
Khalil I. Al-Mughrabi
Department of Agriculture and Aquaculture, Wicklow, New Brunswick, E7L 3S4, Canada
International Journal of Biological Chemistry
Year: 2008 | Volume: 2 | Issue: 1 | Page No.: 14-25







ABSTRACT
Trials were conducted at the Potato Development Centre, Wicklow, New Brunswick, Canada to study the effect of Salicylic Acid (SA) in suppressing black scurf (Rhizoctonia solani Kuhn, AG-3) disease in potatoes (cv. Atlantic) under screenhouse conditions. The trials were designed as a completely randomized block and comprised of eight replicated treatments: Untreated, uninoculated control (CTH); untreated control inoculated with R. solani (CTD); healthy seed treated with SA (STH); seeds inoculated with R. solani and treated with SA (STD); healthy seeds with SA applied foliarly (FAH); seeds inoculated with R. solani and SA applied foliarly (FAD); healthy seeds with SA applied as soil drench (SDH); seeds inoculated with R. solani and SA applied as soil drench (SDD). Seedling emergence, plant canopy, disease severity of black scurf and total tuber weights were recorded. Plant canopy was significantly higher in plants inoculated with R. solani and treated with SA compared to uninoculated treatments. All SA treatments significantly reduced black scurf disease severity in stems compared to the untreated, inoculated controls. Compared to the CTD, black scurf disease severity in stems was reduced by 89.6 and 88.8% when FAH and SDH treatments, respectively, were used. Treatments inoculated with R. solani and treated with SA significantly increased potato tuber weights compared to the uninoculated controls. Present findings indicate that SA has the potential to be used as an alternative tool in managing black scurf disease in potatoes.
PDF Abstract XML References Citation
How to cite this article
Khalil I. Al-Mughrabi, 2008. Salicylic Acid Induces Resistance in Potatoes Against Rhizoctonia solani, the Cause of Black Scurf and Stem Canker. International Journal of Biological Chemistry, 2: 14-25.
URL: https://scialert.net/abstract/?doi=ijbc.2008.14.25
URL: https://scialert.net/abstract/?doi=ijbc.2008.14.25
INTRODUCTION
Black scurf (Rhizoctonia solani Kuhn, AG-3) is a common disease of potatoes which is capable of causing significant reductions in tuber quality and yield worldwide (Anderson, 1982; Little et al., 1988; Carling et al., 1989; Powelson et al., 1993; Jeger et al., 1996; Tsror et al., 1996). The disease is more prevalent under cool and moist environmental conditions (Bandy et al., 1988; Carling et al., 1989; Banville, 1989) and may not occur in drier and warmer conditions (Davis, 1978; Weinhold et al., 1982). Typical disease symptoms include black sclerotia formation on tubers and the presence of sunken necrotic lesions on roots, stolons and subterranean portions of the main stem (Carling and Leiner, 1986). The severity of black dot is not always associated with yield reduction. However, the formation of tuber-borne sclerotia downgrades quality of tubers, resulting in malformed tubers and an alteration in target size and tuber numbers (Anderson, 1982; Carling et al., 1989; Jeger et al., 1996). Infection of potato plants with R. solani can be caused by soil borne inoculum, tuber-borne inoculum, or via infected seed tubers (Banville, 1978; Platt, 1989; Powelson et al., 1993; Platt et al., 1993). Studies have shown that both tuber-borne and soil-borne inocula are important in the development of black scurf disease in potatoes (Frank and Leach, 1980; Tsror and Peretz-Alon, 2005). It is also suggested that the control of tuber-borne inoculum is essential for the integrated management of R. solani in potatoes (Frank and Leach, 1980). Losses caused by this pathogen can be minimized by using disease free seed tubers, planting seed near the soil surface and following a rotation scheme with two or more years between potato crops. In some cases, an increase in black scurf of potato was observed following crop rotation (Celetti et al., 1990; Errampalli et al., 1999). In addition, fungicides effective against soil-borne fungal pathogens of potato, including R. solani, are very limited (Powelson et al., 1993; Wicks et al., 1995; Johnston, 1995).
