
Krishnamurthi Veena
Department of Medical Biochemistry,
Palanivelu Shanthi
Department of Pathology, DR A.L. Mudaliar Post-Graduate Institute of Basic Medical Sciences, University of Madras, Taramani Campus, Chennai-600 113 India
Panchanatham Sachdanandam
Department of Medical Biochemistry,
International Journal of Cancer Research
Year: 2006 | Volume: 2 | Issue: 4 | Page No.: 315-329







ABSTRACT
The effect of Kalpaamruthaa (KA), a modified indigenous preparation, was studied in DMBA induced mammary carcinoma. Glycoprotein components are a biochemical indicator selectively produced by the neoplastic tissue and released into blood and detected in blood or in other body fluids. ATPases is lipid depended membrane bound enzymes and alterations in membrane lipid environment may affect the activities of ATPases and in turn normal cellular functions. Cancerous animals showed a significant decrease in their body weights and significant increase in tumour weights. The levels of Carcino Embryonic Antigen (CEA) in cancer rats were significantly higher, when compared to that of control rats. The levels of these glycoproteins in cancer animals were significantly increased when compared with control animals. The levels of erythrocyte membrane, liver and kidney ATPases are significantly decreased in cancer condition when compared with control animals. Treatment of KA reverted back the pathologic condition by decreasing the level of above enzymes more effectively than Semecarpus anacardium Linn. Nut milk extract (SA). No significant changes were observed in drug control animals when compared with control animals in all the above studies. These evidences suggest that the therapeutic efficacy of the KA was increased on amalgamation of Semecarpus anacardium, Emblica officinalis and honey.
PDF Abstract XML References
How to cite this article
Krishnamurthi Veena, Palanivelu Shanthi and Panchanatham Sachdanandam, 2006. Protective Effect of Kalpaamruthaa on Altered Glycoprotein Component Levels and Membrane Stability In Mammary Carcinoma. International Journal of Cancer Research, 2: 315-329.
DOI: 10.3923/ijcr.2006.315.329
URL: https://scialert.net/abstract/?doi=ijcr.2006.315.329
DOI: 10.3923/ijcr.2006.315.329
URL: https://scialert.net/abstract/?doi=ijcr.2006.315.329
INTRODUCTION
Cancer in various forms is one of the leading causes of death in the human population. Chemotherapy has been employed in reducing human mortality due to cancer or in the control of cancer. There are several natural products of plant origin (phytochemicals) that are of potential value as chemopreventive or therapeutic agents. As therapeutic agents, the phytochemicals kill the cancer cells or stop their growth, leading to cancer remission (Miranda et al., 1999). However, using the incidence of cancer as an evaluation end-point takes a long period and is expensive.
Metastasis is the most life-threatening aspect of cancer and is responsible for the high mortality rates among cancer patients. Tumor metastasis involves a complex series of biochemical interactions during which cancerous cells escape from the primary tumor, migrate and form distant metastases elsewhere at target organs. Tumour cell glycoproteins may be involved in the induction of tumour immunity or pave way for the tumours to escape from immunological defense mechanisms. Cell surface glycoprotein antigens may determine the ability of cancer cells to survive and metastasize. Accelerated degradation or decreased synthesis of terminal cell surface carbohydrate groups may result in the exposure, of antigen on breast carcinoma cells, which are normally hidden or “cryptic” on benign or normal breast epithelium. Glycoproteins are involved in cell interaction like reduced cell adhesiveness, increased degrade active capabilities, altered motility and altered antigenicity. During malignancy, glycoproteins act as antiadhesive molecules that retard T-lymphocyte interactions. The carbohydrate moiety composed of significant amount of sialic acid, contributes to this function. Thus, cell surface carbohydrates act as modulators of immune function (Fukuda, 1991).
Membrane associated carbohydrate is exclusively in the form of oligosaccharides covalently attached to proteins forming glycoproteins. Glycoproteins on cell surfaces are important for communication between cells, for maintaining cell structure and for self-recognition by the immune system. The alteration of cell-surface glycoproteins can therefore, produce profound physiological effects. These include presentation (or masking) of molecular determinants for recognition by cells, microorganisms, or other biological molecules, modulation of the half-life of glycoproteins and physical/structural roles (Hardy, 1989).
The cell surface glycoproteins have been found to play an important role in pinocytosis, tumourigenesis and as mediators of immunological specificity (Hughes, 1976). Carbohydrate moieties of glycoproteins have been implicated in the transport of metabolites across cell membranes and also a direct relation between glycoproteins and tumourigenesis is observed (Glick et al., 1980). Tumour markers correlate strongly with prognosis based on tumour burden and surgical respectability. If chemotherapy is extremely effective in certain stage of the disease, the sensitive marker may be of great use in monitoring disease response and drug treatment (Mujagic et al., 2004). These tumour markers are most useful for monitoring response to therapy and detecting early relapse.
Over expression of glycoproteins is important for tumour progression. They act as anti-recognition factors and in some cases, of the transmembrane component to serve as a growth factor, which can provide a mechanism for autonomous tumour cell proliferation. Carbohydrate variations in tumours are normally explained by variations in glycosyltransferase activities by normal and transformed cells.
