
A.R. Ardjmand
Hematology, Oncology and BMT Research Center, Tehran University of Medical Science, Tehran, Iran
K. Alimoghadam
Hematology, Oncology and BMT Research Center, Tehran University of Medical Science, Tehran, Iran
S. Kaviani
Department of Hematology, Faculty of Medicine,
Tarbiat Modares University, Tehran, Iran
A. Ghavamzadeh
Hematology, Oncology and BMT Research Center, Tehran University of Medical Science, Tehran, Iran
M. Djahani
Hematology, Oncology and BMT Research Center, Tehran University of Medical Science, Tehran, Iran
L. Moezzi
Hematology, Oncology and BMT Research Center, Tehran University of Medical Science, Tehran, Iran
International Journal of Cancer Research
Year: 2006 | Volume: 2 | Issue: 2 | Page No.: 124-130







ABSTRACT
Arsenic trioxide (ATO) has been reported to induce apoptosis in Leukemic cells of Acute Promyelocytic Leukemia (APL) patients through different pathways. However, the exact mechanism of ATO-induced apoptosis is not yet clear. Co stimulation of death receptors CD30 and tumor necrosis factor receptor type one (TNFR-I) is one of the postulate mechanisms which in the present study we aimed to evaluate their involvement in fresh Promyelocytic cells separated from bone marrow of APL patients. Immunomagnetic separated cells were treated up to 48 h at clinically tolerable concentration of ATO (0.5-2.0 μmol L-1) and expression of TNFR-I and CD30 were evaluated within the apoptotic and live populations using a sensitive triple color flow cytometric method for measuring apoptosis in combination with dual color immunofluorescence. Present results suggest that the expression of TNFR-I and CD30 might not be related to ATO-induced apoptotic cell death.
PDF Abstract XML References
How to cite this article
A.R. Ardjmand, K. Alimoghadam, S. Kaviani, A. Ghavamzadeh, M. Djahani and L. Moezzi, 2006. Evaluation of Apoptosis Induced by Arsenic Trioxide Through TNFR-I and CD30 Pathways in Acute Promyelocytic Leukemia Patient with t (15;17) Translocation. International Journal of Cancer Research, 2: 124-130.
DOI: 10.3923/ijcr.2006.124.130
URL: https://scialert.net/abstract/?doi=ijcr.2006.124.130
DOI: 10.3923/ijcr.2006.124.130
URL: https://scialert.net/abstract/?doi=ijcr.2006.124.130
INTRODUCTION
Acute Promyelocytic Leukemia (APL), which comprises about 10-15% of cases of acute leukemia in adults, characterized with prominent malignant Promyelocyte in bone marrow. The pathogenesis of APL involves a reciprocal chromosomal translocation associated with Promyelocytic Leukemia (PML) and Retinoic Acid Receptor α (RAR α) genes on chromosomes 15 and 17, respectively (Warrell et al., 1993; Grignani et al., 1993).
Arsenic trioxide (ATO) as an effective treatment with little adverse effects (Antman, 2001; Guo-Qiang et al., 1997) apparently disturb numerous intracellular signal transduction pathways and causes many alterations in cellular functions among which the most prominant mechanisms are induction of differentiation and apoptosis with low and high does of ATO, respectively (Miller, 2002; Miller et al., 2002).
Induction of apoptosis occurs through multiple pathways, some principal signals pathways of apoptosis have been identified. One pathway involves activation of death receptors such as CD95, Tumor Necrosis Factor (TNF) receptor and TNF-related Apoptosis-Inducing Ligand (TRAIL), followed by recruitment of the Fas-associated protein with death domain and activation of caspase 8. The other pathway is triggered by disruption of the mitochondrial membrane by many insults and results in the loss of mitochondrial transmemberane potential (Δψm) and release of mitochondrial proteins such as cytochrome c and apoptosis-inducing factor into cytosol followed by activation of caspase 9. Caspase 8 and 9 activated by either pathway transduce which signals activate caspase 3 and subsequently cleave poly (ADP-ribose) polymerase (PARP).
