
Ch. Kalyankumar
School of Biotechnology (MGNIRSA), University of Mysore, India
Kaiser Jamil
Department of Genetics, Bhagawan Mahavir Medical Research Center,
Hyderabad, (500 004) A.P, India
International Journal of Cancer Research
Year: 2006 | Volume: 2 | Issue: 2 | Page No.: 143-151







ABSTRACT
In this study investigations were made to detect the association of MTHFR C677T and A1298C polymorphisms in breast cancer patients, in the South Indian population. Eighty eight women patients with breast cancer and 95 healthy women as controls were included in the study after their consent to participate in this program. MTHFR gene polymorphisms were determined by the PCR-RFLP method. The allele frequencies of the MTHFR C677T were 27.84% in the breast cancer subjects and 19.47% in the controls. The allele frequencies of the MTHFR A1298C were 25.56% in the breast cancer subjects and 17.89% in the controls. Frequencies of MTHFR C677C, C677T and T677T were 47.9%, 44.8%, 7.1% in breast cancer patients and 63.2, 34.5 and 4.6% in the controls. Frequencies of MTHFR A1298A, A1298C and C1298C were 54.8, 40.8 and 5.1% in breast cancer patients and 66.9, 30.1 and 2.9% in the controls. When we compared the frequencies of C677T and T677T with C677C, A1298C and C1298C with A1298A, we found less significant association between specific MTHFR variants in breast cancer patients. Our findings suggest that MTHFR C677T and A1298C gene variants do not have a major influence on the susceptibility to breast cancer in south Indian population.
PDF Abstract XML References
How to cite this article
Ch. Kalyankumar and Kaiser Jamil, 2006. Methylenetetrahydrofolate Reductase (MTHFR) C677T and A1298C Polymorphisms and Breast Cancer in South Indian Population. International Journal of Cancer Research, 2: 143-151.
DOI: 10.3923/ijcr.2006.143.151
URL: https://scialert.net/abstract/?doi=ijcr.2006.143.151
DOI: 10.3923/ijcr.2006.143.151
URL: https://scialert.net/abstract/?doi=ijcr.2006.143.151
Introduction
Breast Cancer (BC) is diagnosed in more than 1 million women and it claims 370,000 deaths worldwide annually (Stewart and Kleihues, 2003). Some 5% of breast cancer is familial, i.e., women with breast cancer have a mother with the same disease. Metabolic imbalances have been speculated to contribute to the risk of cancer. Folate metabolism has been of interest because as it serves as a donor of one-carbon groups in both DNA methylation and nucleotide synthesis. Recent epidemiological studies have implicated imbalances in folate status in the development of cancer at several organs, like breast.
Gene and Gene Product
The 5, 10-methylenetetrahydrofolate reductase (MTHFR) gene is located on chromosome 1 at 1p36.3. The complementary DNA sequence is 2.2-kilo bases long and consists of 11 exons (Goyette et al., 1994). MTHFR is a polymorphic gene involved in folate metabolism, DNA biosynthesis, DNAmethylation and genomic integrity in actively dividing cells.
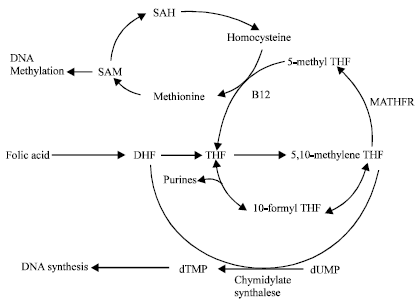 | |
| Fig. 1: | Overview of the human folic acid metabolic pathway and the role of MTHFR, s-adenosylmethionine (SAM), s-adenosylhomocysteine (SAH), dihydrofolic acid (DHF) and tetrahydrofolicacid |
MTHFR is an enzyme that plays a central role in the metabolism of folate. MTHFR catalyzes the conversion of 5, 10-methylenetetrahydrofolate into 5-methyltetrahydrofolate, which is the major circulating form of folate. Folic acid and MTHFR are involved in complex biochemical pathways (Beaudet and Sly, 1995) (Fig. 1). Folate, in its 5- methyl form, participates in single-carbon transfers that occur as part of the synthesis of nucleotides; the synthesis of S-adenosyl-methionine; the remethylation of homocysteine to methionine; and the methylation of DNA, proteins, neurotransmitters and phospholipids. DNA methylation has been suggested as one of the molecular mechanisms by which gene expression is regulated (Jacob et al., 1998) (Fig. 1). For instance, hypomethylation is associated with activation of oncogenes and promoter hypermethylation is associated with loss of function of tumor suppressor genes (Goyette et al., 1994). A recent study (Kim et al., 1997) found that individuals homozygous for the C677T polymorphism have undermethylated genomic DNA in their peripheral leukocytes, a characteristic of many types of cancer, including breast cancer.