A wide group of ATPases is localized on biological membranes. ATPases are exitical for cellular viability because they control many essential cellular functions. ATPase is lipid depended membrane bound enzymes and alterations in membrane lipid environment may affect the activities of ATPase and in turn normal cellular functions (Suzuki et al., 1991). Activities of ATPase indicate active transport system that may be responsible for the active transport of Na+, K+, Ca+ and Mg2+ across cell membranes at the expense of ATP. It’s also vulnerable to oxy radical induced damage and lipid peroxidation (Popp and Schimmer, 1991; Baader et al., 1994). Peroxidation of membrane lipid initiates the loss of membrane integrity and membrane bound enzymes activity which in turn leads to a disruption in cellular homeostatis (Spielholz et al., 1997). Abnormal lipid peroxides affects membrane bound ATPase activities and their levels were decreased due to excessive production of thiobarbituric substances (Langemann et al., 1989).
Plants form an important source of novel chemical compounds with medicinal properties, many of which have been used for prevention and treatment of a variety of human ailments from time immemorial. Experimental and epidemiological studies over the past few decades have provided ample evidence in support of associations between plant food intake and reduced cancer risk. Many phytochemicals are proven to have anticancer activities and many are in use for cancer therapeutics (Miranda et al., 1999). People consuming diets rich in fruits and vegetables have lower incidences of diseases such as cancer (Ziegler, 1991).
Kalpaamruthaa (KA) is a modified indigenous formulation, which has been formulated in our laboratory. KA contains Semecarpus anacardium Linn nut milk extract, dried fruit powder of Emblica officinalis and honey. KA has been found to possess various components such as flavonoids, ascorbic acid, tannins, minerals, sterols, sugars etc., Dose dependent study of Kalpaamruthaa was carried out and it is fixed at 300 mg kg-1 body weight (Veena et al., 2006a). Toxicity study of Kalpaamruthaa showed that there was no toxic upto 2000 mg kg-1 body weight. Effect of KA on reactive oxygen/nitrogen species and lipid metabolism of mammary carcinoma was carried out (Veena et al., 2006b, c). Flavonoids are naturally occurring low molecular weight polyphenolic compounds widely distributed in fruits, vegetables and beverages. Numerous experimental studies have examined the role of specific flavonoids in therapy for diseases. Multiple biological effects of flavonoids have been described, among them, anti-inflammatory; anti-allergic; anti-hemorrhagic; antimutagenic; anti-neoplastic and hepatoprotective activities (Hertog et al., 1993; Vijayalakshmi et al., 2000; Premalatha et al., 1997).
In addition to SA, Emblica and honey are added. Amalgamation of these compounds gives the extra protective effect to the drug. The aim of research is to analyse the effect of Kalpaamruthaa on glycoprotein components and erythrocyte membrane stability in experimental mammary carcinoma. In the present investigation, the glycoprotein components and ATPases in plasma, liver and kidney were measured. Protein, albumin/globulin ratio and Carcinoembryonic Antigens (CEA) in serum were analysed. Changes in body weight and tumour weights were also studied in control and experimental animals.
MATERIALS AND METHODS
Chemicals and Drug
Mammary carcinoma was induced in 8-week old rats by gastric intubation of 7,12-dimethylbenz(a)anthracene (Sigma St Louis, MO; 25 mg mL-1) (Sujatha et al., 1991). The drug (Semecarpus anacardium) preparation was as described in the Formulary of Siddha Medicine (1972). Amalgamation of Semecarpus anacardium Linn. dried powder of Emblica officinalis and honey were added with different ratio.
Animals
Female albino Sprague-Dawley rats, 50-55 days of age, were obtained from the National Institute of Nutrition, Hyderabad and maintained at 27°C (approx.) on a 12±1 h day-night rhythm, given a commercial diet and water (ad libitum).
Experimental Design
The rats were divided into five groups with six animals in each group. Group I: Control animals. Group II: Breast cancer was induced in overnight fasted animals by a single dose of DMBA in olive oil (25 mg kg-1 body weight) by gastric intubation. Group III: Breast cancer induced animals (as in Group II) were treated with the drug, Kalpaamruthaa (300 mg kg-1 body weight/day) dissolved in olive oil orally by gastric intubation for 14 days. Group IV: Breast cancer induced animals (as in Group II) were treated with the drug, Semecarpus anacardium (200 mg kg-1 body weight/day) dissolved in olive oil orally by gastric intubation for 14 days. Group V: Drug treated control, the drug Kalpaamruthaa (300 mg kg-1 body wt.) was administered to control animals for 14 days by gastric incubation.
Experimental Procedure
Body weight and tumour weights were also recorded initially and throughout the end of experimental period. After the experimental period, the animals were sacrificed by decapitation. Liver and kidney were removed immediately, washed well with ice-cold saline and homogenized in Tris-HCI buffer (0.1 M, pH 7.4). Blood was collected for further analyses. The procedures used in this study were approved by Institutional Animal Ethics Committee.
Biochemical Assays
Total protein and albumin were estimated by the method of Reinhold (1953). Globulin content was calculated as the difference between these two values. Hydrolysis of glycoprotein for hexose and hexosamine determination was carried out by a known amount of defatted tissue taken in test tube to which 1 mL of 2N HCl was added and the tubes were sealed. The sealed tubes were kept in 100°C over for 16-18 h for complete hydrolysis. After hydrolysis, the contents were neutralized with sodium hydroxide and made upto a known volume and aliquots were used for hexose and hexosamine determination. Hexose and hexosamine in the aliquots were determined by the method of Niebes (1972) and Wagner (1972), respectively. Analysis of sialic acid was done by a known amount of defatted tissue, hydrolyzed with 0.1N at 90°C and neutralized. The hydrolyzed extract was used for the determination of sialic acid by the method of Warren (1959). CEA was measured by using UBI MAGIWELL enzyme immuno assay Kit (Cianga and Suliman, 1998).