Since activation of Caspase 8 which induces mitochondrial Δψm collapse (Cai et al., 2000; Woo et al., 2002) following arsenic treatment (Li et al., 1998; Luo et al., 1998) is absolutely required for TNF mediated apoptosis (Nicholas et al., 2003). The obsertive of this Study was to check the TNF receptor type I (TNFR-I) involvement in ATO apoptosis induction. But due to divers biological effect of TNF, evaluation of its co-stimulatory family member, CD30, also seemed to be necessary (Duckett and Thompson, 1997).
Here to study the expression of TNFR-I and CD30 within the apoptotic and live leukemic cells of de novo APL patients, we conduct a tricolor flow cytometric method for measuring apoptosis in combination with dual color immunofluorescence.
MATERIALS AND METHODS
Isolation of Promyelocytes and Arsenic Treatment
Bone Marrow (BM) samples were collected from 5 de novo APL patients based on FAB classification and cytogenetic studies for t (15;17) translocation. MNC's were isolated from heparinized BM samples after centrifugation at 600xg for 20 min over Ficoll-Hypaque (1.077). (Pharmacia, Uppsala, Sweden).
Freshly obtained MNC's were then further purified for promyelocyte as previously described (Flo et al., 1991) and according to Dynal bead negative selection system's instruction. Briefly a total number of 4x106 of obtained MNC's were incubated with anti human HLA-DR and Anti-human CD3 antibodies (Serotec, London, UK) for 30 min on ice. Then cells washed for three times in RPMI 1% FBS (Gibco,Gaithersburg, MD) and incubated with Daynabeads pan mouse IgG for the next 30 min on ice and continuous gentle shaking. Finally by pipetting off the fluid from bottom of the tube, HLA-DR negative Promyelocytes were obtained and cultured at 105 cells/well in a 24-well culture plate in the presence of 0.5, 1.5 and 2 μmol L-1 Arsenic trioxide (Sigma, St. Louis, MO) in RPMI 1640 with 10% Fetal Calf Serum (FCS) in a humidified, 5% Co2 atmosphere at 37°C.
TNFR-I (CD120a) and CD30 Expression Within the Context of Apoptotic and Live Cells
To measure apoptosis in combination with cell surface immunophenotyping we used DNA dye 7-AAD (Schmid et al., 1994).
After 36 and 48 h of treatment, cultured Promyelocyte were harvested and washed twice in cold PBS 1% FCS and 105 of cells were incubated for 10 min in 2% mouse serum before staining with anti-human CD120a-FITC and anti-human CD30-PE MoAb (Serotec, London, U.K) or FITC and PE isotype control (Dako and Denmark), respectively. For discrimination of apoptosis from secondary necrosis, cells were then incubated with 7-AAD (20 μg L-1) in PBS for 20 min at 4°C, washed in PBS and immediately submitted for flowcytometric analysis. A minimum of 10,000 events was acquired on each sample. For the analysis of apoptotic cells, gates were set on cells staining dimly with 7-AAD; and cells staining brightly with 7-AAD were discriminated as dead cells and was not considered within the analysis.
Statistical Analysis
Control and ATO treated cells were compared for differences in expression of CD120a and CD30 in apoptotic and live cells after 36 and 48 h of culture.
Statistical comparisons were performed by using the Paired t-test. Significance was considered at p<0.05.
RESULTS
Expression of CD120a and CD30
To study the mechanism responsible for arsenic induced cell death, changes in expression of CD120a and CD30 on Promyelocytes of APL patients were examined after 36 and 48 h of treatment. In this regard, firstly live, apoptotic and dead cells were discriminated by virtue of their size changes and 7-AAD DNA dye uptake (Fig. 1).
