤0.05) |
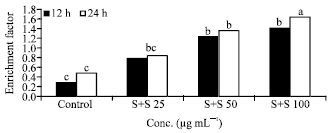 | |
| Fig. 14: | Effect of Syn1® and SM extracts on Histone-DNA fragmentation (12 and 24 h). Syn1®: Synergy1®, SM: Soybean meal. Bars with the same letter are not significantly different using Tukey’s studentized range test (p≤0.05) |
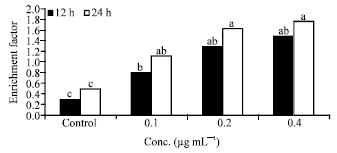 | |
| Fig. 15: | Effect of butyric acid on Histone-DNA fragmentation (12 and 24 h). Butyric acid. Bars with the same letter are not significantly different using Tukey’s studentized range test (p≤0.05) |
DNA fragmentation (expressed as Enrichment Factor-EF) in Syn1® treated cells ranged between 0.57-1.35 after 12 h and 0.64-1.45 after 24 h incubation (Fig. 12). Caco-2 cells incubated with Syn1® and SM alone induced higher DNA fragmentation at a concentration of 100 μg mL-1 following 12 and 24 h, compared to the lower concentrations of these extracts. However, in cells incubated with a mixture of Syn1® and SM, highest release of oligonucleotides into the cytoplasm after 12 and 24 h expressed as EF was seen at both 50 μg mL-1 (1.25 and 1.38) and 100 μg mL-1 (1.42 and 1.65) (Fig. 14). DNA fragmentation in Caco-2 cells was directly proportional to the concentration of the extracts as well as incubation time. The Enrichment Factor (EF) increased with longer incubation time and higher concentrations (0.2 and 0.4 μg mL-1) of butyric acid in Caco-2 cells (Fig. 15). The highest difference in EF was observed after cells were treated with 0.4 μg mL-1 of butyric acid for 12 and 24 h.
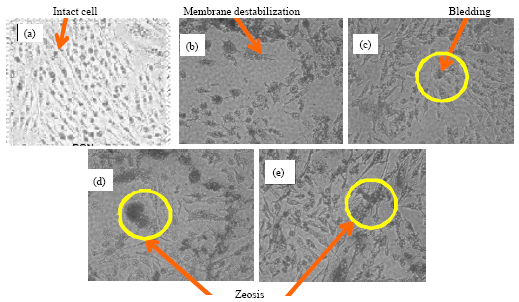 | |
| Fig. 16: | Morphological changes in Caco-2 cells incubated with Syn1®, SM, Syn1®+ SM extracts at 100 μg mL-1 and Butyric acid extract at 0.8 μg mL-1 for 24, (a) Untreated, (b) Syn1® extract, (c) SM extract, (d) Syn1®+ SM and (e) Butyric acid |
Morphological Changes in Caco-2 Cells
Morphology of Caco-2 cells after incubation (24 h) of cells with Syn1®, SM, Syn1® + SM (100 μg mL-1) and butyric acid (0.8 μg mL-1) extracts were observed using microscopy showing membrane blebbing, a characteristic of apoptotic activity (Fig. 16a-e). Untreated Caco-2 cells did not show any features of apoptotic activity. Cells treated with treatment extracts had membrane destabilization, zeosis and associated cell condensation. Cellular and nuclear changes were higher after 24 h incubation of cells with Syn1® + SM and butyric acid extracts compared to the other treatments.
DISCUSSION
Isoflavones which are scarcely distributed in nature are seen in higher concentrations in soybean, cloves, chick peas and some Chinese herbs (Putman et al., 2007). Growth inhibitory effects of genistein in hormone-dependent and hormone-independent cancer cells has been proposed to involve signal transduction pathways (Yu et al., 2005; Messina and Bennink, 1998) and induction of phase-2 metabolizing enzymes such as sulfotransferases (Chen et al., 2008). Beyer-Schlmeyer et al. (2003) reported that fermented dietary fiber sources are potent cell growth inhibitors compared to butyrate alone. Inhibition of Caco-2 cell proliferation by genistein, daidzein and soy extract was reported by MacDonald et al. (2005). A cell culture study using human colon cancer cell line (Caco-2 cells) was conducted in our laboratory to determine the chemopreventive effects of Syn1®, SM and their synergistic effects in equal combinations and butyrate.