MTHFR is highly polymorphic in the general population. Two common polymorphisms, C677T and A1298C, have been identified (Mathews and Rozen, 1994; Vander Put et al., 1998). The nucleotide 677 polymorphism results in an alanine to valine (C-T) substitution (Jacob et al., 1998). Individuals with the variant 677TT genotype have about 30% of the in vitro MTHFR enzyme activity of those with the 677CC wild-type genotype, whereas heterozygotes (677CT) have about 65% of normal enzyme activity (Frosst et al., 1995). Up to 15% of individuals are homozygous 677TT for the variant, which is associated with higher plasma homocysteine levels and reduced plasma folate levels (Deloughery et al., 1996). Among healthy subjects, the C677T genotype is associated with a significantly higher homocysteine and low red cell folate levels than in heterozygote or Individuals with wild type C allele’s Elevated plasma levels of homocysteine are caused by this specific polymorphism, resulting in breast cancer (Gershoni-Baruch et al., 2000; Sharp et al., 2002; Campbell et al., 2002; Semenza et al., 2003; Weisberg et al., 1997). The fact that MTHFR is involved in DNA methylation and the synthesis of uridylate and thymidylate for DNA biosynthesis and repair makes MTHFR a candidate for a susceptibility gene for cancer.
A second MTHFR polymorphism, A1298C in exon 7, results in a glutamate-to-alanine substitution at codon 429 (Van der Put et al., 1998; Stern et al., 2000). This polymorphism lies in the S-adenosylmethionine-regulatory domain of the enzyme The binding of S-adenosylmethionine (SAM) results in conformational changes within the MTHFR enzyme that inhibit the enzyme’s activity (Matthews et al., 1984; Jencks and Mathewes, 1987; Wiemels et al., 2001).
Previous studies (Stampfer et al., 1999) of breast cancer reported a significantly decreased risk of colorectal cancer associated with the 677TT genotype that was not observed among those with low folate intakes or serum levels. However, the association between these two common MTHFR polymorphisms and the risk of breast cancer has not been examined. Because the MTHFR polymorphisms reduce enzyme activity and lower dietary intake of folate is associated with increased risk of breast cancer.
Recent reports have shown that thymidylate deficiencies may result in the misincorporation of uridylate into DNA, thus causing an increased rate of DNA strand breaks and chromosomal damage (Dianov et al., 1991). Reduced MTHFR activity may result in increased levels of cytosolic5, 10-methylenetetrahydrofolate available for thymidylate synthesis and may thus protect cells from DNA damage induced by uridylate misincorporation.
This variant, which is present in the homozygous state in 10-15% of many North American and European populations, correlates with reduced enzyme activity and increased thermolability (Frosst et al., 1995). This decrease in MTHFR activity shifts folate derivatives into the nonmethyl forms. Previous studies have shown a possible link between MTHFR genotype and the folate pool in breast cancer. DNA methylation plays an important role in the regulation of gene expression and maintenance of genomic stability (Kundu et al., 1991; Lengauer et al., 1997) and aberrations in normal methylation patterns have been associated with the development of cancer (Cheng et al., 1997; Stern et al., 2000). The role of MTHFR polymorphism in breast cancer has been studied in which the level of folate in plasma is inversely related to the breast cancer risk.
The aim of this investigation was therefore to evaluate the polymorphism of MTHFR gene at the exon 4 and 7 at position 677 and 1298, respectively and their allele and also to determine if any correlation exits between the polymorphisms and breast cancer. In view of the conflicting reports of the risk of breast cancer to the MTHFR polymorphism in 677 (exon 4) and 1298 (exon 9) we undertook this investigation.
MATERIALS AND METHODS
Study population and sample collection: Breast cancer patients were assessed on the basis of clinical examinations as well as mammography and pathological examinations. The Breast Cancer Study is a population-based case-control study conducted in South India during June 2004-May 2005. A total of 88 breast cancer patients and 95 age matched controls were enrolled in the study.