Principally cells undergoing apoptosis, activate a series of molecular and biochemical events which lead to their total physical disintegration. One of the early events is cell dehydration, cytoplasm and nuclear chromatin condensation. Cells size and shape are characterized by light scattered at both forward and right angle of the laser beam in flowcytometry systems.
Although a transient increase in right angle scatter can be seen during apoptosis in some cell systems, in later stages of apoptosis, however, the intensity of light scatter at both forward and right angle directions is decreased (Ormerod et al., 1995).
Cell membrane permeability changes is another event that features distinguishing dead from live cells and can be detected by the ability of intact membrane in excluding charged cationic dyes such as trypan blue, Propidium Iodide (PI), ethidium bromide or 7-aminoactinomycine D (7-AAD).
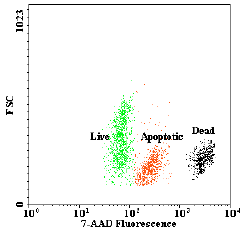 | |
| Fig. 1: | Discrimination of live, apoptotic and dead cells by using Forward light scatter (FSC) and 7-AAD DNA dye uptake. Despite necrosis, cells under going apoptosis show a cellular condensation and hence a decreament in FSC signal, on the other hand due to membrane permeability disruption and inability of cells in excluding 7-AAD, cells uptake more DNA dye and show brighter 7-AAD fluorescence. Accordingly green, red and black populations are live, apoptotic and dead cells, respectively |
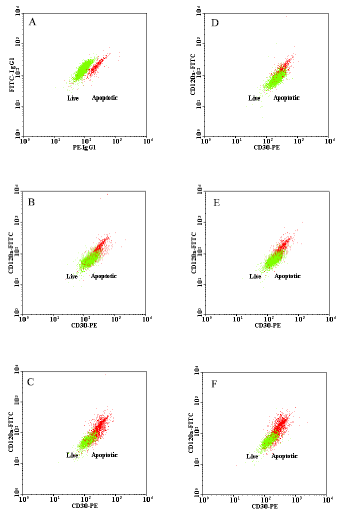 | |
| Fig. 2: | Expressions of TNFR-1, CD120a and CD30 on apoptotic (Red) and live (Green) cells of non-arsenic and arssenic treaed cells: A: nonspecific binding of apoptotic and live cells to nonspecific Abs lgGl-FIITC and lgGl-PE represents higher affinity of apoptotic cells to IgGl-PE. B: Non-arsenictreated promyelocytes as control. C-F: 0.5, 1.5, 2.0 μ Mol aresnic treated cells after 48 h |
During apoptosis the plasma membrane transport function becomes transiently defective prior to total loss of the ability to exclude these charged fluorochromes. At this stage of apoptosis, the rate of uptake of these nuclear dyes increases comparing to live cells. So the more cells progress in apoptosis, the more DNA dye they uptake.
As illustrated by color gating, comparing to live cells (green), apoptotic populations (red) are significantly (p<0.05) over expressing CD120a molecule albeit irrelevant to arsenic. But in the case of CD30, since its isotype control also shows a higher affinity to apoptotic cells we believe that there is no over expression of this molecule (Fig. 2 and 3).