Erythrocyte membrane was isolated according to the method of Dodge et al. (1963) with a change in buffer, according to Quist (1980). Na+, K+-ATPase was measured by the method of Bonting (1970). The activity of Ca2+- ATPase was assayed according to the method of Hjerten and Pan (1983). The activity of Mg2+- ATPase was measured by the method of Ohnishi et al. (1962).
Histological evaluation was performed on mammary gland specimens fixed in 7% formalin and embedded in paraffin wax. Sections were cut at 4 μm thickness, stained with hematoxylin and eosin and viewed under light microscope for histological changes. The slides were examined and scored by a pathologist.
Statistical Analysis
Statistical significance between the experimental groups was assessed by using one-way analysis of variance and by use of Students t-test. Results were presented as Mean±SD of six rats.
RESULTS
Table 1 shows the levels of albumin, globulin and albumin/ globulin (A/G) ratio in control and experimental groups. Significantly decreased levels (p<0.05) of albumin and A/G ratio and significantly (p<0.05) increased levels of globulin content (p<0.05) were observed in cancer bearing animals when compared to control animals. On drug (SA and KA) administration the levels were significantly (p<0.05) increased and decreased, respectively when compared to cancer animals. KA was found to be significantly (p<0.05) more effective when compared to SA. Drug control animals did not show any significant changes when compared with control animals.
Cancerous animals showed a significant decrease in their body weights and significant increase in tumour weights (Fig. 1a-c). The above pathological changes were reverted back to near normal conditions on administration of the drug (SA and KA). KA was observed to be more effective than SA.
The levels of CEA in cancer rats were significantly higher (p<0.05) when compared to that of control rats (Fig. 2). Treatment of KA reverted back the pathologic condition by decreasing the level of CEA more effectively than SA. No significant changes were observed in drug control animals when compared with control animals in all the above studies.
| Table 1: | Effect of KA on levels of albumin, globulin and albumin/globulin ratio in serum of control and experimental animals |
 | |
| Values are expressed as mean±SD for 6 animals. Comparisons are made between: a-when compared with Groups I; b-when compared with Groups II; c-when compared with Groups III. Statistical significance are expressed as *-p<0.05 and NS- Non-significant | |
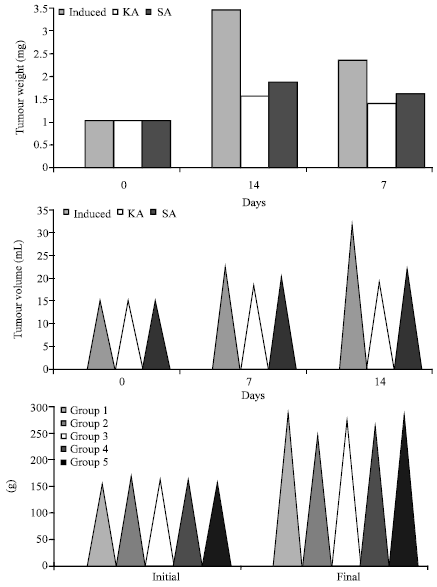 | |
| Fig. 1: | Levels of (a) tumour weight, (b) tumour volume and (c) body weight changes in mammary carcinoma induced and treated rats |
The levels of these glycoproteins in cancer animals were significantly increased when compared with control animals (p<0.05). On drug administration (SA and KA), the levels were brought back to near normal levels. KA more significantly (p<0.05) suppressed the levels of glycoproteins when compared with SA treated animals (Fig. 3, Table 2 and 3).
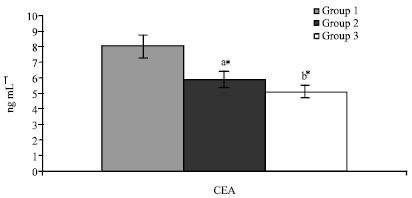 | |
| Fig. 2: | Effect of KA on CEA levels in experimental animals, values are expressed as mean±SD for 6 animals. Comparisons are made between: a-when compared with Groups I; b-when compared with Groups II; Statistical significance are expressed as: *- p<0.05 and NS-Non-significant. Groups I: Tumour induced; Groups II: SA treatment; Groups III: KA treatment |
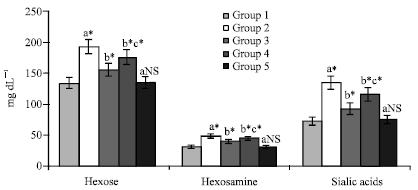 | |
| Fig. 3: | Effect of KA on glycoprotein components in plasma of control and experimental animals, Values are expressed as mean±SD for 6 animals. Comparisons are made between: a-when compared with Groups I; b-when compared with Groups II; c-when compared with Groups III. Statistical significance are expressed as: *- p<0.05 and NS- Non-significant |
| Table 2: | Effect of KA on glycoprotein components in liver of control and experimental animals |
 | |
| Values are expressed as mean±SD for 6 animals. Comparisons are made between: a-when compared with Groups I; b-when compared with Groups II; c-when compared with Groups III. Statistical significance are expressed as: *- p<0.05; and NS-Non-significant | |
| Table 3: | Effect of ka on glycoprotein components in kidney of control and experimental animals |
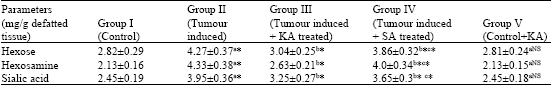 | |
| Values are expressed as mean±SD for 6 animals. Comparisons are made between: a-when compared with Groups I; b-when compared with Groups II; c-when compared with Groups III. Statistical significance are expressed as: *p<0.05 and NS- Non-significant | |
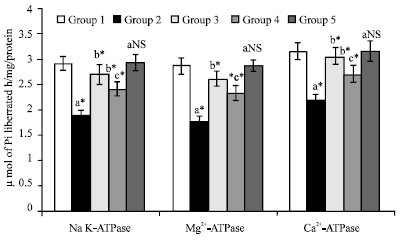 | |
| Fig. 4: | Effect of KA on the changes in erythrocyte membrane adenosine triphosphatase levels in control and experimental animals, Values are expressed as mean±SD for 6 animals. Comparisons are made between: a-when compared with Groups I; b-when compared with Groups II; c-when compared with Groups III. Statistical significance are expressed as: *p<0.05 and NS- Non-significant |