Chemopreventive agents inhibit the growth of tumor cells by inducing cell specific cytotoxicity. Our results show that Syn1®, SM extracts and butyrate induced cell cytotoxicity in a time dependent manner. Incubation (24 h) with Syn1® and SM extracts in combination resulted in an LDH release ranging between 50-80% in Caco-2 cells. Extensive research on the effects of isoflavones especially genistein and daidzein on colon cancer cell lines is well studied with explanatory mechanisms (Radzikowski et al., 2004; Messina et al., 1994; Messina and Bennink, 1998). Blay et al. (2010) reported non-cytotoxic effects of genistein and daidzein following 24 h incubation in mouse macrophages with concentrations ranging between 1-25 μM. Caco-2 cells incubated with these isoflavones (25-100 μM) had no significant LDH release into the medium after 24 h incubation (Subhashinee et al., 2007). Effect of specific herbs (Rosemary, Sage, Oregano and Echinacea) on cell membrane integrity assessed by LDH assay reported higher (40%) release in Caco-2 cells incubated with rosemary extracts (Aherne et al., 2007). Initially, LDH (%) release was higher in cells incubated (12 h) with a combination of Isoflavones (combo). However, with increasing incubation time genistein induced higher cytotoxicity after 24 h incubation ranging between 47-80%. A study conducted in our laboratory (Boateng et al., 2008) using kidney beans (0.5-25 mg/100 mL) reported dose dependent cytotoxic effects after 24 and 48 h incubation in Caco-2 cells. Apoptotic effects of Syn1®, SM and butyrate was demonstrated by measuring caspase-3 activity. Caspase-3 activity was over 3-4 fold higher in cells incubated with SM at highest concentration (100 μg mL-1) compared to the Syn1® at same levels. However, cells incubated with combinations of treatments showed higher caspase-3 activity (Fig. 10). LT97 colon cancer cells incubated with Synergy1® supernatant fraction (SFS) induced apoptosis with a significant increase in PARP (Poly ADP-ribose polymerase) cleavage as demonstrated by western blotting (Munjal et al., 2009). Similar study using phytic acid (0.25-4 g mol L-1) reported cytotoxic and apoptotic effects in caco-2 cells incubated for 24 and 48 h (Verghese et al., 2006). Effect of butyrate on apoptosis has been attributed to multiple pathways such as trigerring caspase-3, PARP cleavage and up-regulation of pro-apoptotic BAK signaling (Avivi-Green et al., 2002). In this study, Caco-2 cells incubated with butyrate (0.1-0.4 g mL-1) showed higher capase-3 activity compared to the untreated cells at 12 and 24 h incubation as measured by fold increase ranging between 2.5 to 4.5 (Fig. 11). Anti-tumor effects of capsaicin (a principal flavor ingredient in hot chilli pepper) was demonstrated using LoVo colon cancer cell lines resulting in the induction of apoptosis, decreased cell viability, DNA fragmentation and cell morphology changes (Yang et al., 2009). Morphological changes in cells incubated with Syn1®, SM and butyrate showed membrane destabilization and zeosis indicating characteristic apoptotic activity. Caco-2 cells treated with Syn1®, SM, isoflavones and butyric acid exhibited distinct inhibitory effects compared to control/untreated cells.
CONCLUSION
Present results demonstrated that Syn1®, SM in combinations caused loss of cell membrane integrity and cell death as shown by increased LDH release showing cytotoxicity. Caco-2 cells incubated with the treatment extracts resulted in higher histone-associated-DNA fragmentation and higher Caspase-3 activity compared to untreated cells through inducing apoptosis. Results from this study support anticancer effects of dietary fiber such as Synergy1® (Inulin and Oligofructose) that were conducted as pilot studies in our lab. The chemopreventive properties of Soybean isoflavones are still inconclusive from extensively conducted preclinical and clinical studies. This cell culture study (Caco-2 cells) approaches to combine in determining the effects of functional ingredients such as Synergy1® and Soybean in human colon cancer cell line (Caco-2) provides additional information on their potential protective effects and applicability as nutraceuticals. Consumption of functional foods and nutraceuticals such as soluble dietary fiber and isoflavones may have inhibitory effects on the development of colon cancer through primary and secondary prevention.
Higher LDH release, histone associated DNA fragmentation, caspase-3 activation and membrane blebbing in Caco-2 cells suggests the anti-cancer effects of Syn1® and SM as a result of synergism between bioactive components present in these functional ingredients.
