
K. Sabitha
Bhagwan Mahavir Hospital and Research Center, Hyderabad, AP, India
M. Vishnuvardhan Reddy
Department of Otolaryngology, Osmania Medical College, Koti, Hyderabad, AP, India
Mrs. Kaiser Jamil
Bhagwan Mahavir Hospital and Research Center, Hyderabad, AP, India
International Journal of Cancer Research
Year: 2009 | Volume: 5 | Issue: 2 | Page No.: 58-68







ABSTRACT
The XRCC1 gene promotes the efficiency of the DNA repair process and a deletion or modification of this gene appears to result in lethal phenotypes, as we have investigated this hypothesis in this study. It was found that Arg280His variant, was associated with increased risk of head and neck cancer [OR = 2.11, 95% CI, 1.28-3.46) in smokers as compared to nonsmokers. The frequency of Arg/Arg polymorphisms was higher in the smokers (64.82%) and nonsmokers (69.03%) compared to the cancer patients (46.6%) suggesting that this particular mutation may not be adverse genotype, whereas the heterozygous variant (Arg/His) and homozygous variant His/His were more frequent in the patients than in the smokers and nonsmokers suggesting that these genotypes were high risk groups. The distribution of Arg/Arg, Arg/Gln and Gln/Gln genotypes at the Arg399Gln site for nonsmokers was 44.6, 43.22 and 6.45% and for smokers it was 47.5, 43.4 and 8.96%, respectively. Individual who smoked >15 years and carried the codon 280 Arg/His genotype had an OR = 2.30, 95% CI, 1.01-5.30 and who smoked less than 15 years had an OR = 1.64, 95% CI, 0.78-3.46, these results suggested that the codon Arg280His genotypes were associated with high risk of head and neck cancers in heavy smokers.
PDF Abstract XML References Citation
How to cite this article
K. Sabitha, M. Vishnuvardhan Reddy and Mrs. Kaiser Jamil, 2009. Mutations in XRCC1 Gene Alters the Genetic Risks of Head and Neck Cancer Patients. International Journal of Cancer Research, 5: 58-68.
DOI: 10.3923/ijcr.2009.58.68
URL: https://scialert.net/abstract/?doi=ijcr.2009.58.68
DOI: 10.3923/ijcr.2009.58.68
URL: https://scialert.net/abstract/?doi=ijcr.2009.58.68
INTRODUCTION
Tobacco chewing and smoking have been identified as the major risk factors for head and neck cancer in India. Tobacco and alcohol account for more than 75% of squamous cell head and neck cancer (including the nasal passages, sinuses, mouth, throat, larynx (voice box), swallowing passages and salivary glands), specific carcinogenic mechanisms are unclear. Studies have suggested that genetic susceptibility plays an important role in risk of SCCHN (squamous cell carcinoma of the head and neck) because even though tobacco and alcohol exposure are the most important risk factor, only small fraction of smokers will ever develop SCHHN.
Cigarette smoke is a rich source of chemical carcinogens and ROS (Reactive Oxygen Species). Polycyclic aromatic hydrocarbons, aldehydes, aromatic amines, nitrosamines, etc., are thought to be carcinogenic components present in tobacco. But chewing of tobacco with betel quid increases the concentrations of carcinogenic tobacco specific nitrosamines and reactive oxygen species in the mouth (Nair et al., 1999). Metabolites of tobacco and alcohol (Pfeifer et al., 2002; Phillips, 2002) cause DNA damage by producing oxidative stress, alkylations, bulky adducts and strand breaks. Altered DNA repair capacity may increase the risk of various cancers, including head and neck cancer (Cheng et al., 1998; Hu et al., 2002; Spitz et al., 1989). Carcinogenic components present in tobacco can produce DNA bulky adducts that may lead to DNA damage (Vineis et al., 1996).
ROS are present in both the gas-phase and the particulate matter (Pryor, 1997; Kiyosawa et al., 1990) and include oxygen radicals, e.g., superoxide radicals (O2•) and hydroxyl radicals (OH•) and some derivatives of O2• that lack unpaired electrons, e.g., hydrogen peroxide (H2O2) and hypochlorous acid (HOCl) (Halliwell and Cross, 1994). The accumulation of ROS leads to oxidative stress, which is a risk factor for cancer development (Bankson et al., 1993). ROS can initiate lipid peroxidation, oxidize proteins and cause damage to DNA indirectly or directly (Joenje, 1989; Floyd, 1990). Indirect damage includes inactivation of target enzymes, such as those involved in DNA synthesis (Friedber et al., 1995). Direct DNA damage includes DNA strand breaks, creation of a basic sites and base adduct formation, such as thymine glycol, 5-hydrozymethyluracil and 8-hydroxy-2-deoxyguanosine. Up to 4x105 oxidatively altered DNA base residues are introduced per day in each cell (Friedber et al., 1995; Roldan-Arjona et al., 1997).