The phenomenon by which a plant exhibits an increased level of resistance to infection by a pathogen after appropriate stimulation is called induced disease resistance. The two forms of induced resistance that are clearly defined are Systemic Acquired Resistance (SAR) and Induced Systemic Resistance (ISR). These two forms of resistance can be differentiated from each other on the basis of the nature of elicitor and the regulatory pathways involved (Knoester et al., 1999; Maleck et al., 2000; Schenk et al., 2000; van Wees et al., 2000; Yan et al., 2002). In SAR, a plant`s own defense mechanisms are triggered by prior treatment with either a biological agent or chemical agent. Salicylic acid is an endogenous signaling molecule required for the induction of SAR (Klessig and Malamy, 1994). SAR is associated with induction of a group of Pathogenisis-Related (PR) genes and their corresponding proteins (Pieterse et al., 1996; Delaney, 2000). In ISR, the resistance is activated by biotic and abiotic inducing agents and is dependent on the host plant`s physical or chemical barriers (Kloepper et al., 1992; Kessmann et al., 1994). Contrary to SAR, ISR does not involve accumulation of PR proteins but relies on pathways regulated by jasmonate and ethylene (Pieterse et al., 1998; Knoester et al., 1999; Yan et al., 2002). ISR is brought about by certain group of plant growth promoting rhizobacteria, which includes several species of Pseudomonas, without causing any damage to the plant`s root system (van Loon et al., 1998). It is believed that different antimicrobial proteins are induced in the two pathways, but the distinction of terminology is somewhat confused due to the possibility of crosstalk between SAR and ISR (Dong, 2001). Plants successfully resisting a pathogen during SAR have the capability to become highly resistant to subsequent infection not only by the original pathogen but also by other pathogens (Coquoz et al., 1995; Lawton et al., 1996). Acetyl Salicylic Acid (ASA; the active ingredient in aspirin) is a product known to provoke the SAR in plants, therefore raising their natural defense against potential diseases (Gaffney et al., 1993; Lawton et al., 1995; Dempsey et al., 1999; Verberne et al., 2000; Rairdan and Delaney, 2002). SA applied as seed treatment, spray and soil drench, reduced the number of lesions, lesion size and the percentage of necrotic leaf area following infection by Phytophthora palmivora in cacao (Okey and Sreenivan, 1996). However, SA has not been tested against potato diseases including black scurf. The objective of the present study was to assess the effect of SA in reducing the severity of black scurf under screenhouse conditions.
MATERIALS AND METHODS
Screenhouse Experiments
Screenhouse experiments were conducted at the Potato Development Centre, Wicklow, New Brunswick, Canada. Disease-free seed potato tubers (cv. Atlantic) were washed, air dried and used in the experiments. A solution of salicylic acid (SA) [HOC6H4COOH, crystals, FW 138.12, EM Science, USA] was prepared by mixing 400 mg of SA in 600 μL of tween 80 and 200 mL of sterile distilled water (Bokshi et al., 2003). This solution was diluted with distilled water to obtain a final volume of 6 L.
Experimental Set up
The experiment was designed as a one way completely randomized block and consisted of 8 treatments which were replicated four times. Each replicate consisted of 5 potato plants (cv. Atlantic). The eight treatment were: 1) untreated, uninoculated control (CTH); 2) untreated control inoculated with R. solani (CTD); 3) healthy seed treated with SA (STH); 4) seed inoculated with R. solani and treated with SA (STD); 5) healthy seed with SA applied as foliar spray (FAH); 6) seed inoculated with R. solani with SA applied as foliar spray (FAD); 7) healthy seed with SA applied as soil drench (SDH); 8) seed inoculated with R. solani with SA applied as soil drench (SDD). The experiment (8 treatments x 4 replicates x 5 plants) was repeated twice. Pots were filled up to half with professional sphagnum peat-based growing medium (Premier Pro-Mix®, Premier Horticulture Ltée, Rivière-du-Loup, Québec) and used for the trial. All purpose plant food fertilizer was applied once every two weeks (Greenleaf Shur-Gro®, Greenleaf Products Inc., Burnaby, BC, N-P-K 20-20-20 applied at the rate of 5 mL in 4 L of water).
Rhizoctonia solani inoculum was prepared by culturing a naturally infected piece of tuber on Potato Dextrose Agar (PDA). The culture obtained was purified and multiplied on fresh PDA plates and allowed to grow for seven days at room temperature. Organic rye seeds were soaked in water for 24 h and then autoclaved at 121°C for 15 min. Each bag of rye seed (1 kg) received six plates of R. solani culture cut up into 1 inch cubes under aseptic conditions. The bags were then stored at room temperature for 60 days and were shaken weekly. The autoclaved, organically-grown rye seed inoculated with R. solani served as inoculum for the screenhouse studies. For treatments where inoculation was required, 50 g of R. solani infected rye seed was added on and around each potato seed piece at planting.
Seedling emergence was recorded once every 2 days starting 21 days after planting and until all plants emerged. A foliar spray and soil drench was carried out after full emergence, 5 weeks after planting. The solution used in foliar spray and soil drench was the same as the one used in seed treatment. It consisted of a mixture of 448 mg SA, 112 μL of Tween 80 and 1.12 L of water. The tween was used in order to make SA more soluble in water. The concentration needed was 0.01% of the final volume of the solution. The final concentration of SA was 400 mg L-1.