ACKNOWLEDGMENTS
This study was supported by ORAFTI, Tienen, Belgium and The Alabama Agricultural Experiment Station, Alabama AandM University, Normal, AL.
REFERENCES
- Aherne, S.A., J.P. Kerry and N.M. O'Brien, 2007. Effects of plant extracts on antioxidant status and oxidant induced stress in Caco-2 cells. Br. J. Nutr., 97: 321-328.
PubMed - Archer, S.Y., J. Johnson, H.J. Kim, Q. Ma and H. Mou et al., 2005. The histone deacetylase inhibitor butyrate downregulates cyclin B1 gene expression via p21/WAF-1 dependent mechanism in human colon cancer cells. Am. J. Physiol. Gastrointest Liver Physiol., 289: G696-703.
PubMed - Avivi-Green, C., S. Polak-Charcon, Z. Madar and B. Schwartz, 2002. Different molecular events account for butyrate-induced apoptosis in two human colon cancer cell lines. J. Nutr., 132: 1812-1818.
PubMed - Beyer-Sehlmeyer, G., M. Glei, E. Hartmann, R. Hughes and C. Persin et al., 2003. Butyrate is only one of several growth inhibitors produced during gut flora-mediated fermentation of dietary fibre sources. Br. J. Nutr., 90: 1057-1070.
Direct Link - Blay, M., A.E. Espinel, M.A. Delgado, I. Baiges, C. Blade, L. Arola and J. Salvado, 2010. Isoflavone effect on gene expression profile and biomarkers of inflammation. J. Pharma Biomed. Anal., 51: 382-390.
PubMed - Boateng, J., M. Verghese, L.T. Walker, L. Shackelford and C.B. Chawan, 2008. Antitumor and cytotoxic properties of dry beans (Phaseolus sp. L.): An in vitro and in vivo model. Int. J. Cancer Res., 4: 41-51.
CrossRefDirect Link - De Kok, T.M., S.G. van Beda and M.M. Manson, 2008. Mechanisms of combined action of different chemopreventive dietary compounds: A review. Eur. J. Nutr., 47: 51-59.
CrossRefPubMedDirect Link - Dong, M., X. He and R.H. Liu, 2007. Phytochemicals of black bean seed coats: isolation, structure elucidation, and their antiproliferative and antoxidant activities. J. Agric. Food Chem., 55: 6044-6051.
PubMed - Duessel, S., R.M. Heuertz and U.R. Ezekiel, 2008. Growth inhibition of human colon cancer cells by plant compounds. Clin. Lab. Sci., 21: 151-157.
PubMed - Gerber, M., 2001. The comprehensive approach to diet: A critical review. J. Nutr., 131: 3051S-3055S.
PubMedDirect Link - Guo, J.M., B.X. Xiao, D.H. Liu, M. Grant and S. Zhang et al., 2004. Biphasic effect of daidzein on cell growth of human colon cancer cells. Food Chem. Toxicol., 42: 1641-1646.
PubMed - Habig, W.H., M.J. Pabst and W.B. Jakoby, 1974. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem., 249: 7130-7139.
CrossRefPubMedDirect Link - Jacobs, D.R. and L.M. Steffen, 2003. Nutrients, foods and dietary patterns as exposures in research: A framework for food synergy. Am. J. Clin. Nutr., 78: 508S-513S.
PubMed - Kuo, S.M., 1996. Antiproliferative potency of structurally distinct dietary flavanoids L-against D-galactosamine and carbon tetra chloride on in situ perfused in liver. Ind. J. Physiol. Pharmacol., 45: 41-48.
PubMed - Lim, C.C., L.R. Ferguson and G.W. Tannock, 2005. Dietary fibres as prebiotics: Implications for colorectal cancer. Mol. Nutr. Food Res., 49: 609-619.
PubMed - Liu, R.H., 2003. Health benefits of fruits and vegetables are from additive and synergistic combinations of phytochemicals. Am. J. Clin. Nutr., 78: 517S-520S.
PubMedDirect Link - Liu, R.H., 2004. Potential synergy of phytochemicals in cancer prevention: Mechanism of action. J. Nutr., 134: 3479S-3485S.