There is considerable evidence that DNA repair capacity is genetically determined. Consequently, DNA repair enzyme gene polymorphisms, which may alter the function or efficiency of DNA repair, may contribute to cancer susceptibility (Spitzet al., 1997; Knight et al., 1993; Wu et al., 1998; Wei et al., 1996). Bulky adduct lesions induced by chemical carcinogens are repaired through the Nucleotide Excision Repair (NER) pathway (Benhamou and Sarasin, 2000). Base damage and DNA single strand breaks are repaired through the BER (Base Excision Repair) pathway (Wilson and Thomposn, 1997). This pathway is a multistep process that requires the activity of several proteins (Friedber et al., 1995). The XRCC1 (X-ray Repair Cross-Complementing group 1) protein is an important component of BER, because it serves as a scaffold for two other proteins, DNA ligase III and POLB (polymerase β) and also serves as a single-strand break sensor by its interaction with PARP (poly-(ADP-ribose) polymerase) (Caldecott et al., 1994, 1996; Masson et al., 1998).
XRCC1 was originally discovered in radiation-sensitive mutants and assigned to the double-strand break/recombination pathway of DNA repair (Zdzienicka, 1995). A BCRT domain found in many proteins responsive to DNA damage with cell cycle check point functions has also been identified in XRCC1 (Bork et al., 1997). XRCC1 is thought to form protein complexes with DNA ligase III via the BCRT domain in its COOH terminus and with DNA polymerase B via., its NH2- terminal domain to repair gaps left during base excision repair (Kubota et al., 1996). Because amino acid residues at the protein-protein interfaces of multiprotein complexes and residues involved in the active sites play a role in enzyme function; it is possible that the XRCC1 polymorphisms may result in altered efficiency of the protein.
XRCC1 has been mapped to human chromosome 19q13.2–13.3 (Thompson et al., 1989; Mohrenweiser et al., 1989) and two polymorphisms that lead to amino acid substitutions have been described in codon 280 (exon 9, base 27466 G to A, Arg to His) and codon 399 (exon 10, base 28152 G to A, Arg to Gln) ( Schneider et al., 2008; Shen et al., 1998). Both the polymorphisms occur at residues that are conserved across hamster, mouse and human (Lamerdin et al., 1995) but their functional consequences remain unknown.
Recent genetic association studies on cancer risk have focused on identifying effects of single nucleotide polymorphisms/haplotypes in candidate genes, among which DNA repair genes are increasingly studied because of their critical role in maintaining genome integrity. Some studies have investigated associations between XRCC1 variants in DNA repair genes and cancer risk (Yin et al., 2007; Hu et al., 2005). Pachouri et al. (2007) examined the association of a polymorphism in a XRCC1 gene and lung cancer in a case-control study. They observed XRCC1 polymorphisms at codon 399 were found to be protective in the development of lung cancer. The codon 194 Trp/Trp genotype was associated with a slightly increased risk of lung cancer (Pachouri et al., 2007).
Assays that measure DNA repair capacity suggest that this function can vary widely among individuals. Cancer patients have lower DNA repair capacities than healthy controls (Mohrenweiser and Jones, 1998). This finding supports the hypothesis that variants in DNA repair genes could affect cancer susceptibility. In the present study, support the hypothesis that XRCC1 polymorphisms could affect the capacity of cells to repair ROS-induced DNA damage and consequently influence head and neck cancer risk. Using a case-control study, it was tested whether these polymorphisms were associated with head and neck cancer risk and examined gene-environment interactions, by testing whether XRCC1 polymorphisms modified the risk effect of cigarette smoking, a measure of ROS exposure.
Studies have suggested that genetic susceptibility plays an important role in risk of SCCHN because even though tobacco and alcohol exposure are the most important risk factor, only small fraction of smokers will ever develop SCCHN.
MATERIALS AND METHODS
Enrolment of Cases and Controls
The present case controlled study comprises of 150 smoker cases with histopathological confirmed SCC of the head and neck cancers (smokers), 145 non-cancerous smokers and 155 healthy controls. Both incident and prevalent cases of head and neck cancers were included in this study. All the patients were recruited from the out-patient department of the ENT- Hospital, Hyderabad from Jan 2006 to Jan 2008; in this study all 150 cases had SCC of the various head and neck parts. This study was approved by the institutional ethical committee.
Inclusion and Exclusion Criteria
The inclusion criterion for the cases was the presence of histopathologicaly diagnosed SCC of the head and neck cancers. Information on smoking and alcohol habits were obtained in a structured questionnaire and tabulated. The inclusion criterion for the controls was absence of prior history of cancer or pre-cancerous lesions. Unrelated healthy controls were recruited from blood donors who accompanied patient seeking treatment at the hospital and from members of different community centers. After obtaining informed consent, all individuals were personally interviewed using the questionnaire. Information on age, gender, occupation, region of origin, type of tobacco used, daily frequency and duration of use was recorded. Data pertaining to histopathological diagnosis and clinical staging of the disease was collected from the hospital records. The cases reported habits such as smoking of bidi or cigarettes. The majority had a single habit some cases reported dual habits comprising of different combinations of smoking and drinking.