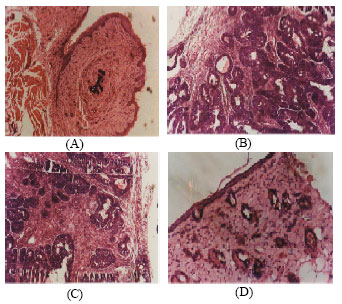 | |
| Fig. 5: | Histopathological changes in breast tissue, (A) Control: Shows the normal architecture of the mammary gland (B) Mammary carcinoma: Pleomorphism with increased mitotic activity. Cluster of multiple group of adenoma cells, irregular in size and shape (C) Mammary carcinoma + KA: Fatty tissue with small lobules, tumour regression is seen (D) Drug control: Kalpaamruthaa alone treated rats showing normal breast tissue structure |
Figure 4 shows the activities of ATPases namely Na+, K+-ATPase; Ca2+- ATPase; Mg2+- ATPase; in erythrocyte membrane of control and experimental animals. Table 4 and 5 shows the levels of Na+, K+-ATPase; Ca2+- ATPase; Mg2+- ATPase; in liver and kidney of control and experimental animals. The levels of erythrocyte membrane, liver and kidney ATPases are significantly (p<0.05) decreased in cancer condition when compared with control animals. On drug administration (SA and KA) the levels were brought back to near normal levels.
| Table 4: | Effect of KA on ATPases in liver of control and experimental animals |
 | |
| Values are expressed as mean±SD for 6 animals. Comparisons are made between: a-when compared with Groups I; b-when compared with Groups II; c-when compared with Groups III. Statistical significance are expressed as: *- p<0.05; and NS-Non-significant | |
| Table 5: | Effect of KA on ATPases in kidney of control and experimental animals |
 | |
| Values are expressed as mean±SD for 6 animals. Comparisons are made between: a-when compared with Groups I; b-when compared with Groups II; c-when compared with Groups III. Statistical significance are expressed as: *- p<0.05; and NS-Non-significant | |
But the effect of KA was more significant (p<0.05) when compared with SA treated animals. No significant changes were noticed in drug control animals when compared with control animals.
Figure 5 shows the histopathological changes in breast tissue. A-represents the normal architecture of the mammary gland (control). B-pleomorphism with increased mitotic activity. Cluster of multiple group of adenoma cells, irregular in size and shape (mammary carcinoma). C- Fatty tissue with small lobules, tumour regression is seen (treatment). D- Kalpaamruthaa alone treated rats showing normal breast tissue structure (drug control).
DISCUSSION
Glycoproteins include presentation (or masking) of molecular determinants for recognition by cells, microorganisms, or other biological molecules, modulation of the half-life of glycoproteins and physical/structural roles. In breast carcinoma, the levels of hexose were significantly increased when compared with their controls (Patel et al., 1990), which are useful indicators of carcinogenic process and these changes alter the structure, rigidity and function of cell membrane (Selvam and Nagini, 1995). Glycoconjugates containing sialic acid are found as cell-specific markers throughout the body. They play a central role in fundamental physiological and pathological processes, e.g., embryogenesis, organogenesis, immune defence, migration and homing of leucocytes, metastasization of neoplastic cells and inflammatory processes, but also in the penetration of cells by pathogens.
Hexosamine elevation in cancerous condition may be due to the damage of connective tissues in mammary tumour malignancy (Winzler, 1953). The glycosyltransferases responsible for synthesis of carbohydrates that play key roles in immune function and human disease are targets for development of novel therapeutic agents.
Sialic acid is an acylated derivative of neuraminic acid and exists as a terminal component of the non-reducing end of carbohydrate chains of glycoprotein in mammals. Sialic acid act as a tumour marker which should be examined from the perspective of aberrant glycosylation in cancer cell membranes owing to activation of new glycosyl transferases that are characteristic of tumour cells and the role played by sialic acid in tumour cell metastasis including increased capacity to adhere to vascular endothelium and decreased capacity of cancer cells to be destroyed by host defense mechanisms. The high sensitivity of sialic acid as a tumour marker has been reported in a variety of cancerous conditions (Narayanan, 1994). Sialic acid measurements have value in monitoring cancer patients during treatment.
The amount of sialic acid in serum was compared to serum carcinoembryonic antigen (CEA) values. Significantly elevated serum sialic acid concentrations were found in breast cancer and showed correlation with tumour stage. The results suggest that measurement of serum sialic acid concentrations may be of adjunctive value in assessing tumour stage (Phan et al., 2004).