Inclusion and Exclusion Criteria
All incident breast cancer cases were newly diagnosed during the study period and meeting the following criteria was eligible for this study: 25-64 years age of women, residents of Andhra Pradesh (south India), with no previous history of any cancer. The breast cancer patients studied here had not been exposed to chemo- and/or radiotherapy before. They underwent clinical examinations at the Various Hospitals in Hyderabad. Written informed consent was obtained from all subjects and relevant ethical committees approved the study benefit of humans in general. Two senior pathologists confirmed all diagnoses.
Collection of Biopsy Samples
The biopsy samples from patients who underwent surgery for removal of tumor (breast tumor) were collected from various institutes like Indo American Cancer Institute and M.N.J. Hospital.
Collection of Blood Samples
Blood samples from healthy women were collected by venipuncture. This sample was used as controls.
Genotype Analysis (MTHFR Polymorphism Studies)
DNA Isolation from Breast Tissue and Blood Samples
Genomic DNA was extracted from biopsies (tissues) and blood using standard procedure (Miller et al., 1998). The MTHFR C677T and A1298C polymorphisms were analyzed as previously described (Frosst et al., 1995; Weisberg et al., 1998). Mutations were detected by P.C.R amplification with specific primers, as described below.
C677T Mutation
PCR amplification of Exon 7 of MTHFR was adapted from the method described by Frosst et al. (1995) as follows: Initial denaturation step was for 4 min at 94°C followed by 30 cycles of denaturation for 30 sec at 94°C, annealing for 30 sec at 62°C and elongation for 90 sec at 72°C. The final elongation step was for 7 min at 72°C. Primers used were: primer A, 5'-TGA AGG AGA AGG TGT CTG CGG GA; primer B, 5' AGG ACG GTG CGG TGA GAG TG resulted in a198 bp product. Each primer was used at 0.25 μmol L-1 in a 25 μL final volume in the presence of standard PCR master mix. Amplifications were performed. Amplicons were then digested for at least 2 h (usually 4 h) with HinfI in the buffer supplied by the manufacturer, C677T mutation creates HinfI restriction site causing cleavage of the 198 bp, C677T heterozygotes were identified by 198, 175 and 23 bp fragments and T677T homozygotes were identified by 175 and 23 bp fragments. The wild-type allele (alanine) remains undigested. The products were visualized by ethidium bromide staining (0.5 mg L-1) of a 3% agarose gel.
A1298C Mutation
PCR amplification of Exon 4 of MTHFR was adapted from the method described by Frosst et al. (1995) as follows: Initial denaturation step was for 4 min at 94°C followed by 30 cycles of denaturation for 30 sec at 94°C, annealing for 30 sec at 62°C and elongation for 90 sec at 72°C. The final elongation step was for 7 min at 72°C. Primers used were: 5'-CTC CCT TCA CTT TCA GAA CTA CA; and primer 5’ GAC CTC TCA GTT TTC ACC TTT resulted in a163-bp product each primer was used at 0.25 μmol L-1 in a 25 μL final volume in the presence of standard PCR master mix. Amplifications were performed. Amplicons were then digested for at least 2 h (usually 4 h) with MobII in the buffer supplied by the manufacturer and the products were visualized by ethidium bromide staining (0.5 mg L-1) of a 3% agarose gel. A1298C mutation creates MboII, restriction site causing cleavage of the 163 bp fragments. 1298AA wild-type homozygotes produce five fragments of 56, 31, 30, 28 and 18 bp. The 1298AC heterozygotes produce six fragments of 84, 56, 31, 30, 28 and18 bp and the 1298CC homozygous variants produce four fragments of 84, 31, 30 and 18 bp.
Statistical Analysis
The results of the polymorphic studies in cases and controls evaluated statistically. Odd Ratios (OR) and 95% Confidence Intervals (CI) for matched analysis were computed by using Epi InfoVersion6 (CDC) Yates corrected chi-square (χ2 ) and p-values with a level for each allelic variant were generated for MTHFR C677T and MTHFR A1298C.
RESULTS
Table 1 shows the frequency and allelic distributions of 88 breast cancer patients together with 95 controls. In this study the frequencies were in Yates corrected Chi-square and p-values. The MTHFR C677T allele frequency was 27.84% in the breast cancer patients and 19.47% in the controls, which was not statistically significant (OR = 0.62; 95% CI =0.32-1.19 χ2 = 2.28; p = 0.123). Frequencies of MTHFR C677C, C677T and T677T genotypes were 47.9, 44.8 and 7.1% in breast cancer patients and 63.2, 34.5 and 4.6% in the controls, respectively. The T677T genotype OR = 0.37; 95% CI = 0.17-0.79; χ2 = 1.97; p = 0.160).