Collection of Blood Samples
Venous blood samples (5 mL) were collected in syringes (heparinised) from patients and control groups (smokers and healthy controls). Immediately after collection, genomic DNA was extracted and used for Polymerase Chain Reaction (PCR) to determine the genotypes.
XRCC1 Genotyping
Genomic DNA was isolated from peripheral blood and XRCC1 genotypes at the Arg280His and Arg399Gln sites were determined by PCR-based restriction fragment length polymorphism as described by Zhongbin et al. (2005). The primers for the XRCC1 Arg280His polymorphism were 5’TGGGGCCTGGATTGCTGGGTCTG 3’ and 5’ CAGCACCACTACCACACCCTGAAGG 3’ which generate a 280 bp fragment. The primers for the XRCC1 Arg399Gln polymorphism were 5’ TTGTGCTTTCTCTGTGTCCA 3’ and 5’TCCTCCAGCCTTTTCTGATA 3’, which generate a 615 bp, fragment. PCR was performed in a 25 μL reaction mixture containing approximately 100 ng DNA, 1.0 μM each primer, 0.2 mM each dNTP, 2.0 mM MgCl2, 1.0 U Taq DNA polymerase with 1x reaction buffer and 2% dimethyl sulfoxide. Amplification of the Arg280His site was carried out under the following conditions: an initial melting step of 2 min at 95 °C, followed by 35 cycles of 30 sec at 94 °C, 30 sec at 65 °C 45 sec at 72 °C and a final elongation of 7 min at 72 °C. The reaction conditions for the Arg399Gln site were identical except that the annealing temperature was 62 °C.
The restriction enzymes RsaI and MspI were used to distinguish the Arg280His and Arg399Gln polymorphisms, respectively. The wild-type Arg280His allele generates a single band representing the entire 280 bp fragment, the variant allele (His/His) generates 140 bp fragment and the heterozygote (Arg/His) displays 2 bands (280, 140 bp). Arg399Gln site, the wild-type allele (Arg/Arg) generates 2 DNA bands (377 and 238 bp), the variant allele (Gln/ Gln) has a single 615 bp fragment and the heterozygote (Arg/Gln) displays all 3 bands (615, 377 and 238 bp). The restriction products were analyzed by electrophoresis on a 3% agarose gel containing ethidium bromide.
Statistical Analysis
The associations between XRCC1 polymorphisms and risk of head and neck cancer were estimated by Odds Ratios (OR) and their 95% Confidence Intervals (CI), which were calculated by unconditional logistic regression. The ORs were adjusted for age, gender and smoking status. All analysis were performed using the EPI6 software (Epiinfo6 CDC). Bonferroni’s correction of p-value was evaluated when multiple comparison analysis was used.
RESULTS
The category of the disease of the patients with head and neck cancer as per the diagnosis of the oncologists are given here. In this study group of 150 cases found that about 24% were suffering from cancer of Larynx followed by hypo-pharynx 23.40%, tongue 18%, epiglottis 14%, oropharynx 7.3%, nasopharynx 6 and 4% had cancer of paranasal sinuses and 3.3% oral cavity. It was found that the patients were mostly males between the age group of 30 to 80 years. The details of their smoking-bidi or cigarette habits are given in Table 1.
In Table 1 presented some of the characteristics of cases and controls. Besides age match also presented smokers in both controls and cases; however there was only a small percentage of female participation in this study. It was also recorded the number of cigarettes /bidi’s the group was using, in both controls and cases (Table 1). This data has some correlation with genotypes; hence details were included.
The Arg280 His variant allele was associated with increased risk of head and neck cancer (OR = 2.11, 95% CI, 1.28-3.46) compared with smokers and nonsmokers. The frequency of the Arg/Arg, Arg/His, His/His genotypes (codon 280) was 46.6, 38, 15.3%, respectively. The frequency Arg/Arg polymorphisms were higher in the smokers (64.82%) and nonsmokers (69.03%) compare to the cases (46.6%). It is suggested that this may not be the adverse genotype, whereas the heterozygous variant (Arg/His) and homozygous variant His/His which are more frequent in the cases than in the smokers and nonsmokers may be the high risk genotype.
| Table 1: | Characteristics of the study and control group (smokers) |
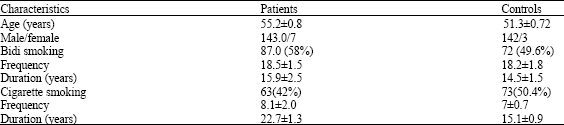 | |
In logistic regression model the heterozygous variant type (Arg/His) appeared to be a significant risk factor for SCCHN (OR = 2.07, 95% CI, 1.22-3.53) (Table 3). It was also a significant risk factor compared to smokers (OR = 1.91, 95% CI, 1.11-3.29) (Table 2). The XRCC1 His/His genotype was highly significant risk factor compared to smokers (OR = 2.81, 95% CI, 1.21-6.62; p = 0.007) and nonsmokers (OR = 5.86, 95% CI, 2.12-17.00; p = 00006). We did not find statistically significant differences among smokers and nonsmokers with His/His genotypes (OR = 2.09, 95% CI, 0.68-6.68). But it was found His/His genotype frequency was more in smokers compared to nonsmokers (Table 4). However, the association between His/His variant genotypes and the risk of developing head and neck cancers belonging to either category of smokers or nonsmokers group was found to be higher.