The amount of spray applied was based on the standard pesticide application rate of 101 L acre-1. This amounted for 14 mL of solution per plant. The soil drench was applied using a disposable pipette. However, for the foliar application, a spray bottle was calibrated in order to determine the number of sprays needed to deliver 14 mL. The tubers were harvested and were later assessed for disease severity on a scale of 0-100% (Cruickshank et al., 1982; Dorrance and Inglis, 1997). Data were analyzed using CoStat (CoHort Software, Monterey, CA, USA) and the means were separated using LSD test at p = 0.05.
RESULTS
Seedling emergence was significantly higher in the uninoculated treatments compared to the inoculated ones in the initial stages. As the days progressed, the emergence in R. solani inoculated treatments did not differ significantly compared to uninoculated treatments (Fig. 1). In the initial stages significant differences in emergence were observed between FAH (25%), SDH (25%) and CTH (25%) over FAD (0%), SDD (0%), CTD (0%) and STD (0%). Over time, the significant differences in emergence were observed between all uninoculated treatments and CTD and STD (Fig. 1).
Plant canopy was significantly higher in plants inoculated with R. solani and STD (12.19 L plant-1), FAD (13.48 L plant-1) and SDD (12.31 L plant-1) treatments over the uninoculated treatments (Fig. 2). The highest canopy was for FAD (13.48 L plant-1) followed by SDD (12.31 L plant-1), STD (12.19 L plant-1) and CTD (9.37 L plant-1). Canopies of all SA treated plants were significantly higher than that for CTD.
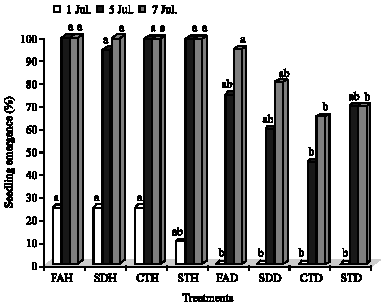 | |
| Fig. 1: | Seedling emergence of potato tubers inoculated with Rhizoctonia solani and treated with salicylic acid. CTH = uninoculated untreated control; STH = healthy seed treated with salicylic acid; FAH = healthy seeds with salicylic acid applied as a foliar spray; SDH = healthy seeds with salicylic acid applied as a soil drench; CTD = inoculated untreated control; STD = seeds infected with R. solani and treated with salicylic acid; FAD = seeds infected with R. solani with salicylic acid applied as a foliar spray and SDD = seeds infected with R. solani with salicylic acid applied as a soil drench. Bars followed by the same letters are not significantly different from each other at p = 0.05 |
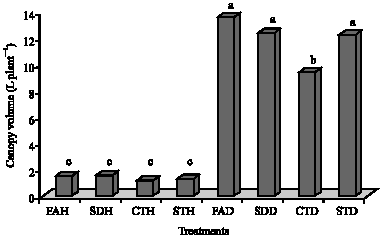 | |
| Fig. 2: | Plant canopy measurements of potato tubers inoculated with Rhizoctonia solani and treated with salicylic acid. CTH = uninoculated untreated control; STH = healthy seed treated with salicylic acid; FAH = healthy seeds with salicylic acid applied as a foliar spray; SDH = healthy seeds with salicylic acid applied as a soil drench; CTD = inoculated untreated control; STD = seeds infected with R. solani and treated with salicylic acid; FAD = seeds infected with R. solani with salicylic acid applied as a foliar spray and SDD = seeds infected with R. solani with salicylic acid applied as a soil drench. Bars followed by the same letters are not significantly different from each other at p = 0.05 |
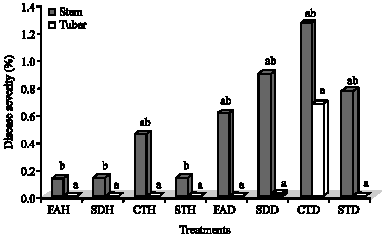 | |
| Fig. 3: | Disease severity of black scurf in potato tubers inoculated with Rhizoctonia solani and treated with salicylic acid. CTH = uninoculated untreated control; STH = healthy seed treated with salicylic acid; FAH = healthy seeds with salicylic acid applied as a foliar spray; SDH = healthy seeds with salicylic acid applied as a soil drench; CTD = inoculated untreated control; STD = seeds infected with R. solani and treated with salicylic acid; FAD = seeds infected with R. solani with salicylic acid applied as a foliar spray; and SDD = seeds infected with R. solani with salicylic acid applied as a soil drench. Bars followed by the same letters are not significantly different from each other at p = 0.05 |
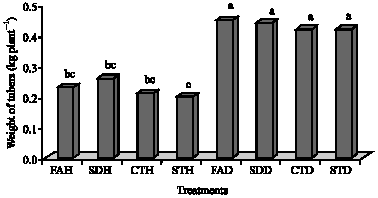 | |
| Fig. 4: | Total weight of tubers inoculated with Rhizoctonia solani and treated with salicylic acid. CTH = uninoculated untreated control; STH = healthy seed treated with salicylic acid; FAH = healthy seeds with salicylic acid applied as a foliar spray; SDH = healthy seeds with salicylic acid applied as a soil drench; CTD = inoculated untreated control; STD = seeds infected with R. solani and treated with salicylic acid; FAD = seeds infected with R. solani with salicylic acid applied as a foliar spray and SDD = seeds infected with R. solani with salicylic acid applied as a soil drench. Bars followed by the same letters are not significantly different from each other at p = 0.05 |
Compared to the inoculated, untreated control (CTD), black scurf disease severity was reduced by 88.8 and 89.6% in stems of plants receiving SDH and FAH treatments, respectively. The highest disease severity of R. solani in stems (1.25%) and tubers (0.67%) was observed in the CTD treatment (Fig. 3). CTD (1.25%) differed significantly from STH (0.14%), SDH (0.14%) and FAH (0.13%) with regard to disease severity in the stems. All treatments tested did not differ significantly among themselves with respect to disease severity in tubers (Fig. 3).