The allele frequency of MTHFR A1298C was 25.56% in the breast cancer patients and 17.89% in the controls. Frequencies of MTHFR A1298A, A1298C and C1298C were 54.8, 40.8 and 5.1% in the breast cancer patients and 66.9, 30.1 and 2.9% in the controls, respectively. The MTHFR A1298C allele frequency was 25.56% in the breast cancer patients and 17.89% in the controls, which was statistically significant (OR = 0.59; 95% CI =0.30-1.17 χ2 = 2.61; p = 0.105). The C1298C genotype OR = 0.50; 95% CI = 0.11-2.16; χ2 = 1.08; p = 0.298).
We found the MTHFR C677C genotype present among 45 (51.5%) breast cancer cases and 61 (64.2%) controls, the C677T genotype among 37 (42.0%) breast cancer cases and 31(32.6%) controls and the T677T genotype among 6(6.8%) breast cancer cases and 3 (3.1%) controls. For MTHFR 1298, the A1298A genotype was observed in 49(55.6%) of the breast cancer cases and 65 (68.4%) of the controls, the A1298C genotype was observed in 33(37.5%) breast cancer cases and 26 (27.3%) controls and the C1298C genotype among 6 (6.8%) breast cancer cases and 4 (4.2%) controls. When we compared C677T and C677T with C677C. A1298C and C1298C with A1298A. In this study the results showed a less significant association of these MTHFR variants in breast cancer women.
| Table 1: | Genotype, allele frequencies, OR, 95% CI, χ2 and p values of the C677T and A1298C polymorphisms of the MTHFR gene in breast cancer cases and controls |
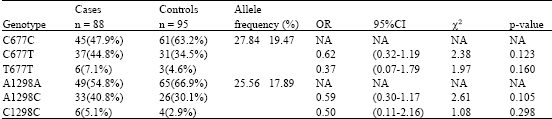 | |
| OR - Odd Ratios, CI - 95% Confidence Intervals, NA – Not Applicable, (%) number of individuals with a given genotype/total number of individuals | |
DISCUSSION
The 5, 10-methylenetetrahydrofolate reductase (MTHFR) gene has been reported to be a genetic modifier of BRCA1 and BRCA2 allele. The potential influence of MTHFR activity on DNA methylation and on the availability of uridylate and thymidylate for DNA synthesis and repair makes MTHFR an attractive candidate for cancer-predisposing gene.
Several independent studies support this hypothesis in which the MTHFR C677T allele may reduce the risk for colon cancer (Slattery et al., 1999) and acute lymphocytic leukemia. Individuals with the C677C genotype had a protective effect for breast cancer, because of the availability of the enzyme in an active form. On the contrary, the MTHFR C677T allele has also been associated with an increased risk for endometrial cancer (Esteller et al., 1997) and cervical intraepithelial neoplasia. Individuals with the C677C genotype had a protective effect for breast cancer, because of the availability of the enzyme in an active form.
The MTHFR C677T and A1298C polymorphisms are likely to play an important role in the susceptibility to breast cancer. In this case control study we have examined the role of MTHFR C677T and A1298C polymorphism in breast cancer patients. A polymorphism in MTHFR gene may also determine a patients risk for chemotherapeutic toxicity. A polymorphism in MTHFR exists at position 677, C to T producing a thermoliable and rapidly degraded enzyme. Folate intake exhibited no modifying effect on the genotype- breast cancer relationship (Merchand et al., 2005).
Ergul et al. (2003) have demonstrated that MTHFR T677T genotype showed a 2.5 fold increase in risk for breast cancer and C1298C genotype showed a 1.9 fold increased risk of breast cancer. Also in compound genotype the 677T homozygous and 1298A homozygous and 677C homozygous and 1298C homozygous showed a 4.472 and 2.301 fold increased risk of breast cancer. However our data shows that MTHFR T677T genotypes and C1298C genotype did not show the risk for breast cancer. The lack of clear relation between MTHFR TT genotypes and breast cancer in this study and the study reported as above by Ergul et al. (2003) and by Marchand et al. (2004) is in sharp contrast to the inverse association that has been reported in colorectal cancer.