The distribution of Arg/Arg, Arg/Gln and Gln/Gln genotypes at the Arg399Gln site for nonsmokers was 44.6, 43.22 and 6.45% and for smokers it was 47.5, 43.4 and 8.96%, respectively. The distribution of the Arg/Arg Arg/Gln genotypes among head and neck cancer cases (44.6%) did not differ from the controls except Gln/Gln genotype (15.3%). XRCC1 Arg/Gln polymorphism was not associated with a higher risk of developing head and neck cancer.
| Table 2: | XRCC1 genotype frequencies in head and neck cancer cases and controls (smokers) |
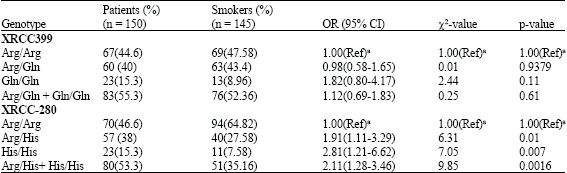 | |
| (Ref)a used as reference group | |
| Table 3: | XRCC1 genotype frequencies in head and neck cancer cases and controls (non-smokers) |
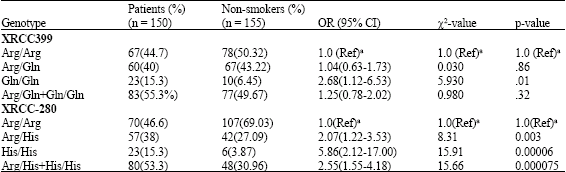 | |
| (Ref)a used as reference group | |
| Table 4: | XRCC1 genotype frequencies in smokers and non-smokers |
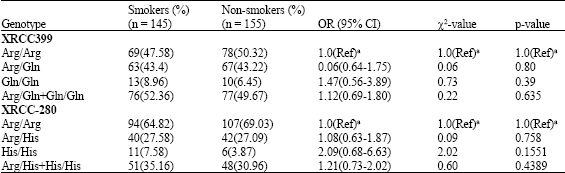 | |
| (Ref)a used as reference group | |
| Table 5: | XRCC1 codon 280 and 399 polymorphisms and smoking, stratified analysis |
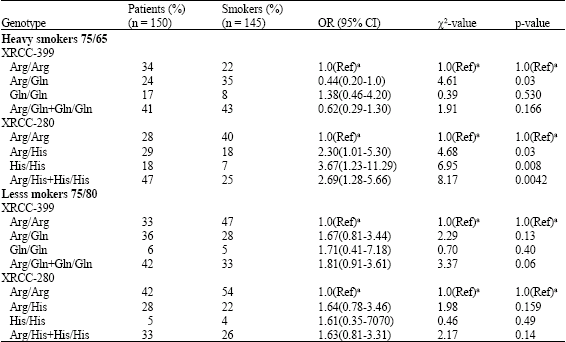 | |
| (Ref)a used as reference group | |
Arg/Gln genotype (OR = 0.980, 95% CI, 0.58-1.61) did not differ from individuals with the Arg/Arg genotype. Association between the XRCC1 Gln/Gln homozygous variant genotype and the risk of developing head and neck cancer was found to be significant (OR = 5.86, 95% CI, 2.12-11.0; p-0.01).
The association of XRCC1 Arg280His and Arg399Gln polymorphisms and the risk of developing head and neck cancer were analyzed in the smokers’ category. Decreasing level of smoking constantly decreased head and neck cancer risk. For example, individuals who smoked >15 years and carried the Arg/His genotype (codon 280) had an (OR = 2.30, 95% CI, 1.01-5.30) and compared to those who smoked less than 15 years and had an (OR = 1.64, 95% CI, 0.78-3.46), these results suggested that the Arg280His genotypes were associated with statistically higher risk of head and neck cancers, in high frequency smokers than in the low frequency smokers (Table 5).