All treatments that were inoculated with R. solani and treated with SA yielded significantly higher tuber weights than the remaining treatments (Fig. 4). The highest tuber weight (yield) was recorded in FAD (0.45 kg plant-1) followed by SDD (0.44 kg plant-1), CTD (0.42 kg plant-1) and STD (0.42 kg plant-1) (Fig. 4).
DISCUSSION
Black scurf of potatoes caused by Rhizoctonia solani is an economically important disease resulting in poor quality tubers and reduced yields (Anderson, 1982; Little et al., 1988; Powelson et al., 1993; Tsror et al., 1996). Fungicides that are effective in managing soil-borne fungal pathogens including R. solani are scarce (Powelson et al., 1993; Johnston, 1995; Wicks et al., 1995). In addition, crop rotations followed to reduce incidence of black scurf have not been very successful (Celetti et al., 1990; Errampalli et al., 1999). There is a need to look for alternative methods to control black scurf. Use of SA and other derivatives have been shown to be involved in the suppression of several pathogens (Okey and Sreenivan, 1996; Romero et al., 2001; Pérez et al., 2003). In the present study, the effect of SA in reducing the severity of black scurf was studied under screenhouse conditions.
The treatments which received SA in the form of seed treatment, foliar spray or soil drench improved the germination of seed tubers and plant canopy compared to the control. In addition, all SA treatments increased plant canopy in a significant manner compared to the uninoculated treatments. The SA treatments reduced the severity of R. solani considerably in both stems and tubers, ultimately improving tuber yield. In another study, lesion number, lesion area and percentage of necrotic leaf area of two cacao clones infected with Phytophthora palmivora were reduced when SA was applied as seed treatment, spray and soil drench (Okey and Sreenivan, 1996). Foliar application of synthetic analogs of SA such as 2,6-dichloroisonicotinic acid (INA) also reduced fire blight symptoms by 45% when compared to uninoculated controls (Kessmann et al., 1994). In chickpea, exogenously supplied SA stimulated systemic resistance against Fusarium wilt and reduced the disease severity by 23 and 43% in plants which were treated with 40 and 80 μg mL-1 of SA through root application (Saikia et al., 2003). Pre-harvest treatment of sweet cherry with 2 mM SA significantly reduced lesion diameters caused by Monilinia fructicola compared to control post-harvest treatments (Yao and Tian, 2005). Similarly, treating sweet cherry with SA at 0.5 mM for 10 min reduced the incidence of decay and lesion size caused by Penicillium expansum (Chan and Tian, 2006).
In plants, exposure to pathogens triggers the activation of resistance mechanisms which prevents infection, aids in recovery from disease and prevents further infection. This phenomenon by which a plant`s own defense mechanisms are induced due to prior treatment with either biological or chemical agent is known as Systemic Acquired Resistance (SAR) (Ryals et al., 1996; Sticher et al., 1997). SAR-protected plants when infected with a pathogen exhibit morphological and biochemical changes such as faster lignification response which corresponds to an increase in peroxidase activity (Ajilan and Potter, 1992) increased glucose and fructose concentrations in systemic tissue (Chandra and Bhatt, 1998); accumulation of fungitoxic β-ionone derivatives (Wyatt and Kuc, 1992); induction of lipoxygenase (Staub et al., 1992), antimicrobial fatty acid derivatives (Namai et al., 1993), phenylalanine ammonialysase, phytoalexins (Elliston et al., 1977) and hydroxyproline-rich glycoprotein (Raggi, 1998). When activated, SAR can prevent infection from a wide range of pathogens (Bowling et al., 1994; Coquoz et al., 1995; Lawton et al., 1996). It is associated with the induction of a suite of pathogenesis-related (PR) genes including acidic and basic β-1-3-glucanases, chitinases and a wide array of other genes of unknown function (Linthorst, 1991; Ward et al., 1991). The pathways in induced resistance mechanisms are regulated by important signal molecules such as SA, jasmonate and ethylene, all of which bring about substantial alterations in gene expression and are involved in complex crosstalk (Maleck et al., 2000; Glazebrook et al., 2003; Spoel et al., 2003). SA is a naturally occurring, endogenously produced compound which is required for the induction of SAR (Gaffney et al., 1993; Rairdan and Delaney, 2002). Treatment of plants with SA or its synthetic analogs 2,6-dichloroisonicotinic acid (INA) or benzo (1,2,3)thiadiazole-7-carbothioic acid S-methyl ester (BTH) results in induction of PR gene expression and resistance.