We have observed increased levels of glycoprotein such as hexose, hexosamine and sialic acid in mammary carcinoma bearing animals. Altered levels of protein bound carbohydrate are well documented during neoplastic diseases (Shetlar et al., 1950). This change in surface carbohydrate during cellular differentiation and neoplastic transformation suggests their importance in physiology and behaviour of the cells. Such changes have long been implicated in malignant transformations (Hynes, 1976).
On drug treatment, glycoprotein component levels were reverted back to near normal levels. This may be due to the presence of flavonoids, which are proved to possess inhibitory action against carcinogenesis (Elangovan et al., 1994) and cytostabilizing property of the drug. The alkaloids and other components of the drug may alter the expression of glycosyltransferases and interfere with cell membrane glycoprotein synthesis and structure, indicating its potent antitumour property. The reduction in the levels of glycoprotein components indicates that the drug has the ability to suppress malignancy by modulating cell transformation, decreasing the degree of metastases, inhibiting the progression of tumour growth, controlling the cancer cell proliferation and differentiation.
As the cell membrane, very susceptible and sensitive to free radical attack, the enhanced abnormal lipid peroxides, inhibit the activities of ATPases (Rauchova et al., 1995). Inhibited activities of ATPases may also be due to the tumour burden and stress (Cohen et al., 1986), which initially damages the cell membranes. The oxidation of unsaturated fatty acids in biological membranes may cause impairment of membrane function, decreased membrane fluidity, inactivation of membrane bound receptors and enzymes, increase of non-specific permeability to ions and disruption of membrane structure.
Any perturbation in the activities of these enzymes brings about changes in exerting and normal homeostasis (Bean, 1992). Inhibition of ATPase activities in cancerous condition has been reported (Hebell et al., 1986). As they are membrane bound any restrainment to membrane may alter their activities such as membrane fluidity and ionic concentration (Bioj et al., 1973; Jain and Shohet, 1981). The inhibited activities of ATPase may also be due to the cellular damage, because it plays a role in the cancerous condition. So any changes in cell membrane causes disturbance in the activities of membrane bound ATPase thereby indicating the severity of the disease.
Na+, K+, ATPase activity decreased due to alter in the ATPase activity induced by the LPO is partially related to modification of the membrane bound ATPase molecule through changes in lipid protein interaction due to alteration of the lipid bilayer environment around the enzymes. The inhibited enzyme activity may be due to accelerated oxidative stress, which enhanced membrane lipid forming dienes, which ultimately damage the membrane structure and its enzyme activities. In the crucial condition, K+ leakage is enhanced through membrane in which the leakage dependent on the rate of LPO which finally disturb the Na+, K+, ATPase activities (Duticke et al., 1986).
In mammalian cell membrane, the transport system regulates the intra cellular concentration of Na+ and K+. The Na+ and K+ transportation of the cell are carried out by the hydrolysis of ATP against an electrochemical gradient (Jain and Shohet, 1981). Active transport of Na+, K+ directly depends on the active calcium transport and resultant low calcium concentration. Since sodium and calcium are believed to be competitive at number of membrane sites, the higher concentration of Ca2+ in cancer cells may compete with sodium at sodium specific site at the inner surface of the membrane, which may lead to the decrease in sodium content.
Magnesium is one of the intracellular ions and has a key role in the intermediary metabolism. It is a component of the high energy ATP-Mg2+ complex that provides a universal form of energy for the organism. Lipid peroxides inhibit Mg2+ dependent ATPase activity involved in maintenance of cell homeostasis and cell energy production. Mg2+-dependent ATPases represent key enzymes in maintenance of the integrity of cells and are indispensable for their viability.
On drug treatment, elevations were observed in ATPases’ activities. This may be due to the membrane stabilizing property of the drug. Drug contains mainly flavonoids, which are known to influence the permeability of biomembranes and it interacts with Na+, K+-ATPase pumps in animal cells and may have the ability to catalyse electron transport. By this property, the activities of ATPases might have regained their normal efficiency and the cell assumed normal properties (Lee, 1999). These compounds also possess antioxidant and antiradical properties. Antioxidant activity of flavonoids may also be due to their structural features and its action on the membrane (Havstein, 1983). All these cluster of properties may be responsible for the recoupment of ATPases in drug treated animals. Drug control animals did not show any significant changes when compared to control animals, indicating its non-interference with normal cells and cell membranes.
The presence of flavonoids, ascorbic acid, sterols, polyphenols, glycosides in KA, which are responsible for the activity of the enzymes. Flavonoids, including chalcones and flavanones, have been shown to inhibit the proliferation of cancer cells and inhibit tumour growth (Anto et al., 1995). Recording the adenocarcinoma incidence weekly in DMBA induced rats of Sprague-Dawley strain assessed the therapeutic activity of KA. In the initial stage of the induction period, there was no tumour in the animals, after that significantly increased by the next 3 months. The weights were reduced during the treatment period. An extensive literature, including many reviews (Keenan et al., 1996) covers investigations into the extent, mechanisms and applications of the phenomena that energy restriction protects against many of the diseases associated with ageing, including spontaneous tumour formation in rats and mice and promotes longevity.
Part of an animal’s energy expenditure is related to the rates of turnover and metabolism of cellular and intra-cellular systems and to the size and number of metabolically active cells, hence, the importance of body size and composition in the amount of energy expended for basal (i.e. minimal, resting, fasting and unstimulated) metabolic states. Weight loss and tissue wasting were observed in cancer patients. This implies poor prognosis and shorter survival time for cancer patients (Tessitore et al., 1994).
