
R. Karthikeyan
Center of Advanced Study in Marine Biology, Annamalai University, Parangipettai 608 502, Tamil Nadu, India
S. Karthigayan
Center of Advanced Study in Marine Biology, Annamalai University, Parangipettai 608 502, Tamil Nadu, India
M. Sri Balasubashini
Center of Advanced Study in Marine Biology, Annamalai University, Parangipettai 608 502, Tamil Nadu, India
S. Vijayalakshmi
Center of Advanced Study in Marine Biology, Annamalai University, Parangipettai 608 502, Tamil Nadu, India
S.T. Somasundaram
Center of Advanced Study in Marine Biology, Annamalai University, Parangipettai 608 502, Tamil Nadu, India
T. Balasubramanian
Center of Advanced Study in Marine Biology, Annamalai University, Parangipettai 608 502, Tamil Nadu, India
International Journal of Cancer Research
Year: 2007 | Volume: 3 | Issue: 4 | Page No.: 167-173







ABSTRACT
The present study was carried out to assess the antitumor effect of venom from snake, Hydrophis spiralis on the Ehrlich Ascites Carcinoma (EAC). Four groups of albino Swiss mice were treated with three doses of the sub lethal dose of venom, viz., 0.418, 4.18 and 6.27 μg kg-1 body weight (intraperitoneal injection) along with the standard drug 5 flurouracil (20 mg kg-1 b.w.). The biochemical analysis and rest was left to calculate the mean survival time. In EAC bearing mice, mean life span tumor volume, hemoglobin, red blood cell and lymphocytes were significantly decreased when compared to the normal animals. Whereas, body weight, neutrophils and viable tumor cell count was increased in the EAC bearing mice. These changes were brought back to near normal levels in different treatment groups. The macromolecule concentration of peritoneal cells, such as, DNA, RNA and protein, were altered in the EAC bearing mice and observed to be near normal in the treatment groups. The caspase 3 activity was significantly increased in the peritoneal cells of the treatment groups when compared to the EAC bearing mice. The role of apoptotic cascade in EAC cell death was confirmed by the DNA fragmentation on agarose gel. Apart from the antitumor effect, snake venom reduced the tumor burden on the liver and altered the changes in the activities of alanine transaminase (ALT), aspartate transaminase (AST) and alkaline phosphatase (ALP). Hence the venom from H. spiralis has a potential antitumor effect on the EAC bearing mice.
PDF Abstract XML References Citation
How to cite this article
R. Karthikeyan, S. Karthigayan, M. Sri Balasubashini, S. Vijayalakshmi, S.T. Somasundaram and T. Balasubramanian, 2007. Antitumor Effect of Snake Venom (Hydrophis spiralis) on Ehrlich Ascites Carcinoma Bearing Mice. International Journal of Cancer Research, 3: 167-173.
DOI: 10.3923/ijcr.2007.167.173
URL: https://scialert.net/abstract/?doi=ijcr.2007.167.173
DOI: 10.3923/ijcr.2007.167.173
URL: https://scialert.net/abstract/?doi=ijcr.2007.167.173
INTRODUCTION
Cancer continues to represent the largest cause of mortality in the world and claims over 6 million lives every year Abdullaev et al. (2000). An extremely promising strategy for cancer prevention today is chemoprevention, which is defined as the use of synthetic or natural agents (alone or in combination) to block the development of cancer in humans.
The anticancer agents search has aroused the interest of scientists since the beginning of this century and anticancer effect of the venom of Naja sp. on adenocarcinoma cells was observed by Calmette et al. (1933). From that time on, a number of papers have been published on the subject (Calmette et al., 1933; Yeh et al., 2001; Sun et al., 2003; Mora et al., 2005). Particularly, when used in combination of venoms, snake venom has greatly improved the treatment of several tumors cell lines (Lipps, 1994). Snake venom is effective against tumors in vivo and in vitro, but causes less toxicity in normal cells (Araya and Lomonte, 2007).
The aim of the present study is to investigate the antitumour effect of the crude venom of snake on the Ehrlich ascites caricinoma and the metabolic changes in serum and peritoneal fluid of the tumor bearing mice in response to the treatment with different sublethal dose of the venom.
MATERIALS AND METHODS
Venom
Sea snake Hydrophis spiralis was collected during January 2006 from Kalpakkam to Cuddalore waters of Coromandel Coast (India). The identification of live snake was carried out in the zoological survey of India. Venom from live snake was squeezed out manually by milking method, lyophilized immediately and stored at 4°C until further use (Ali et al., 1999).