The importance of SA in SAR was illustrated by removing SA through the ectopic expression of salicylate hydroxylase (NahG) which ultimately blocked the onset of SAR (Gaffney et al., 1993). The product of salicylate hydroxylase gene catalyzes conversion of SA into catechol which is an inactive compound. In NahG plants, the production of SA and induction of SAR was not observed and the NahG plants are hyper-susceptible to a wide range of pathogens (Delaney et al., 1994; Navarre and Mayo, 2004). Similarly, increasing SA concentrations through endogenous synthesis or exogenous application played a role in the induction of SAR (White, 1979; Métraux et al., 1990; Rasmussen et al., 1991; Malamy et al., 1992; Enyedi et al., 1992; Verberne et al., 2000). SA-mediated signaling is well characterized in Arabidopsis, tobacco and cucumber compared to potato and rice which produce high levels of basal SA (Hammerschmidt and Kuc, 1982; Raskin et al., 1990; Coquoz et al., 1995; Dempsey et al., 1999; Navarre and Mayo, 2004).
The precise mechanism by which induction of SAR leads to resistance is not completely understood while the several PR genes expressed during resistance development were antagonistic to pathogens (Alexander et al., 1993; Cao et al., 1994; Sticher et al., 1997). So far, no single PR gene has been shown to be essential for SAR indicating the necessity of coordinated expression of these genes for production of resistance (Rairdan et al., 2001). In plants, SA levels increase following pathogen infection in proximal and distal ends of the tissue (Malamy et al., 1990; Rasmussen et al., 1991) which mediates the development of SAR causing substantial changes in gene expression including in genes encoding pathogenesis-related proteins. Such scenario operates well where there is no endogenous production of SA but raises several questions on the operation of SA-mediated signaling and triggering of SAR in plants which have high constitutive SA levels. It is not clear whether such plants respond positively to the increase in inherent high levels of SA or the higher levels of SA become a hindrance and make the cells less competent in perceiving or transducing the SA signal as is the norm with other plant hormones (Weyers and Paterson, 2001). One school of thought believes that potato might have poor SA signal perception based on a study wherein SA could not induce resistance to Phytophthora infestans while arachidonic acid was able to induce the resistance (Yu et al., 1997). There are others who believe that some mutants of Arabidopsis with high basal levels of SA constitutively express SAR (Bowling et al., 1994) and there are suggestions that the presence of high SA levels makes potato more resistant to diseases (Vleeshouwers et al., 2000). Although potato has high amounts of total SA, it is capable of tightly regulating the amount of free SA which will allow the tissue to remain responsive to free SA (Navarre and Mayo, 2004).
There are also reports which state that SA-mediated resistance are restricted to effects in the treated tissue and when applied exogenously it does not translocate efficiently throughout the plants (Enyedi and Raskin, 1993). In addition, SA deposited on leaf surface is rapidly broken down and brings about a short term response. The safety margin that separates the rates at which SA is effective and the rate at which it becomes strongly phytotoxic is very narrow (Romero et al., 2001). These limitations have prevented the consideration of SA as a practical solution for disease control. SAR has not been adopted successfully for disease control in potato and other crops but it has the capabilities of becoming a viable strategy for disease control with the rapid increase in characterization of SA-mediated signaling in plants. Since the SA treatments improved emergence and canopy, reduced disease severity and improved tuber weight, their importance in plant disease cannot be completely ruled out. Further field trials and analysis of SA levels prior to planting and after harvest will give a better understanding about the role of SA in plant disease control.
ACKNOWLEDGMENTS
Excellent assistance of Rene Poirier, Lisa Bernier and Vikram Appanna is appreciated.
REFERENCES
- Ajilan, A.M. and D.A. Potter, 1992. Lack of effect of tobacco mosaic virus-induced systemic acquired resistance to anthropoid herbivores. Phytopathology, 82: 647-651.
Direct Link - Alexander, D., R.M. Goodman, M. Gut-Rella, C. Glascock and K. Weymann et al., 1993. Increased tolerance to two oomycete pathogens in transgenic tobacco expressing pathogenesis-related protein 1a. Proc. Natl. Acad. Sci. USA., 90: 7327-7331.