The Carcinoembryonic antigen (CEA) has been used as a tumour marker for breast cancer (Mujagic et al., 2004). CEA is a tumour antigen of less differentiated cancer cells. Down-regulation of carcinoembryonic antigen-related cell adhesion molecule 1 (CEACAM1) tumour suppressor gene expression is common in several malignancies including prostate, colon and breast cancer. The mechanism that mediates this down-regulation is not completely known.
The decrease in albumin levels in DMBA treated rats could be attributed to the impaired hepatic function resulting from infiltration with tumour. The serum albumin level has been used as a test for liver function because, serum protein concentration is controlled by hepatic protein synthesis and also albumin is the most abundant circulatory protein and its synthesis is a typical function of normal liver cells (Mc Intyre and Rosalki, 1973; Waitzberg et al., 1989).
The elevation of globulin content is compensatory as the ratio of albumin to globulin shows a significant drop in the cancer bearing animals. On drug treatment, the levels were brought back to near normal levels. In tumour bearing rats, there was a notable reduction in the body weight. The reason for the reduction in body mass may be due to cancer cachexia which is characterized by the features of anorexia and early satiety, weight loss and marked muscle weakness, anemia of a non-specific type and altered host metabolism (Argiles and Aczon-Bieto, 1988). Other possible mechanism of weight reduction is the acute-phase response and cytokines. Recent researchers explain that cancer cachexia may be a chronic inflammatory condition involving the products of arachidonic acid rather than a nutritional alteration. So food that modulates the synthesis or activity of proinflammatory mediators, especially the synthesis of PGE2 from arachidonic acid can be used in contracting cancer cachexia (Donna and McCarthy, 2003).
Food rich in polyphenols, flavonoids, catechins could mediate favourable changes due to their antioxidant, immunopotentiating and anti-inflammatory properties (Barber et al., 2001). Upon drug administration, there was a gradual increase in body weight, which denotes the antineoplastic nature of the drug Kalpaamruthaa. No mortality was observed with drug control animals and decreased mortality in experimental animals proves the non-toxic nature of the drug. Drug control animals did not show any statistically significant changes in these parameters at quest.
Flavonoids might influence the studied carcinogenic parameters and might be the main tumour inhibitory representative in the drug. Chakraborty et al. (2004) also showed that the plant product acts as inducers of apoptosis in tumour cells and thus acts as a potent anti-proliferative agent and this strategy suggests their potential use in cancer control. The flavonoid may inhibit growth and cause regression of tumours via modulating the protein kinases activity and through induction of apoptosis (Choi et al., 2003). The mechanism of carcinogen inactivation, antiproliferation, cell cycle arrest, induction of apoptosis on differentiation, inhibition of angiogenesis and antioxidation property of flavonoids may be responsible for tumour growth inhibition (Ren et al., 2003).
One of the important functions of the liver cell is the synthesis of albumin, the circulating protein. Its reduction in cancerous animals may be due to the improper functioning of the liver in cancerous conditions. In Morris hepatoma 9121 (Rotermund et al., 1970) and gastrointestinal malignancies (Landel et al., 1985), hypoalbuminemia is observed which may be due to the increased catabolism in cancerous conditions. The elevation of globulin content is compensatory as the ratio of albumin to globulin shows a significant drop in the cancer bearing animals. On drug treatment, the levels were recouped to there near normal levels, which indicates its safe guard property. This could be attributed to the cytoprotective effect of the constituents present in the drug.
The fruits have been reported to contain constituents with variable biological activity. Experiments conducted with the fruit of Amla have been shown to possess antioxidant (Bhattacharya et al., 1999), hepato-protective (Jeena et al., 1999) and anti-tumour activities (Jose and Kuttan, 2001). Amla fruits have long been postulated to be a rich source of vitamin C and the prophylactic, curative and restorative effects of the fruits were thought to be mainly due to this factor. Chrysin is a natural, biologically active compound extracted from honey. It possesses potent anti-inflammation, anti-cancer and anti-oxidation properties. The mechanism by which chrysin suppresses COX-2 expression remains poorly understood. In the present report, the effect of chrysin on the expression of COX-2 in lipopolysaccharide (LPS)-activated Raw 264.7 cells has been investigated (Woo et al., 2005).
This study demonstrates the reduced cancer risk on drug (Kalpaamruthaa) administration. Therapeutic efficacy of the drugs was studied in Membrane stability; membrane bound glycoproteins, A/G ratio and CEA and body and tumour weight changes. Further it should be confirmed by histopathological studies. Amalgamation of these drugs increased the antineoplastic activity of a single drug.
REFERENCES
- Anto, R.J., K. Sukumaran, G. Kuttan, M.N.A. Rao, V. Subbaraju and R. Kuttan, 1995. Anticancer and antioxidant activity of synthetic chalcones and related compounds. Cancer Lett., 97: 33-37.
CrossRef - Argiles, J.M. and J. Aczon-Bieto, 1988. The metabolic environment of cancer. Mol. Cell. Biochem., 81: 3-17.
CrossRef - Baader, S.L., G. Bruchelt, M.C. Trautner, H. Boschert and D. Niethammer, 1994. Uptake and cytotoxicity of ascorbic acid and dehydroascorbic acid in neuroblastoma (SK-N-SH) and neuroectodermal (SK-N-LO) cells). Anticancer Res., 14: 221-227.
PubMed - Barber, M.D., K.C. Fearon, M.J. Tisdale, D.C. McMillan and J.A. Ross, 2001. Effect of a fish oil-enriched nutritional supplement on metabolic mediators in patients with pancreatic cancer cachexia. Nutr. Cancer, 40: 110-124.