DISCUSSION
DNA repair pathways play a vital role in maintaining genetic integrity and it is becoming clear that defects in repair pathways are connected to many different types of diseases. DNA repair systems maintain genomic integrity, in the face of environmental insults, cumulative effects of age and general DNA replication errors. XRCC1 play a role in the multi step base excision repair pathways where non bulky base adducts produced by methylation, oxidation, reduction, or fragmentation of base by ionization radiation or oxidation damages are removed (Yu et al., 1999). Although the specific function of XRCC1 has not been identified, it is believed that XRRC1 complexes with DNA ligase III via a BRCT (BRCA1 carboxyl-terminal) domain in COOH terminus and with DNA polymorphisms β in this NH2 terminus repairs the gaps left during base excision. The XRRC1 codon 280 polymorphisms are located in the linker region that separates the DNA POLB interacting domain from the PARP interaction domain. The codon 399 polymorphisms located in the COOH terminal side of the PARP interacts with the BRCT domain. Although the 2 polymorphisms lead to amino acid substitution, there is no direct data on the functional consequences.
Cigarette smoke is a major cause of a variety of malignancies including cancers of the larynx, oral cavity and pharynx, esophagus, bladder and lung. Numerous studies have consistently shown that cigarette smoke plays an important role as an environmental etiological factor in the development of head and neck cancers (Hecht, 2003). Cigarette smoke contains a myriad of genotoxic agents and carcinogens such as nitrosamine 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK). Some studies reported that the XRCC1 399Gln/399Gln genotypes were associated with a decreased cancer risk among heavy smokers (OR = 0.71, 95% CI, 0.51–0.99), but an increased cancer risk among light smokers (OR = 1.38, 95% CI, 0.99 – 1.94) (Hung et al., 2005). It was also observed a significantly lower risk of head and neck cancer (OR = 0.44; 95% CI, 0.20-1.0) in heavy smokers and risk among light smokers (OR = 1.67; 95% CI, 0.81-3.44) carrying the XRCC1 399Arg/Gln genotype. These findings suggest that the XRCC1 Arg280His polymorphism is associated with the risk of head and neck cancer. Individuals with the variant allele were at 80% greater risk compared with those with the homozygous wild-type genotype after adjustment for radon and tobacco exposure. This is a very interesting finding, but it only relates to the particular genotypes of XRCC1 in this study group and perhaps may not hold true for other genotypes.
The activation and detoxification of carcinogens and ROS, as well as the repair of the damage induced by xenobiotics like polycyclic aromatic hydrocarbons, aromatic amines and N-nitroso compounds, are important aspects in carcinogenesis. Both are under genetic control and there is increasing evidence of genetic variation of these pathways in the human population. Therefore, integrating information on allelic variants of these genes may be useful in determining groups of individuals at highest risk from environmental exposure. In the case of XRCC1, the analysis of possible interactions with GSTM1 polymorphisms could be appropriate given the role of this enzyme in detoxifying products of oxidative stress (Strange and Fryer, 1999).
The XRCC1 codon 280 polymorphisms are located in the vicinity of two sequences mediating the protein-protein interactions with ADPRT and polymerase (ElKhamisy et al., 2003; Pleschke et al., 2000). Thus, the variant alleles may encode a twisted XRCC1 protein, resulting in increased or decreased affinity to other proteins, which may affect normal DNA repair capability. This results indicated that there was significant difference in distribution of genotype of XRCC1 Arg280His and the risk was increased in those with XRCC1 280Arg/His and XRCC1 280Arg/His+His/His genotypes. These data were consistent with earlier reports on risk of cancers of the breast (Duell et al., 2001), bladder and lung (Ratnasinghe et al., 2001) XRCC1 may be associated with altered function of the coding products and thus contribute to the individual’s genetic susceptibility to head and neck cancer.
Functional studies of XRCC1 suggest that the codon 399 Gln allele may be associated with multiple DNA damage phenotypes in human cells and tissues. Lunn et al. (1999) reported that the 399Gln allele was associated with an increased aflatoxin DNA adduct burden in placental tissue and an elevated glycophorin A mutant frequency in erythrocytes. Duell et al. (2000) reported a positive association between the same allele and detection of polyphenol DNA adducts from blood mononuclear cells, as well as a positive association between the variant 399Gln allele and baseline sister chromatid exchange frequencies in lymphocytes from smokers. Together, these studies suggest a role for XRCC1 in the repair of multiple DNA damage end points in human cells and tissues and imply that the 399Gln allele of XRCC1 has an important, potentially harmful phenotype. Consistent with this hypothesis, two recent epidemiological studies have shown a positive association between the XRCC1 Gln allele and cancer. Sturgis et al. (1999) reported an OR of 1.6 (95% CI, 1.0 -2.6) for the XRCC1 codon 399 Gln/Gln genotype in a case-control study of head and neck cancer and Divine et al. (2001) observed an odds ratio of 2.8 (95% CI, 1.2-7.9) for XRCC1 codon 399 Gln/Gln genotype in a case-control study of lung cancer. However, two other epidemiological studies reported contrary findings. Stern et al. (2001) observed an inverse association between XRCC1 codon 399 Gln/Gln genotype and ladder cancer; ORs for smoking were stronger among carriers of the codon 399 Arg/Arg genotype compared with Gln-containing genotypes. Similarly, Watson et al. (2002) reported an inverse association between XRCC1 codon 399 Gln/Gln genotype and cancer of the head and neck; ORs for smoking were stronger among carriers of the codon 399 Arg/Arg genotype.