Direct Link - Anderson, N.A., 1982. The genetics and pathology of Rhizoctonia solani. Annu. Rev. Phytopathol., 20: 329-347.
CrossRefDirect Link - Bandy, B.P., S.S. Leach and S.M. Tavantzis, 1988. Anastomosis group 3 is the major cause of Rhizoctonia disease of potato in maine. Plant Dis., 73: 596-598.
CrossRefDirect Link - Banville, G.J., 1989. Yield losses and damage to potato plants caused by Rhizoctonia solani Kuhn. Am. Potato J., 66: 821-834.
CrossRefDirect Link - Bokshi, A.I., S.C. Morris and B.J. Deverall, 2003. Effects of benzothiadiazole and acetylsalicylic acid on β-1, 3-glucanase activity and disease resistance in potato. Plant Pathol., 52: 22-27.
Direct Link - Bowling, S.A., A. Guo, H. Cao, A.S. Gordon, D.F. Klessig and X. Dong, 1994. A mutation in Arabidopsis that leads to constitutive expression of systemic acquired resistance. Plant Cell, 6: 1845-1857.
CrossRefDirect Link - Cao, H., S.A. Bowling, A.S. Gordon and X. Dong, 1994. Characterization of an Arabidopsis mutant that is nonresponsive to inducers of systemic acquired resistance. Plant Cell, 6: 1583-1592.
CrossRefDirect Link - Carling, D.E. and R.H. Leiner, 1986. Isolation and characterization of Rhizoctonia solani and binucleate R. solani-like fungi from aerial stems and subterranean organs of potato plants. Phytopathology, 76: 725-729.
CrossRefDirect Link - Carling, D.E., R.H. Leiner and P.C. Westphale, 1989. Symptoms, signs and yield reduction associated with Rhizoctonia disease of potato induced by tuberborne inoculum of Rhizoctonia solani AG-3. Am. Potato J., 66: 693-701.
CrossRefDirect Link - Celetti, M.J., H.W. Johnston, J. Kimpinski, H.W. Platt and R.A. Martin, 1990. Incidence of soil-borne plant pathogens isolated from barley and winter wheat and other crops in the rotation, on Prince Edward Island. Plant Pathol., 39: 606-611.
CrossRefDirect Link - Chandra, A. and R.K. Bhatt, 1998. Biochemical and physiological response to salicylic acid in relation to the systemic acquired resistance. Photosynthetica, 35: 255-258.
CrossRef - Chan, Z. and S. Tian, 2006. Induction of H2O2-metabolizing enzymes and total protein synthesis by antagonistic yeast and salicylic acid in harvested sweet cherry fruit. Postharvest Biol. Technol., 39: 314-320.
CrossRefDirect Link - Coquoz, J.L., A.J. Buchala, P. Meuwly and J.P. Metraux, 1995. Arachidonic acid induces local but not systemic synthesis of salicylic acid and confers systemic resistance in potato plants to Phytophthora infestans and Alternaria solani. Phytopathology, 85: 1219-1224.
CrossRef - Cruickshank, G., H.E. Stewart and R.L. Wastie, 1982. An illustrated assessment key for foliage blight of potatoes. Potato Res., 25: 213-214.
CrossRefDirect Link - Delaney, T.P., S. Uknes, B. Vernooij, L. Friedrich and K. Weymann et al., 1994. A central role of salicylic acid in plant disease resistance. Science, 266: 1247-1250.
Direct Link - Delaney, T.P., 2000. New mutants provide clues into regulation of systemic acquired resistance. Trends Plant Sci., 5: 49-51.
Direct Link - Dempsey, D.A., J. Shah and D.F. Klessig, 1999. Salicylic acid and disease resistance in plants. Crit. Rev. Plant Sci., 18: 547-575.
CrossRefDirect Link - Dong, X., 2001. Genetic dissection of systemic acquired resistance. Curr. Opin. Plant Biol., 4: 309-314.
CrossRefDirect Link - Dorrance, A.E. and D.A. Inglis, 1997. Assessment of greenhouse and laboratory screening methods for evaluating potato foliage for resistance to late blight. Plant Dis., 81: 1206-1213.
CrossRefDirect Link - Elliston, J., J. Kuc, E.B. Williams and J.E. Raje, 1977. Relationship of phytoalexin accumulation to local and systemic protection of bean against anthracnose. J. Phytopathol., 88: 114-130.
CrossRefDirect Link - Enyedi, A.J., N. Yalpani, P. Silverman and I. Raskin, 1992. Localization, conjugation and function of salicylic acid in tobacco during the hypersensitive reaction to tobacco mosaic virus. Proc. Natl. Acad. Sci. USA., 89: 2480-2484.