Lethality and Toxicity Assay
The toxicity of the Crude Venom (CV) was quantitatively assayed by intraperitoneal injection (i.p.) in albino rats (10 per dose) using 5-10 mg of crude venom in 0.9% NaCl (w/ v). LD50 was determined at five dose levels and observed for period of 24 h following i.p. injection.
Animals and Ascites Tumor
The inoculum for inducing Ehrlich’s Ascites Carcinoma (EAC) was kindly provided by the Amala Cancer Research Institute, Thirissur, Kerala (India). EAC was thereafter propagated by weekly intraperitoneal injection of 0.3 mL freshly drawn ascites fluid (diluted 1:5 in sterile saline) from a donor mice bearing ascites tumor of 6-8 days old into each Swiss albino mice. Transplantation was carried out using sterile disposable syringes under aseptic conditions.
Experimental Design
In this experiment, 72 mice were randomly assorted into 6 groups (12 animals per group). Group 1:Normal (received 100 μL of sterile saline by i.p), Group 2: EAC control (received 100 μL of sterile saline by i.p), Group 3: EAC induced mice + 1% of LD50 dose of venom in 100 μL of sterile saline (0.418 μg kg-1 bw. by i.p-Low dose: LD), Group 4: EAC induced mice +10% of LD50 dose of venom in 100 μL of sterile saline (4.18 mg kg-1 bw. by i.p-Medium dose: MD), Group 5:EAC induced mice +15% of LD50 dose of venom in 100 μL of sterile saline (6.27 mg kg-1 bw. by i.p-High dose: HD), Group 6; EAC induced mice + Standard 5-floururacil (20 mg kg-1 bw.).
Total experimental period was 10 days and after administering the last dose, 6 animals in each group were fasted overnight, anaesthetized and sacrificed by cervical decapitation. The liver tissues of animals were stored in 20°C for further analysis. The remaining animals were left to calculate the mean survival time.
Antitumor effect of venom was assessed by observing the following changes with respect to body weight, ascites tumor volume, packed cell volume, viable and non-viable tumor cell count, Mean Survival Time (MST) and percentage Increase in Life Span (%ILS). MST of each group containing six mice was monitored by recording the mortality daily for six weeks and ILS (%) was calculated using following equation described by Abu-Sinna et al. (2003).
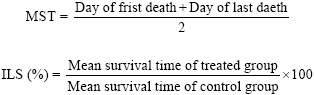 |
Hematological Study
Hemoglobin content, Red Blood Cell (RBC) and White Blood Cell (WBC) counts were measured from freshly withdrawn tail vein blood (Wintrobe et al., 1961). Differential counts (Dacie and Lewis, 1953) were analysed in Leishman stained blood smears from normal, EAC control and venom treated groups.
Biochemical Parameters
Aspartate transaminase (AST) and Alanine transaminase (ALT) activities were determined by the method of Reitman and Frankel (1957) using Qualigens (Mumbai, India) reagent kit. Alkaline phosphatase (ALP) was assayed by the method of King and Armstrong (1988) using Qualigens (Mumbai, India) reagent kit.
Assay for Total Protein, DNA, RNA and Caspase-3 Activity
The peritoneal fluid in heparinised tubes was centrifuged at low speed, washed in sterile saline and the cells were suspended in saline. The cells were then stained with 0.32% trypan blue and counted microscopically in a hemocytometer. The number of viable and dead cells were determined by the dye exclusion method of Maclimans et al. (1957). Total protein was assayed by the method of Lowry et al. (1951). Nucleic acid was extracted by the method of Schneider (1945). DNA and RNA were estimated by diphenylamine (Dische, 1951) using orcinol reaction for RNA (Dishce and Shwartz, 1951). Packed peritoneal cells lysate were used to measure Caspase-3 activity using sigma (St Louis, MO, USA) reagent kit (Nicholson, 1995). Total Genomic DNA was extracted and analyzed on agarose gel (Sambrook and Russell, 2001).
Statistical Analysis
Values are expressed as means±SD of 6 animals in each group. Data within the groups were analyzed using paired t-test to determine the significance. A value of p<0.05 was considered statistically significant.