PubMedDirect Link - MacDonald, R.S., J. Guo, J. Copeland, Jr. J.D. Browning, D. Sleper, G.E. Rottinghaus and M.A. Berhow, 2005. Environmental influences on isoflavones and saponins in soybeans and their role in colon cancer. J. Nutr., 135: 1239-1342.
PubMed - Messina, M. and M. Bennink, 1998. Soyfoods, isoflavones and risk of colonic cancer: A review of the in vitro and in vivo data. Clin. Endocrinol. Metab., 4: 707-728.
PubMed - Messina, M.J., 1999. Legumes and soybeans: Overview of their nutritional profiles and health effects. Am. J. Clin. Nutr., 70: 439S-450S.
CrossRefPubMedDirect Link - Messina, M.J., V. Persky, K.D.R. Setchell and S. Barnes, 1994. Soy intake and cancer risk: A review of the in vitro and in vivo data. Nutr. Cancer, 21: 113-131.
PubMedDirect Link - Munjal, U., M. Glei, B.L. Pool-Zobel and D. Scharlau, 2009. Fermentation products of Inulin type fructans reduce proliferation and induce apoptosis in human colon tumor cells of different stages of carcinogenesis. Br. J. Nutr., 102: 663-671.
PubMed - Omoni, A.O. and R.E. Aluko, 2005. Soybean foods and their benefits: Potential mechanisms of action. Nutr. Rev., 63: 272-283.
CrossRefDirect Link - Potter, J.D., 1993. Colon cancer-do the nutritional epidemiology, the gut physiology and the molecular biology tell the same story. J. Nutr., 123: 418-423.
PubMed - Putman, S.E., A.M. Scutt, K. Bicknell, C.M. Priestley and E.M. Williamson, 2007. Natural products as alternate treatments for metabolic bone disorders and for maintenance of bone health. Phytother Res., 21: 99-112.
PubMed - Radzikowski, C., J. Wietrzyk, G. Grynkiewicz and A. Opolski, 2004. Genistein: A soy isoflavone revealing a pleiotropic mechanism of action- clinical implications in the treatment and prevention of cancer. Postepy Hig. Med. Dosw, 58: 128-139.
PubMed - Spector, D., M. Anthony, D. Alexander and L. Arab, 2003. Soy consumption and colorectal cancer. Nutr. Cancer, 47: 1-12.
PubMed - Wijeratne, S.S and S.L. Cuppett, 2007. Soy isoflavones protect the intestine from lipid hydroperoxide mediated oxidative damage. J. Agric. Food Chem., 55: 9811-9816.
PubMed - Verghese, M., D.R. Rao, C.B. Chawan, L.T. Walker and L. Shackelford, 2006. Anticarcinogenic effect of phytic acid (IP6): Apoptosis as a possible mechanism of action. LWT-Food Sci. Technol., 39: 1093-1098.
CrossRefDirect Link - Yu, Z., Y. Tang, D. Hu and J. Li, 2005. Inhibitory effect of genistein on mouse colon cancer MC-26 cells involved TGF-beta1/Smad pathway. Biochem. Biophys Res. Commun., 333: 827-832.
PubMed - Verghese, M., D.R. Rao, C.B. Chawan, L.L. Williams and L. Shackelford, 2002. Dietary inulin suppresses azoxymethane-induced aberrant crypt foci and colon tumors at the promotion stage in young fisher 344 rats. J. Nutr., 132: 2809-2813.
PubMedDirect Link - Willet, W.C., 1995. Diet, nutrition and avoidable cancer. Environ. Health Prospect., 103: 165-170.
CrossRefDirect Link - Zessner, H., L. Pan, F. Will, K. Klimo and J. Knauft et al., 2008. Fractionation of polyphenol-enriched apple juice extracts to identify constituents with cancer chemopreventive potential. Mol. Nutr. Food Res., 52: S28-44.
PubMed - Zhu, Q., J. Meisinger, D.H. van Thiel, Y. Zhang and S. Mobarhan, 2002. Effects of soybean extract on morphology and survival of Caco-2, SW620 and HT-29 cells. Nutr. Cancer, 42: 131-140.
PubMedDirect Link - Yang, K.M., J.O. Pvo, G.Y. Kim, R. Yu and I.S. Han et al., 2009. Capsaicin induces apoptosis by generating recative oxygen species and disrupting mitochondrial transmembrane potential in human colon cancer cell lines. Cell Mol. Biol. Lett., 14: 497-510.
PubMed