These findings showed that there was a positive association between the XRCC1 (codon 399) Gln/Gln genotype in head and neck cancer which was consistent with two previously reported epidemiological studies by Sturgis et al. (1999) and Divine et al. (2001) as well as with published functional studies by Lunn et al. (1999) and Duell et al. (2000). According to our observation XRCC1 codon 399 Gln/Gln genotype was not a statistically significant risk factor (OR = 1.82, 95% CI, 0.80-4.17) in smokers but it is significant compared to non-smokers (OR = 2.68, 95% CI, 1.12-6.53). The frequencies of this polymorphism in smokers were higher as compared to non-smokers. These data may also support hypothesis that smokers the XRCC1 codon 399 Gln/Gln genotypes were unable to carry out the function of the DNA repair process and therefore were at high risk. The G to A transition in XRCC1 codon 399 results in the change from Arg to Gln in the XRCC1 BRCA1 COOH terminus domain that interacts with poly (ADP) ribose polymerase. Hence, the variant amino acid (i.e., Gln instead of Arg) at 399 of XRCC1 protein may be less efficient in DNA repair (Sturgis et al., 1999; Tae et al., 2004). These variant genotypes at codon 399 of XRCC1 increased the risk of head and neck squamous cell carcinomas.
CONCLUSION
One of the strengths of this study is its prospective design. The collection of covariate data (e.g., smoking and alcohol) before case diagnosis minimized the potential for recall bias for measures of environmental exposures and the availability of these data also allowed us to explore gene-environment interactions. This is the first observational study to examine the association between the Arg280His XRCC1 polymorphism and risk of head and neck cancer. This data suggest a high risk for head and neck cancer in individuals with the Arg280His XRCC1 polymorphism. Also, the Gln/Gln genotypes among smokers in cases and controls showed higher risk in cases compared to controls. These findings support the Hypothesis that XRCC1 Arg399Gln amino acid change may alter the phenotype of the XRCC1 protein, resulting in deficient DNA repair. It was conclude that XRCC1 genotype may be related to head and neck cancer risk, but this relationship is complex. Present results and those of other recent epidemiological studies of XRCC1 suggest that BER represents an important biochemical pathway for future epidemiological studies of cancer.
REFERENCES
- Bankson, D.D., Kestin, M. and N. Rifai, 1993. Role of free radicals in cancer and atherosclerosis. Clin. Lab. Med., 13: 463-480.
PubMed - Benhamou, S. and A. Sarasin, 2000. Variability in nucleotide excision repair and cancer risk: A review. Mutat. Res., 462: 149-158.
PubMed - Bork, P., K. Hofmann, P. Bucher, A.F. Neuwald, S.F. Altschul and E.V. Koonin, 1997. A superfamily of conserved domains in DNA damage-responsive cell cycle checkpoint proteins. FASEB J., 11: 68-76.
PubMedDirect Link - Caldecott, K.W., S. Aoufouchi, P. Johnson and S. Shall, 1996. XRCC1 polypeptide interacts with DNA polymerase b and possibly poly (ADP-ribose) polymerase and DNA ligase III is a novel molecular nick-sensor in vitro. Nucleic Acids Res., 24: 4387-4394.
PubMedDirect Link - Caldecott, K.W., C.K. McKeown, J.D. Tucker, S. Ljungquist and L.H. Thompson, 1994. An interaction between the mammalian DNA repair protein XRCC1 and DNA ligase III. Mol. Cell. Biol., 14: 68-76.
PubMedDirect Link - Cheng, L., S.A. Eicher, Z. Guo, W.K. Hong, M.R. Spitz and Q. Wei, 1998. Reduced DNA repair capacity in head and neck cancer patients. Cancer Epidemiol. Biomarkers Prev., 7: 465-468.
PubMedDirect Link - Divine, K., F. Gilliland, R. Crowell, C. Stidley, T. Bocklage, D. Cook and S. Belinsky, 2001. The XRCC1 399 glutamine allele is a risk factor for adenocarcinoma of the lung. Mutat. Res., 461: 273-278.
PubMedDirect Link - Duell, E.J., R.C. Millikan, G.S. Pittman, S. Winkel and R.M. Lunn et al., 2001. Polymorphisms in the DNA repair gene XRCC1 and breast cancer. Cancer Epidemiol. Biomarkers Prev., 10: 217-222.
PubMedDirect Link - Duell, E.J., J.W. Wiencke, T.J. Cheng, A. Varkonyi and Z.F. Zuo et al., 2000. Polymorphisms in the DNA repair genes XRCC1 and ERCC2 and biomarkers of DNA damage in human blood mononuclear cells. Carcinogenesis (Lond.), 21: 965-971.