RESULTS
Effect of Snake Venom on Mean Survival Time and Tumor Growth
The toxicity assay of Hydrophis spiralis venom on the albino mice was used to arrive at sublethal concentrations required for the present study. Based on the values, three different concentration of the snake venom was fixed (1, 10 and 15% of LD50 values as given in the experimental design). The EAC bearing mice showed increased body weight due to the increased ascites volumes by actively proliferating peritoneal cells. Whereas, the bodyweight was decreased from 40.20 to 30.16 g in treated groups. In the EAC control group, the mean survival time was 16.0 days, whereas, it increased significantly in all treated groups. A better effect was observed in medium dose treatment with an increase in the life span by 182.81%, when compared to the carcinoma control group. Increased life span and decreased bodyweight resulted in the decrease tumor volume, packed cell volume and viable tumor cell count in a dose dependent manner (Table 1).
Effect of Snake Venom on Hematological Parameters
Table 2 shows that hemoglobin content was decreased by 2.9% and RBC count decreased by approximately 1.7 fold in EAC control group when compared to normal group. The total WBC counts were found to be increased in EAC control group by 2.6 fold when compared to normal mice.
| Table 1: | Effect of snake venom on body weight changes and survival time |
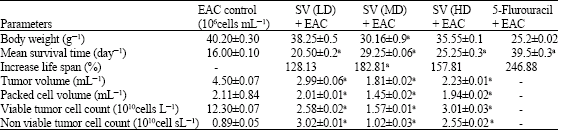 | |
| Values are mean±SD, n = 6, a: p<0.01 vs EAC control group, Body weight of normal mice 22±2 g | |
| Table 2: | Effect of snake venom on Hematological parameters of EAC bearing mice |
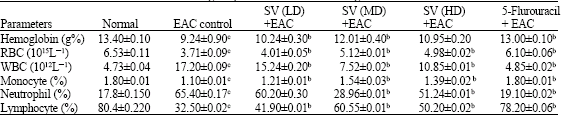 | |
| Values are mean±SD, n=6, cp<0.05 normal group vs EAC control group bp<0.05 treated vs EAC control | |
| Table 3: | Effect of liver marker enzymes AST, ALT and ALP in EAC group and treated groups compared to normal control |
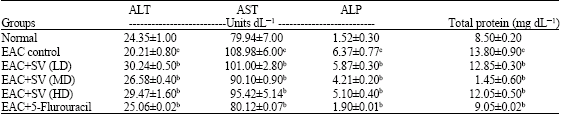 | |
| Unit s = μmoles product released per minute, Values are mean±SD, n = 6, cp<0.05 normal group vs EAC control group bp<0.05 treated vs EAC control | |
| Table 4: | Effect of snake venom on Total DNA, RNA, protein and Capsase-3 in the EAC bearing mice and treated groups |
 | |
| Values are mean±SD, n = 6, c p<0.05 normal group vs EAC control group bp<0.05 Treated vs EAC control | |
In differential count of WBC, neutrophils increased, in contrast lymphocyte count decreased in EAC bearing mice. The above altered levels were brought back to near normal level and more significant results (hemoglobin-12.01%, RBC-5.12x1015 L-1 and WBC 7.52x1012 L-1) were observed with the medium dose of venom when compared to the other treated groups.
Effect on Liver Marker Enzymes
The serum ALT activity was decreased in the EAC mice by 4 units whereas; the activities of AST (by 29 units) and ALP (by 4.8 units) were increased when compared to the normal. The treatment with different doses of snake venom effectively restored the activities of the above enzyme and reduced the burden to the liver caused by the peritoneal EAC cells. In these parameters, again the medium dose (ALT-increased by 3 units, AST-by 1 units and ALP-2units) was found to be more significant when compared to the other treated groups (Table 3).
Effect of Snake Venom on the Total DNA, RNA and Caspase-3 Activity
The selective cytotoxicity of snake venom is demonstrated by a significant reduction of total DNA (4.12 μg 10-6 cells), RNA (7.25 μg 10-6 cells) and protein content (61.2 μg 10-6 cells) of EAC cells in the peritoneal cavity of the treated groups when compared to the EAC tumor control (Table 4). But the caspase-3 activity (4.58 units in the medium dose) was increased in the cell lysate of the treated groups when compared to the EAC mice. All the three treated groups showed good changes in the above parameters and better change was observed with the medium dose of the snake venom.