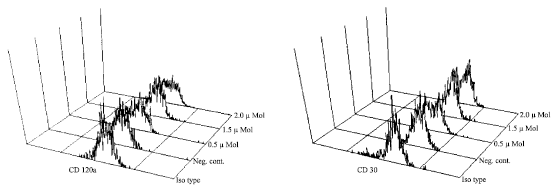 | |
| Fig. 3: | Expression of TNFR-I (CD120a) and CD30 on apoptotic cells of different doses of arsenic after 48 h treatment. Events were normalized and no over expression were detected on both receptors |
DISCUSSION
One of the important mechanisms of apoptosis is regulated by death receptor associated death domain signaling intermediates, which contain receptor activation induced binding death domain proteins, such as Tumor Necrosis Factor (TNF) receptor-associated death domain protein (TRADD) and Fas-Associated Death Domain Protein (FADD) and further activation of caspase cascade that initiate apoptotic death (Wallach et al., 1999). A major apoptotic signal in MNCs is mediated by TNF-α and its receptor. TNF-α which was shown to be produced as a proinflamatory cytokine from the inorganic arsenicals treated peritoneal macrophage (Sakurai et al., 1998) is a pleiotropic cytokine that mediates a wide variety of biological responses, including the effects on the growth and differentiation of normal and malignant hematopoetic cells and also the induction of tumor cell death. Two types of TNF receptors have been identified for the TNF, namely TNFR-I and TNFR-II which may mediate the activation of independent downstream signaling pathway (Tartaglia et al., 1991) TNFR-II functions primarily to bind ligand rapidly, passing soluble TNF trimers to TNFR-I (Pinckard, 1997). Signaling by TNF receptor causes the association of an adaptor protein TRADD with the intercellular death domain of the TNFR-I molecule (Ashkenzai, 1998) TRADD mediates the subsequent recruitment of an adapter protein FADD to form a death inducing signaling complex, which initiates apoptosis through activation of caspase cascades. Signaling through TNFR-I is, however more complex than CD95 indeed, in addition to recruiting adaptor molecules that bind and activate caspases, TNFR-I recruits proteins that engage various signal transduction pathways, some of which either abrogates or potencies the apoptotic response for example, in some cells exposed to TNF, NF-κB is induced through the activation of TNF Receptor Associated Factor II (TRAF II) and the cells survives (NF-κB protects some cells from TNF induced program cell death). However, ligation of CD30 induces TRAF II degradation, which then disconnects the TNF receptor and NF-κB activation. Consequently, when cells are simultaneously stimulated through the TNFR-I and CD30 molecules, the cells will under go apoptosis.
Expression of CD30 in several types of malignancies including Hodgkin's disease, anaplastic large-cell lymphoma, immunoblastic lymphoma, multiple myeloma, adult T-cell leukemia lymphoma and also its TNF family member CD30L in a variety of hematopoietic cells (Younes and Kadin, 2003) prompt us to evaluate the above co-stimulatory pathway in arsenic apoptosis induction.
In the present study CD30 expression was not evidenced whiles significant (p<0.05) over expression of TNFR-I on apoptotic populations of arsenic treated and non-treated Promyelocytes after 36 and 48 h indicated that arsenic does not affect TNFR-I expression either.
Since the PML/RAR-α fusion protein of leukemic promyelocytes which is responsible for resistance induction to TNF-α mediated apoptosis (Ugo et al., 1998) was shown to be degraded by arsenic (Miller et al., 2002) and that TNF-α dose not necessarily induce or enhance apoptosis through its ligand binding pathway (Schmelz et al., 2004), arsenic hypothetically may restore the abrogated sensitivity of these cells to TNF-α rather than induction of their over expression.
REFERENCES
- Ashkenazi, A. and V.M. Dixit, 1998. Death receptors: Signaling and modulation. Science, 281: 1305-1308.
CrossRefPubMedDirect Link - Cai, X.,Y.L. Shen, Q. Zhu, P.M. Jia and Y. Yu et al., 2000. Arsenic trioxide-induced apoptosis and differentiation are associated, respectively with mitochondrial transmembrane potential collapse and retinoic acid signaling pathways in acute Leukemia, 14: 262-270.
Direct Link - Flo, R.W., A. Naess, F. Lund-Johansen, B.O. Maehle, H. Sjursen, V. Lehmann and C.O. Solberg, 1991. Negative selection of human monocytes using magnetic particles covered by anti-lymphocyte antibodies. J. Immunol. Methods, 137: 89-94.
PubMedDirect Link - Grignani, F., P.F. Ferrucci, U. Testa, G. Talamo and M. Fagioli et al., 1993. The acute promyelocytic leukemia-specific PML-RAR alpha fusion protein inhibits differentiation and promotes survival of myeloid precursor cells. Cell, 74: 423-431.