Direct Link - Enyedi, A.J. and I. Raskin, 1993. Induction of UDP-glucose: Salicylic acid glucosyltransferase activity in tobacco mosaic virus-inoculated tobacco (Nicotiana tabacum) leaves. Plant Physiol., 101: 1375-1380.
Direct Link - Errampalli, D., W. Arsenault and K.A. MacIsaac, 1999. Efficacy of seed piece treatment fungicide, Maxim (fludioxonil), Dividend/ Maxim (difenaconazole/fludioxonil) and Easout (thiophanate-methyl) on black scurf, silver scurf and dry rot of potatoes, 1998-1999. Agric. Agri-Food Can. 1999 Pest Manage. Res. Rep. No. 113, pp: 305-307.
- Frank, J.A. and S.S. Leach, 1980. Comparison of tuberborne and soilborne inoculum in the Rhizoctonia disease of potato. Ecol. Epidemiol., 70: 51-53.
Direct Link - Gaffney, T., L. Friedrich, B. Vernooj, D. Negrotto and G. Nye et al., 1993. Requirement of salicylic acid for the induction of systemic acquired resistance. Science, 261: 754-756.
CrossRefDirect Link - Glazebrook, J., W. Chen, B. Estes, H.S. Chang, C. Nawrath and J.P. Métraux, 2003. Topology of the network integrating salicylate and jasmonate signal transduction derived from global expression phenotyping. Plant J., 34: 217-228.
Direct Link - Hammerschmidt, R. and J. Kuc, 1982. Lignification as a mechanism for induced systemic resistance in cucumber. Physiol. Plant Pathol., 20: 61-71.
CrossRefDirect Link - Jeger, M.J., G.A. Hide, P.H.J.F. Van den Boogert, A.J. Termorshuizen and P. Van Baarlen, 1996. Pathology and control of soil-borne fungal pathogens of potato. Potato Res., 39: 437-469.
CrossRefDirect Link - Kessmann, H., T. Staub, C. Hofmann, T. Maetzke and J. Herzog et al., 1994. Induction of systemic acquired disease resistance in plants by chemicals. Annu. Rev. Phytopathol., 32: 439-459.
CrossRefDirect Link - Klessig, D.F. and J. Malamy, 1994. The salicylic acid signal in plants. Plant Mol. Biol., 26: 1439-1458.
CrossRefDirect Link - Knoester, M., C.M.J. Pieterse, J.F. Bol and L.C. Van Loon, 1999. Systemic resistance in Arabidopsis induced by rhizobacteria requires ethylene-dependent signaling at the site of application. Mol. Plant-Microb. Interact., 12: 720-727.
CrossRefDirect Link - Lawton, K., K. Weymann, L. Friedrich, B. Vernooij, S. Uknes and J. Ryals, 1995. Systemic acquired resistance in Arabidopsis requires salicylic acid but not ethylene. Mol. Plant Microb. Interact., 6: 863-870.
Direct Link - Lawton, K.A., L. Friedrich, M., Hunt, K. Weymann and T. Delaney et al., 1996. Benzothiadiazole induces disease resistance in Arabidopsis by activation of the systemic acquired resistance signal transduction pathway. Plant J., 10: 71-82.
Direct Link - Linthorst, H.J.M. and L.C. Van Loon, 1991. Pathogenesis-related proteins of plants. Crit. Rev. Plant Sci., 10: 123-150.
CrossRefDirect Link - Malamy, J., J.P. Carr, D.F. Kessig and I. Raskin, 1990. Salicylic acid: A likely endogenous signal in the resistance response of tobacco to viral infection. Science, 250: 1002-1004.
CrossRefDirect Link - Malamy, J., J. Hennig and D.F. Klessig, 1992. Temperature-dependent induction of salicylic acid and its conjugates during the resistance response to tobacco mosaic virus infection. Plant Cell, 4: 359-366.
Direct Link - Maleck, K., A. Levine, T. Eulgem, A. Morgan, J. Schmid and K.A. Lawton, 2000. The transcriptome of Arabidopsis thaliana during systemic acquired resistance. Nat. Genet., 26: 403-410.
Direct Link - Metraux, J.P., H. Signer, J. Ryals, E. Ward and M. Wyss-Benz et al., 1990. Increase in salicylic acid at the onset of systemic acquired resistance in cucumber. Science, 250: 1004-1006.
CrossRefPubMedDirect Link - Namai, T., T. Kato, Y. Yamaguchi and T. Hirukawa, 1993. Anti-rice blast activity and resistance induction of C-18 oxygenated fatty acids. Biosci. Biotechnol. Biochem., 57: 611-613.
CrossRefDirect Link - Navarre, D.A. and D. Mayo, 2004. Differential characteristics of salicylic acid-mediated signaling in potato. Physiol. Mol. Plant Pathol., 64: 179-188.