There are only few reports on the potential influence of MTHFR activity and breast cancer risk. Gershoni-Baruch et al. (2000) reported that the C677T allele occurred significantly more frequently in Jewish women diagnosed with bilateral breast cancer or combined breast and ovarian cancer (Sharp et al., 2002) did not observe any significant difference in the genotype distributions within a case-control study of unselected breast cancers from a Scottish population Semenza et al. (2003) reported that the MTHFR C677T allele increased the risk for breast cancer in premenopausal rather than postmenopausal women from an American population Campbell et al. (2002) demonstrated that the MTHFR C677T genotype may increase the risk of early-onset breast cancer before the age of 40 years in an English population. In the present study it was found that south Indian women with breast cancer do not show susceptibility to breast cancer with the MTHFR polymorphisms. The importance of the present investigation lies in the fact that the information generated from the genotyped participants had no major difference between case and control studies showing null association between MTHFR gene polymorphism and breast cancer. Even though the current sample size does not allow a generalized prediction to the risk or no-risk factors. TT-homozygous has been reported to have high level of methylenetetrahydrfolate (Merchand et al., 2005).
CONCLUSIONS
The MTHFR C677T and A1298C polymorphisms may be a useful Pharmocogenetic determinant for providing rational evaluation and effective method to chemotherapy, (Kyoung-Jin Sohn et al., 2004) but it has not shown a direct relationship with risks involved in development of breast cancer in our investigation in the south Indian population. This study corroborates previous findings of a null association of the MTHFR (677and1298) genotypes with breast cancer.
REFERENCES
- Cheng, P., C. Schmutte, K.F. Cofer, J.C. Felix, M.C. Yu and L. Dubeau, 1997. Alterations in DNA methylation are early, but not initial, events in ovarian tumorigenesis. Br. J. Cancer, 75: 396-402.
PubMedDirect Link - Deloughery, T.G., A. Evans, A. Sadeghi, J. McWilliams, W.D. Henner, L.M.Jr. Taylor and R.D. Press, 1996. Common mutation in methylenetetrahydrofolate reductase. Correlation with homocysteine metabolism and late-onset vascular disease, 94: 3074-3078.
Direct Link - Ergul, E.A., N.Z. Utkan and Z. Cant, 2003. Polymorphism in the MTHFR gene is associated with Breast cancer. Tumor Biol., 24: 286-290.
CrossRef - Esteller, M., A. Garcia, J.M. Martinez-Palones, J. Xercavins and J. Reventos, 1997. Germ line polymorphisms in cytochrome-P450 1A1 (C4887 CYP1A1) and methylenetetrahydrofolate reductase (MTHFR) genes and endometrial cancer susceptibility. Carcinogenesis, 18: 2307-2311.
Direct Link - Frosst, P., H.J. Blom, R. Milos, P. Goyette and C.A. Sheppard et al., 1995. A candidate genetic risk factor for vascular disease: A common mutation in methylenetetrahydrofolate reductase. Nat. Genet., 10: 111-113.
CrossRefDirect Link - Gershoni-Baruch, R., E. Dagan, D. Israeli, L. Kasinetz, E. Kadouri and E. Friedman, 2000. Association of the C677T polymorphism in the MTHFR gene with breast and/or ovarian cancer risk in Jewish women. Eur. J. Cancer, 36: 2313-2316.
Direct Link - Goyette, P., J.S. Sumner, R. Milos, A.M. Duncan, D.S. Rosenblatt, R.G. Matthews and R. Rozen, 1994. Human methylenetetrahydrofolate reductase: Isolation of cDNA, mapping and mutation identification. Nat. Genet, 7: 195-200.
CrossRef - Jacob, R.A., D.M. Gretz, P.C. Taylor, S.J. James and I.P. Pogribny et al., 1998. Moderate folate depletion increases plasma homocysteine and decreases lymphocyte DNA methylation in postmenopausal women. J. Nutr., 128: 1204-1212.
Direct Link - Jencks, D.A. and R.G. Mathews, 1987. Allosteric inhibition of methylenetetrahydrofolate reductase by adenosylmethionine. Effects of adenosylmethionine and NADPH on the equilibrium between active and inactive forms of the enzyme and on the kinetics of approach to equilibrium. J. Biol. Chem., 262: 2485-2493.