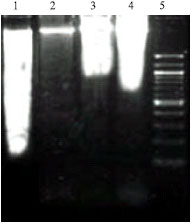 | |
| Fig. 1: | DNA fragmentation assay of EAC bearing mice and treated groups. Lane 1: DNA of EAC + Snake venom (MD); Lane 2: DNA of peritoneal cells from normal mice; Lane 3: DNA of EAC + Snake venom (LD); Lane 4: DNA of EAC + Snake venom (HD); Lane 5: 100 bp DNA ladder |
DNA Fragmentation
Figure 1 shows the agarose gel electrophoresis of chromosomal DNA isolated from the EAC cell of tumor and treated mice. Lane 5 shows the marker, 100 bp DNA ladder. Lane 1 shows the fragmented DNA of the medium dose of snake venom compared to the intact DNA on the lane 2. Lane 3 and 4 shows the DNA of low and medium dose treatment, respectively. Here lane 1 clearly demonstrates the effective DNA damage caused by the medium dose of snake venom when compared to the other two doses.
DISCUSSION
The present study was carried out to evaluate the antitumor efficacy of the snake venom-Hydrophis spiralis. A reliable criteria for assessing the potential of any anticancer agent is the prolongation of life span of the animals (Hogland, 1982). A decrease in tumor volume and viable tumor cell count observed by us can be considered as an important marker of reduced tumor burden and enhanced the lifespan of EAC bearing mice. Andreani et al. (1983) has suggested that an increase in the lifespan of ascites bearing animals by 25% is considered as indicative of significant drug activity. This suggests that the cytotoxic activity of snake venom on the EAC cells might be due to mechanisms other than direct cytolytic effect. In this context, several authors have reported the anti cancer potential of snake venom and exact mechanism of them are not still understood (Abu-Sinna et al., 2003). Lipps (1994) have suggested that venoms act directly on the tumor cells and cause their lysis. But Markland et al. (2001) has suggested that venoms act indirectly by destroying the microenvironment produced by the tumor cells.
In order to understand the mechanism of antitumor effect by snake venom, the main apoptotic marker, Caspase 3 was estimated in the peritoneal cells. Caspases are the central executioners of the apoptotic pathway. They bring about most of the visible changes like cell shrinkage, blebbing of plasma membrane, nuclear condensation and cell death which characterize the apoptosis (Nancy and Luzebnik, 1998). Caspase is particularly activated during apoptosis, which cleave substrates such as poly (ADP-ribose) polymerase, actin and lamin (Hengartner, 2000; Porter and Janicke, 1999). Caspase 3 activity was increased in tumor cell lysate of snake venom treated mice when compared to the EAC bearing mice. The increase was significant in the medium dose of treatment when compared to the other two doses. This suggests that, snake venom can destroy the existing tumor cells by activating the apoptotic cascade at a specific concentration of 4.18 mg kg-1 b.w. of mice.
Willey (1980) has suggested that, during apoptosis a specific nuclease cuts the genomic DNA between nucleosomes to generate DNA fragments and the presence of this ladder has been extensively used as a marker for apoptotic cell death. Nagata (2000) showed that, the DNA ladder nuclease (now known as caspase activated DNase or CAD) pre-exists in living cells as an inactive complex with an inhibitor subunit. Activation of CAD occurs by means of caspase 3 mediated cleavage of inhibitory subunit, resulting in the release and activation of the catalytic subunit. Hence present results, the increased caspase 3 activity, decreased DNA, RNA, protein content and DNA fragmentation (Fig. 1) of the peritoneal cells strongly suggest that snake venom activates the apoptotic cascade and implements the antitumor activity on the EAC cells. However, further studies are warranted for better understanding of snake venom activated apoptotic cascade.
In cancer chemotherapy, other major problems are myelosuppression and anemia (Price and Greenfeild, 1954). The anemia encountered in tumor bearing mice is mainly due to reduction in RBC or hemoglobin percentage or both, which occur either due to iron deficiency or due to hemolytic or myelopathic conditions (Sinclair et al., 1990). Treatment with diluted snake venom brought back the hemoglobin content, RBC and WBC count to near normal. This indicates that snake venom posse’s protective action on the hematopoietic system and the reduced tumor burden has reverted the blood cell count to near normal levels.
Since liver is considered to be the main organ of drug activation, detoxification and other metabolic reactions; activity of some liver marker enzymes and total protein content were measured in the plasma. Innoculation of EAC cells into the mice significantly reduced the activity of ALT and increased the activity of serum AST and ALP. Present findings are in line with the previous reports of Greengard (1979). Yet no explanation has been given for such changes in the activities of these enzymes. But Abu-Sinna et al. (2003) has suggested that, the consumption of free amino acids in building the proteins of rapidly dividing tumor cells might result in the disturbances of the enzyme activity in the liver. Treatment with the snake venom ameliorated these changes to near normal levels indicating the protective role on liver.