Direct Link - Bean, B.P., 1992. Pharmacology and electrophysiology of ATP-activated ion channels. Trends. Pharmacol. Sci., 13: 87-90.
PubMed - Bhattacharya, A., A. Chatterjee, S. Ghosal and S.K. Bhattacharya, 1999. Antioxidant activity of active tannoid principles of Emblica officinalis (Amla). Indian J. Exp. Biol., 37: 676-680.
PubMedDirect Link - Chakraborty, S., M. Roy, A.K. Taraphdar and R.K. Bhattacharya, 2004. Cytotoxic effect of root extract of Tiliacora racemosa and oil of Semecarpus anacardium nut in human tumour cells. Phytother. Res., 18: 595-600.
Direct Link - Choi, Y.J., J.S. Kang, J.H.Y. Park, Y.J. Lee, J.S. Choi and Y.H. Kang, 2003. Polyphenolic flavonoids differ in their antiapoptotic efficacy in hydrogen peroxide-treated human vascular endothelial cells. J. Nutr., 133: 985-991.
Direct Link - Donna, O. and R.N. McCarthy, 2003. Rethinking nutritional support for persons with cancer cachexia. Biol. Res. Nur., 5: 3-17.
CrossRef - Duticke, B., K.B. Heller and C.W.M. Haest, 1986. Leak formation in human Biochem. Biophys. Acta., 854: 169-183.
PubMed - Elangovan, V., N. Ramamoorthy, S. Balasubramanian, N. Sekar and S. Govindasamy, 1994. Studies on the antiproliferative effect of some naturally occurring biflavonoidal compounds against human carcinoma of larynx and sarcoma-180 cell lines. Indian J. Pharmacol., 26: 266-269.
Direct Link - Fukuda, M., 1991. Leukosialin, a major O-glycan-containing sialoglycoprotein defining leukocyte differentiation and malignancy. Glycobiology, 1: 347-356.
Direct Link - Havstein, B., 1983. Flavonoids a class of natural products of high pharmacological potency. Biochem. Pharmacol., 32: 1141-1148.
PubMed - Hebbel, R.P., O. Shalev, W. Foker and B.H. Rank, 1986. Inhibition of erythrocyte Ca2+-ATPase by activated oxygen through thiol and lipid-dependant mechanisms. Biochim. Biophys. Acta, 862: 8-16.
PubMed - Hertog, M.G.L., P.C.H. Hollman, M.B. Katan and D. Kromhout, 1993. Intake of potentially anticarcinogenic flavonoids and their determinants in adults in the Netherlands. Nutr. Cancer, 20: 21-29.
CrossRefPubMedDirect Link - Hjerten, S. and H. Pan, 1983. Purification and characterization of two forms of a low-affinity Ca2+-ATPase from erythrocyte membranes. Biochimica Biophysica Acta (BBA)-Biomembr., 728: 281-288.
CrossRefDirect Link - Hynes, R.O., 1976. Cell surface proteins and malignant transformation. Biochim. Biophys. Acta (BBA)-Rev. Cancer, 458: 73-107.
CrossRefPubMedDirect Link - Jain, K.S. and S.B. Shohet, 1981. Calcium potentiase the peroxidation of erythrocyte membrane lipids. Biochem. Biophys. Acta, 642: 46-54.
PubMed - Jeena, K.J., K.L. Joy and R. Kuttan, 1999. Effect of Emblica officinalis, Phyllanthus amarus and Picrorrhiza kurroa on N-nitrosodiethylamine induced hepatocarcinogenesis. Cancer Lett., 136: 11-16.
PubMed - Jose, J.K., G. Kuttan and R. Kuttan, 2001. Antitumour activity of Emblica officinalis. J. Ethnopharmacol., 75: 65-69.
CrossRef - Keenan, K.P., P. Laroque, G.C. Ballam, K.A. Soper and R. Dixit et al., 1996. The effects of diet, ad libitum overfeeding and moderate dietary restriction on the rodent bioassay: The uncontrolled variable in safety assessment. Toxicol. Pathol., 24: 757-768.
CrossRef - Landel, A.M., W.G. Hammond, M.M. Meguid, 1985. Aspects of amino acid and protein metabolism in cancer bearing states. Cancer., 55: 230-237.
PubMed - Langemann, H., J. Torhorst, A. Kabiersch, W. Krenger and C.G. Honegger, 1989. Quantitative determination of water- and lipid-soluble antioxidants in neoplastic and non-neoplastic human breast tissue. Intl. J. Cancer, 43: 1169-1173.
PubMed - Lee, I.M., 1999. Antioxidant vitamins in the prevention of cancer. Proc. Assoc. Am. Physicians, 111: 10-15.
PubMed - Mirandai, C.L., J.F. Stevens, A. Helmrich, M.C. Henderson and R.J. Rodriguez et al., 1999. Antiproliferative and cytotoxic efects of prenylated flavonoids from hops (Humulus lupulus) in human cancer cell lines. Food Chem. Toxicol., 37: 271-285.
CrossRef - Mujagic, Z., H. Mujagic and B. Prnjavorac, 2004. The relationship between circulating carcinoembryonic antigen (CEA) levels and parameters of primary tumour and metastases in breast cancer patients. Med. Arh., 58: 23-26.
PubMed - Niebes, P., 1972. Determination of enzymes and degradation products of glycosaminoglycan metabolism in the serum of healthy and varicose subjects. Clinica Chimica Acta, 42: 399-408.