Direct Link - Kim, Y.I., I.P. Pogribny, A.G. Basnakian, J.W. Miller, J. Selhub, S.J. James and J.B. Mason, 1997. Folate deficiency in rats induces DNA strand breaks and hypomethylation within the p53 tumor suppressor gene. Am. J. Clin. Nutr., 65: 46-52.
Direct Link - Kundu, T.K. and M.R. Rao, 1999. CpG islands in chromatin organization and gene expression. J. Biochem., 125: 217-222.
Direct Link - Lengauer, C., K.W. Kinzler and B. Vogelstein, 1997. DNA methylation and genetic instability in colorectal cancer cells. Proced. Natl. Acad. Sci. USA, 94: 2545-2550.
Direct Link - Goyette, P., J.S. Sumner, R. Milos, A.M. Duncan, D.S. Rosenblatt, R.G. Matthews and R. Rozen, 1994. Human methylenetetrahydrofolate reductase: Isolation of cDNA, mapping and mutation identification. Nat. Genet, 7: 195-200.
CrossRef - Matthews, R.G., M.A. Vanoni, J.F. Hainfeld and J. Wall, 1984. Methylenetetrahydrofolate reductase. Evidence for spatially distinct subunit domains obtained by scanning transmission electron microscopy and limited proteolysis. J. Biol. Chem., 259: 11647-11650.
Direct Link - Merchand, L.L., C.A. Haiman, R.L.L. Wilkens, N. Kolonel and B.E. Henderson, 2004. MTHFR polymorphism, diet, HRT and breast cancer risk. The multiethenic cohort study. Cancer Epid. Biomar. Prev., 13: 2071-2077.
Direct Link - Miller, S.A., D.D. Dykes and H.F. Polesky, 1988. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res., 16: 1215-1215.
PubMedDirect Link - Semenza, J.C., R.J. Delfino, A. Ziogas and H. Anton-Culver, 2003. Breast cancer risk and methylenetetrahydrofolate reductase polymorphism. Breast Cancer Res. Treat., 77: 217-223.
PubMedDirect Link - Sharp, L., J. Little, A.C. Schofield, E. Pavlidou and S.C. Cotton et al., 2002. Folate and breast cancer: The role of polymorphisms in ethylenetetrahydrofolate reductase (MTHFR). Cancer Lett, 181: 65-71.
PubMedDirect Link - Slattery, M.L., J.D. Potter, W. Samowitz, D. Schaffer and M. Leppert, 1999. Methylenetetrahydrofolate reductase, diet and risk of colon cancer. Cancer Epid. Biomar. Prev., 8: 513-518.
Direct Link - Sohn, K.J., R. Croxford, Z. Yates, M. Glascock and Y. Kim, 2004. Effect of Methylenetetrahydrofolate reductase C677T polymorphism on chemo sensitivity of colon and breast cancer cells to 5- Fluorouracil and Methotrexate. J. Natl. Cancer Inst., 9: 134-144.
Direct Link - Stampfer, M.J., E. Giovannucci, C. Artigas, D.J. Hunter and C. Fuchs et al., 1997. Methylenetetrahydrofolate reductase polymorphism, dietary interactions and risk of colorectal cancer. Cancer Res., 57: 1098-1102.
PubMedDirect Link - Stern, L.L., J.B. Mason, J. Selhub and S.W. Choi, 2000. Genomic DNA hypomethylation, a characteristic of most cancers, is present in peripheral leukocytes of individuals who are homozygous for the C677T polymorphism in the methylenetetrahydrofolate reductase gene. Cancer Epid. Biomar. Prev., 9: 849-853.
PubMed - Van der Put, N.M., F. Gabreels, E.M. Stevens, J.A. Smeitink and F.J. Trijbels et al., 1998. A second common mutation in the methylenetetrahydrofolate reductase gene: An additional risk factor for neural-tube defects? Am. J. Hum. Genet., 62: 1044-1051.
PubMed - Weisberg, I., P. Tran, B. Christensen, S. Sibani and R. Rozen, 1998. A second genetic polymorphism in methylenetetrahydrofolate reductase (MTHFR) associated with decreased enzyme activity. Mol. Genet. Metab., 64: 169-172.
CrossRef - Wiemels, J.L., R.N. Smith and G.M. Taylor, 2001. Methylenetetrahydrofolate reductase (MTHFR) polymorphisms and risk of molecularly defined subtypes of childhood acute leukemia. Proced. Natl. Acad. Sci., USA, 98: 4004-4009.
Direct Link