In conclusion, the diluted snake venom exhibited a significant antitumor activity against EAC cells and detrimental toxicity on the liver of the treated animals. Hence snake venom is an excellent anticancer agent, which acts specifically on tumor cells by activating the apoptosis cascade.
ACKNOWLEDGMENTS
Sincere appreciation is extended to the authorities of DOD-COMAPS, Center of Advanced Study in Marine Biology, Parangipettai and Annamalai University for providing fellowship and facilities to perform this research. We would also like to acknowledge the Chief conservator of forests and Chief wildlife warden for according permission for the collection of sea snakes.
REFERENCES
- Abdullaev, F.I., R. Rivera-Luna, V. Roitenburd-Belacortu and J. Espinosa-Aguirre, 2000. Pattern of childhood cancer mortality in Mexico. Arch. Med. Res., 31: 526-531.
PubMed - Abu-Sinna, G., A.M. Esmat, S. Al-Zahaby, N.A. Soliman and T.M. Ibrahim, 2003. Fractionation and characterization of Cerastes cerastes cerastes snake venom and the antitumor action of its lethal and non-lethal fractions. Toxicon, 42: 207-215.
Direct Link - Ali, S., J.M. Alam, S. Stoeva, J. Schutz, A. Abbasi, Z.H. Zaidi and W. Voelter, 1999. Sea snake Hydrophis cyanocinctus and N-terminal sequence of two phospholipases A2. Toxicon, 37: 1505-1520.
Direct Link - Andreani, A., G. Scapini, I. Galatulas and R. Bossa, 1983. Potential antitumor agents IX: Synthesis and antitumor activity of two analogues of ketocaine. J. Pharmaceut. Sci., 72: 814-815.
CrossRefDirect Link - Araya, C. and B. Lomonte, 2007. Antitumor effects of cationic synthetic peptides derived from Lys49 phospholipase A2 homologues of snake venoms. Cell Biol. Int.
Direct Link - Hengartner, M.O., 2000. The biochemistry of apoptosis. Nature, 407: 770-776.
CrossRefPubMedDirect Link - Hoagland, H.C., 1982. Hematologic complications of cancer chemotherapy. Semin. Oncol., 9: 95-102.
PubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Maclimans, W.F., E.V. Davis, F.L. Glover and G.W. Rake, 1957. The submerged culture of mammalian cells. The spinner culture. J. Immunol., 79: 428-433.
Direct Link - Markland, F.S., K. Shieh, Q. Zhou, R. Sherwin, V. Richters and R. Sposto, 2001. A novel snake venom disintegrin that inhibits human ovarian cancer dissimenation and angiogenesis in an orthotopic nude mouse. Haemostasis, 31: 183-191.
Direct Link - Mora, R., B. Valverde, C. Diaz, B. Lomonte and J.M. Gutierrez, 2005. A Lys49 phospholipase A2 homologue from Bothrops asper snake venom induces proliferation, apoptosis and necrosis in a lymphoblastoid cell line. Toxicon, 45: 651-660.
Direct Link - Thornberry, N.A. and Y. Lazebnik, 1998. Caspases: Enemies within. Science, 281: 1312-1316.
CrossRefPubMedDirect Link - Nicholson, D.W., 1995. Identification and inhibition of the ICE/CED-3 protease necessary for mammalian apoptosis. Nature, 376: 37-43.
Direct Link - Porter, A.G. and R.U. Janicke, 1999. Emerging role of caspase-3 in apoptosis. Cell Death Differ., 6: 99-104.
Direct Link - Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Sun, P., X. Da Ren, H.W. Zhang, X.H. Li, S.H. Cai, K.H. Ye and X.K. Li, 2003. Serum from rabbit orally administered cobra venom inhibits growth of implanted hepatocellular carcinoma cells in mice. World J. Gastroenterol., 9: 2441-2444.
Direct Link - Yeh, C.H., H.C. Peng, R.S. Yang and T.F. Huang, 2001. Rhodostomin, a snake venom disintegrin, inhibits angiogenesis elicited by basic fibroblast growth factor and suppresses tumor growth by a selective αv β3 blockade of endothelial cells. Mol. Pharmacol., 59: 1333-1342.
Direct Link - Wyllie, A.H., 1980. Glucocorticoid-induced thymocyte apoptosis is associated with endogenous endonuclease activation. Nature, 284: 555-556.
CrossRefDirect Link