PubMed - Chen, G.Q., X.G. Shi, W. Tang, S.M. Xiong and J. Zhu et al., 1997. Use of Arsenic Trioxide (As2O3) in the Treatment of Acute Promyelocytic Leukemia (APL): I. As2O3 exerts dose-dependent dual effects on APL cells. Blood, 89: 3345-3353.
Direct Link - Li, H., H. Zhu, C.L. Xu and J. Yuan, 1998. Cleavage of BID by caspase 8 mediates the mitochondrial damage in the Fas pathway of apoptosis. Cell, 94: 491-501.
CrossRefPubMedDirect Link - Luo, X., I. Budihardjo, H. Zou, C. Slaughter and X. Wang, 1998. Bid, a Bcl2 interacting protein, medi-ates cytochrom C release from mito-chondria in response to activation of cell surface death receptors. Cell, 94: 481-490.
PubMedDirect Link - Miller, W.H., 2002. Molecular targets of arsenic trioxide in malignant cells. Oncologist, 7: 14-19.
Direct Link - Miller, W.H., H.M. Schipper, J.S. Lee, J. Singer and S. Waxman, 2002. Mechanisms of action of arsenic trioxide. Cancer Res., 62: 3893-3903.
PubMedDirect Link - Nicholas, H., M. Hughes, M. Marion and M.C. Gerald, 2003. Fas-associated death domain protein and Caspase-8 are not recruited to the Tumor Necrosis Factor Receptor 1 signaling complex during tumor necrosis factor-induced apoptosis. J. Biol. Chem., 278: 1125534-1125541.
PubMedDirect Link - Ormerod, M.G., F. Paul, M. Cheetham and X.M. Sun, 1995. Discrimination of apoptotic thymocytes by forward light scatter. Cytometry, 21: 300-304.
Direct Link - Sakurai, T., T. Kaise and C. Matsubara, 1998. Inorganic and methylated arsenic compounds induce cell death in murine macrophages via different mechanisms. Chem. Res. Toxicol., 11: 273-283.
CrossRef - Schmid, I., C. H. Uittenbogaart, B. Keld and J.V. Giorgi, 1994. A rapid method for measuring apoptosis and dual-color immunofluorescence by single laser flowcytometry. J. Immunol. Methods, 170: 145-157.
PubMedDirect Link - Schmelz, K., T. Wieder, I. Tamm, A. Muller and F. Essmann et al., 2004. Tumor necrosis factor alpha sensitizes malignant cells to chemoteraputic drugs via the mitochondrial apoptosis pathway independently of caspase-8 and NF-κB. Oncogene., 23: 6743-6759.
Direct Link - Tartaglia, L.A., R.F. Weber, I.S. Figari, C. Reynolds, M.A. Palladino and D.V. Goeddel, 1991. The two different receptor for tumor necrosis factor mediate distinct cellular responses. Proc. Natl. Acad. Sci. USA, 88: 9292-9296.
Direct Link - Wallach, D., E.E. Varfolomeev, Y.V. Goltsev, A.V. Kovalenko, M.P. Boldin, 1999. Tumor necrosis factor receptor and Fas signaling mechanisms. Ann. Rev. Immunol., 17: 331-367.
PubMedDirect Link - Warrell, Jr. R.P., H. de The, Z.Y. Wang and L. Degos, 1993. Acute promyelocytic leukemia. N. Engl. J. Med., 329: 177-189.
PubMed - Woo, S.H., I.C. Park, M.J. Park, H.C. Lee and S.J. Lee et al., 2002. Arsenic trioxide induces apoptosis through a reactive oxygen species-dependent pathway and loss of mito-chondrial membrane potential in Hela cells. Int. J. Oncol., 21: 57-63.
Direct Link - Younes, A. and M.E. Kadin, 2003. Emergimg application of the tumor necrosis factor family of ligands and receptors in cancer therapy. J. Clin. Oncol., 21: 3526-3534.
Direct Link