CrossRefDirect Link - Ohnishi, T., T. Suzuki, Y. Suzuki and K. Ozawa, 1982. A comparative study of plasma membrane Mg2+-ATPase activities in normal, regenerating and malignant cells. Biochim. Biophys. Acta (BBA)-Biomembr., 684: 67-74.
CrossRefPubMedDirect Link - Patel, P.S., B.R. Baxi, S.G. Adhvaryu and D.B. Balar, 1990. Individual and combined usefulness of lipid associated sialic acid, mucoid proteins and hexoses as tumor markers in breast carcinoma. Cancer Lett., 51: 203-208.
CrossRefPubMedDirect Link - Popp, R. and O. Schimmer, 1991. Induction of sister-chromatid exchanges (SCE), polyploidy and micronuclei by plant flavonoids in human lymphocyte cultures. A comparative study of 19 flavonoids. Mutat. Res., 246: 205-213.
PubMed - Premalatha, B., V. Muthulakshmi, T. Vijayalakshmi and P. Sachdanandam, 1997. Protective role of Serankottai nei, a siddha preparation on cell membranes, in aflatoxin B1 induced hepatocellular cacinoma bearing rats. Indian drugs, 34: 384-389.
Direct Link - Quist, E.H., 1980. Regulation of erythrocyte membrane shape by calcium anion. Biochem. Biophys. Res. Commun., 92: 631-637.
Direct Link - Rauchova, H., J. Ledvinkova, M. Kalous and Z. Drahota, 1995. The effect of lipid peroxidation on the activity of various membrane-bound ATPases in rat kidney. Intl. J. Biochem. Cell Biol., 27: 251-255.
CrossRefPubMedDirect Link - Ren, W., Z. Qiao, H. Wang, L. Zhu and L. Zhang, 2003. Flavonoids: Promising anticancer agents. Med. Res. Rev., 23: 519-534.
CrossRefPubMedDirect Link - Rotermund, H., G. Schreiber, H. Maeno, U. Weinssen and K. Weigand, 1970. The ratio of albumin synthesis to total protein synthesis in normal rat liver in host liver and in Morris hepatoma 9121. Cancer Res., 30: 2139-2146.
Direct Link - Selvam, S. and S. Nagini, 1995. Administration of plasticizer di(2-ethyl hexyl)phthalate alters glycoconjugate profile. Ind. J. Physiol. Pharmacol., 39: 252-254.
Direct Link - Shetlar, M.R., C.P. Erwir and M.R. Evereft, 1950. Serum polysaccharide levels in rats bearing the walker 256 tumour. Cancer Res., 10: 445-447.
Direct Link - Spielholz, C., D.W. Golde, A.N. Houghton, F. Nualart and J.C. Vera, 1997. Increased facilitated transport of dehydroascorbic acid without changes in sodium-dependent ascorbate transport in human melanoma cells. Cancer Res., 57: 2529-2537.
Direct Link - Suzuki, S., T. Takada, Y. Sugawara, T. Muto and R. Kominami, 1991. Quercetin induces recombinational mutations in cultured cells as detected by DNA fingerprinting. Japan. J. Cancer Res., 82: 1061-1064.
PubMed - Tessitore, L., P. Costelli and M.F. Baccino, 1994. Pharmacological interference with tissue hypercatabolism in tumour-bearing rats. Biochem. J., 299: 71-78.
PubMed - Vijayalakshmi, T., V. Muthulakshmi and P. Sachdanandam, 2000. Toxic studies on biochemical parameters carried out in rats with Serankottai nei, a Siddha drug-milk extract of Semecarpus anacardium nut. J. Ethnopharmacol., 69: 9-15.
CrossRef - Veena, K., P. Shanthi and P. Sachdanandam, 2006. The biochemical alterations following administration of Kalpaamruthaa and Semecarpus anacardiumin mammarycarcinoma. Chem. Biol. Interact., 161: 69-78.
PubMed - Veena, K., P. Shanthi and P. Sachdanandam, 2006. Anticancer effect of Kalpaamruthaa on mammary carcinoma in rats with reference to glycoprotein components, lysosomal and marker enzymes. Biol. Pharm. Bull., 29: 565-569.
Direct Link - Veena, K., P. Shanthi and P. Sachdanandam, 2006. Therapeutic efficacy of Kalpaamruthaa on reactive oxygen/nitrogen species levels and antioxidative system in mammary carcinoma bearing rats. Mol. Cell. Biochem., 294: 127-135.
CrossRef - Waitzberg, D.L., E.L. Goncalves, J. Faintuch, A. Bevilacqua, C.L. Rocha and A.M. Cologni, 1989. Effect of diets with different protein levels on the growth of Walker 256 carcinosarcoma in rats. Braz. J. Med. Biol. Res., 22: 447-455.
PubMed - Warren, L., 1959. The thiobarbturic acid assay of sialic acid. J. Biol. Chem., 234: 1971-1975.
Direct Link - Woo, K.J., Y.J. Jeong, H. Inoue, J.W. Park and T.K. Kwon, 2005. Chrysin suppresses lipopolysaccharide-induced cyclooxygenase-2 expression through the inhibition of nuclear factor for IL-6 (NF-IL6) DNA-binding activity. FEBS Lett., 579: 705-711.
CrossRef - Ziegler, R.G., 1991. Vegetables, fruits and carotenoids and risk of cancer. Am. J. Clin. Nutr., 53: 251s-259s.
Direct Link