PubMedDirect Link - El-Khamisy, S.F., M. Masutani, H. Suzuki and K.W. Caldecott, 2003. A requirement for PARP-1 for the assembly or stability of XRCC1 nuclear foci at sites of oxidative DNA damage. Nucl. Acids Res., 31: 5526-5533.
PubMedDirect Link - Floyd, R.A., 1990. The role of 8-hydroxyguanine in carcinogenesis. Carcinogenesis (Lond.), 11: 1447-1450.
PubMedDirect Link - Halliwell, B. and C.E. Cross, 1994. Oxygen-derived species: their relation to human disease and environmental stress. Environ. Health Perspect., 102: 5-12.
PubMedDirect Link - Hung, R.J., P.Brennan, F. Canzian, N. Szeszenia-Dabrowska and D. Zaridze et al., 2005. Large-scale investigation of base excision repair genetic polymorphisms and lung cancer risk in a multicenter study. J. Natl. Cancer Inst., 97: 567-576.
CrossRefPubMedDirect Link - Kiyosawa, H., M. Suko, H. Okudaira, K. Murata, T. Miyamoto and M.H. Chung et al., 1990. Cigarette smoking induces formation of 8-hydroxydeoxyguanosine, one of the oxidative DNA damages in human peripheral leukocytes. Free Radic. Res. Commun., 11: 23-27.
PubMed - Kubota, Y., R.A. Nash, A. Klungland, P. Schar, D.E. Barnes and T. Lindahl, 1996. Reconstitution of DNA base excision-repair with purified human proteins: Interaction between DNA polymerase b and the XRCC1 protein. EMBO. J., 15: 6662-6670.
PubMedDirect Link - Lamerdin, J.E., M.A. Montgomery, S.A. Stilwagen, L.K. Scheidecker and R.S. Tebbs et al., 1995. Genomic sequence comparison of the human and mouse XRCC1 DNA repair gene regions. Genomics, 25: 547-554.
PubMed - Lunn, R., R. Langlois, L. Hsieh, C. Thompson and D. Bell, 1999. XRCC1 polymorphisms: Effects on aflatoxin B1-DNA adducts and glycophorin a variant frequency. Cancer Res., 59: 2557-2561.
PubMedDirect Link - Masson, M., C. Niedergang, V. Schreiber, S. Muller, J. Menissier-de-Murcia and G. de Murcia, 1998. XRCC1 is specifically associated with poly(ADP-ribose) polymerase and negatively regulates its activity following DNA damage. Mol. Cell Biol., 18: 3563-3571.
PubMedDirect Link - Mohrenweiser, H.W. and I.M. Jones, 1998. Variation in DNA repair is a factor in cancer susceptibility: A paradigm for the promises and perils of individual and population risk estimation? Mutat. Res., 400: 15-24.
PubMed - Nair, U.J., J. Nair, B. Mathew and H. Bartsch, 1999. Glutathione S-transferase M1 and T1 null genotypes as risk factors for oral leukoplakia in ethnic Indian betel quid/tobacco chewers. Carcinogenesis, 20: 743-748.
PubMedDirect Link - Pachouri, S.S., R.C. Sobti, P. Kaur and J. Singh, 2007. Contrasting impact of DNA repair gene XRCC1 polymorphisms Arg399Gln and Arg194Trp on the risk of lung cancer in the north-Indian population. DNA Cell Biol., 26: 186-191.
PubMed - Pfeifer, G.P., M.F. Denissenko, M. Olivier, N. Tretyakova, S.S. Hecht and P. Hainaut, 2002. Tobacco smoke carcinogens, DNA damage and p53 mutations in smoking-associated cancers. Oncogene, 21: 7435-7451.
PubMedDirect Link - Phillips, D.H., 2002. Smoking-related DNA and protein adducts in human tissues. Carcinogenesis, 23: 1979-2004.
PubMedDirect Link - Pleschke, J.M., H.E. Kleczkowska, M. Strohm and F.R. Althaus, 2000. Poly (ADP-ribose) binds to specific domains in DNA damage checkpoint proteins. J. Biol. Chem., 275: 40974-40980.
PubMedDirect Link - Pryor, W.A., 1997. Cigarette smoke radicals and the role of free radicals in chemical carcinogenicity. Environ. Health Perspect., 105: 875-882.
PubMedDirect Link - Ratnasinghe, D., S.X. Yao and J.A. Tangrea, Y.L. Qiao and M.R. Andersen et al., 2001. Polymorphisms of the DNA repair gene XRCC1 and lung cancer risk. Cancer Epidemiol. Biomarkers Prev., 10: 119-123.
PubMedDirect Link - Roldan-Arjona, T., Y.F. Wei, K.C. Carter, A. Klungland and C. Anselmino et al., 1997. Molecular cloning and functional expression of a human cDNA encoding the antimutator enzyme 8-hydroxyguanine-DNA glycosylase. Proc. Natl. Acad. Sci., 94: 8016-8020.
PubMedDirect Link - Schneider, J., V. Classen and S. Helmig, 2008. XRCC1 polymorphism and lung cancer risk. Expert Rev. Mol. Diagn., 8: 761-780.
PubMed - Strange, R.C. and A.A. Fryer, 1999. The glutathione S-transferases: Influence of polymorphism on cancer susceptibility. IARC Sci. Publ., 148: 231-249.
PubMed - Sturgis, E.M., E.J. Castillo, L. Li, R. Zheng and S.A. Eicher et al., 1999. Polymorphisms of DNA repair gene XRCC1 in squamous cell carcinoma of the head and neck. Carcinogenesis (Lond.), 20: 2125-2129.
PubMedDirect Link - Stern, M.C., D.M. Umbach, G.C.H. Van, R.M. Lunn and J.A. Taylor, 2001. DNA repair gene XRCC1 polymorphisms, smoking and bladder cancer risk. Cancer Epidemiol. Biomark. Prev., 10: 125-132.
PubMedDirect Link - Shen, M.R., I.M. Jones and H. Mohrenweiser, 1998. Nonconservative amino acid substitution variants exist at polymorphic frequency in DNA repair genes in healthy humans. Cancer Res., 58: 604-608.
PubMedDirect Link - Spitz, M.R., J.J. Fueger, N.A. Beddingfield, J.F. Annegers and T.C. Hsu et al., 1989. Chromosome sensitivity to bleomycin-induced mutagenesis, an independent risk factor for upper aerodigestive tract cancers. Cancer Res., 29: 4626-4628.
PubMedDirect Link - Spitz, M.R., R.S. McPherson, H. Jiang, T.C. Hsu and Z. Trizna et al., 1997. Correlates of mutagen sensitivity in patients with upper aerodigestive tract cancer. Cancer Epidemiol. Biomark. Prev., 6: 687-692.
PubMedDirect Link - Tae, K., H.S. Lee, B.J. Park et al, 2004. Associations of DNA repair gene XRCC1 polymorphisms with head and neck cancer in Korean population. Int. J. Cancer, 111: 805-808.
CrossRefPubMedDirect Link - Thompson, L.H., L.L. Bachinski, R.L. Stallings, G. Dolf, C.A. Weber, A. Westerveld and M.J. Siciliano, 1989. Complementation of repair gene mutations on the hemizygous chromosome 9 in CHO: A third repair gene on human chromosome 19. Genomics, 5: 670-679.
PubMed - Vineis, P., G. Talaska, C. Malaveille, H. Bartsch, T. Martone, P. Sithisarankul and P. Strickland, 1996. DNA adducts in urothelial cells: Relationship with biomarkers of exposure to arylamines and polycyclic aromatic hydrocarbons from tobacco smoke. Int. J. Cancer, 65: 314-316.
PubMed - Watson, M.A., A.F. Olshan, M.C. Weissler and D.A. Bell, 2002. XRCC1 polymorphisms and head and neck cancer. Cancer Lett., 178: 181-186.
PubMed - Wei, Q., L. Cheng, W.K. Hong and M.R. Spitz, 1996. Reduced DNA repair capacity in lung cancer patients. Cancer Res., 56: 4103-4107.
PubMedDirect Link - Wilson, D.M. and L.H. Thompson, 1997. Life without DNA repair. Proc. Natl. Acad. Sci. USA, 94: 12754-12757.
PubMedDirect Link - Wu, X., J. Gu, C.I. Amos, H. Jiang, W.K. Hong and M.R. Spitz, 1998. A parallel study of in vitro sensitivity to benzo[a]pyrene diol epoxide and bleomycin in lung carcinoma cases and controls. Cancer (Phila.), 83: 1118-1127.
PubMedDirect Link - Yin, J., U. Vogel, Y. Ma, R. Qi, Z. Sun, H. Wang, 2007. The DNA repair gene XRCC1 and genetic susceptibility of lung cancer in a northeastern Chinese population. Lung. Cancer, 56: 153-160.
PubMed - Yu, Z., J. Chen, B.N. Ford, M.E. Brackley and B.W. Glickman, 1999. Human DNA repair systems: An overview. Environ. Mol. Mutagen., 33: 3-20.
PubMedDirect Link - Zdzienicka, M.Z., 1995. Mammalian mutant's defective in the response to ionizing radiation-induced DNA damage. Mutat. Res., 336: 203-213.
PubMed - Zhongbin, Z., W. Junxiang, J. Xipeng, J. Taiyi, S. Hongbing, L. Daru and X. Zhaolin, 2005. Genetic polymorphisms in xrcc1, ape1, adprt, xrcc2 and xrcc3 and risk of chronic benzene poisoning in a chinese occupational population. Cancer Epidemiol. Biomark. Prev., 14: 2614-2619.
PubMedDirect Link - Hu, Z., H. Ma, F. Chen, Q. Wei and H. Shen, 2005. XRCC1 polymorphisms and cancer risk: A meta-analysis of 38 case-control studies. Cancer Epidemiol. Biomarkers Prev., 143: 1810-1818.
PubMed