Direct Link - Pérez, L., M.E. Rodríguez, F. Rodríguez and C. Roson, 2003. Efficacy of acibenzolar-S-methyl, an inducer of systemic acquired resistance against tobacco blue mould caused by Peronospora hyoscyami f. sp. tabacina. Crop Prot., 22: 405-413.
Direct Link - Pieterse, C.M.J., S.C.M. van Wees, E. Hoffland, J.A. Van Pelt and L.C. Van Loon, 1996. Systemic resistance in Arabidopsis induced by biocontrol bacteria is independent of salicylic acid accumulation and pathogenesis-related gene expression. Plant Cell, 8: 1225-1237.
Direct Link - Pieterse, C.M.J., S.C.M. Van Wees, J.A. Van Pelt, M. Knoester and R. Laan, 1998. A novel signaling pathway controlling induced systemic resistance in Arabidopsis. Plant Cell, 10: 1571-1580.
Direct Link - Platt, H.W., 1989. Potato growth and tuber production as affected by inoculation of cut and whole seed with Rhizoctonia solani (Ag-3) and the use of seed treatment fungicides. Am. Potato J., 66: 365-378.
CrossRefDirect Link - Platt, H.W., F. Canale and G. Giminéz, 1993. Effects of tuber-borne inoculum of Rhizoctonia solani and fungicidal seed potato treatment on plant growth and Rhizoctonia disease in Canada and Uruguay. Am. Potato J., 70: 553-559.
CrossRefDirect Link - Raggi, V., 1998. HydroxyproUne‐rich glycoprotein accumulation in TMV‐infected tobacco showing systemic acquired resistance to powdery mildew. J. Phytopathol., 146: 321-325.
CrossRefDirect Link - Rairdan, G.J., N.M. Donofrio and T.P. Delaney, 2001. Salicylic acid and NIM1/NPR1-independent gene induction by incompatible Perenospora parasitica in Arabidopsis. Mol. Plant Microb. Interact., 14: 1235-1246.
Direct Link - Rairdan, G.J. and T.P. Delaney, 2002. Role of salicylic acid and NIM1/NPR1 in race-specific resistance in Arabidopsis. Genetics, 161: 803-811.
Direct Link - Raskin, I., H. Skubatz, W. Tang and B. Meeuse, 1990. Salicylic acid levels in thermogenic and non-thermogenic plants. Ann. Bot., 66: 369-373.
Direct Link - Rasmussen, J.B., R. Hammerschmidt and M.N. Zook, 1991. Systemic induction of salicylic acid accumulation in cucumber after inoculation with Pseudomonas syringae pv syringae. Plant Physiol., 97: 1342-1347.
CrossRefDirect Link - Romero, A.M., C.S. Kousik and D.F. Ritchie, 2001. Resistance to bacterial spot in bell pepper induced by acibenzolar-S-methyl. Plant Dis., 85: 189-194.
Direct Link - Ryals, J.A., U.H. Neuenschwander, M.G. Willits, A. Molina, H.Y. Steiner and M.D. Hunt, 1996. Systemic acquired resistance. Plant Cell, 8: 1809-1819.
PubMedDirect Link - Saikia, R., T. Singh, R. Kumar, J. Srivastava, A.K. Srivastava, K. Singh and D.K. Arora, 2003. Role of salicylic acid in systemic resistance induced by Pseudomonas fluorescens against Fusarium oxysporum f. sp. ciceri in chickpea. Microbiol. Res., 158: 203-213.
CrossRefDirect Link - Schenk, P.M., K. Kazan, I. Wilson, J.P. Anderson and T. Richmond, 2000. Coordinated plant defense responses in Arabidopsis revealed by microarray analysis. Proc. Natl. Acad. Sci. USA., 97: 11655-11660.
Direct Link - Spoel, S.H., A. Koornneef, S.M. Claessens, J.P. Korzelius and J.A. Van Pelt et al., 2003. NPR1 modulates cross-talk between salicylate-and jasmonate-dependent defense pathways through a novel function in the cytosol. Plant Cell, 15: 760-770.
Direct Link - Sticher, L., B. Mauch-Mani and J.P. Metraux, 1997. Systemic acquired resistance. Annu. Rev. Phytopathol., 35: 235-270.
CrossRefPubMedDirect Link - Tsror, L.L. and I. Peretz-Alon, 2005. The influence of the inoculum source of Rhizoctonia solani on development of Black Scurf on Potato. J. Phytopathol., 153: 240-245.
CrossRefDirect Link - Van Loon, L.C., P.A.H.M. Bakker and C.M.J. Pieterse, 1998. Systemic resistance induced by rhizosphere bacteria. Annu. Rev. Phytopathol., 36: 453-483.
CrossRefPubMedDirect Link - Van Wees, S.C.M., E.A.M. De Swart, J.A. Van Pelt, L.C. Van Loon and C.M.J. Pieterse, 2000. Enhancement of induced disease resistance by simultaneous activation of salicylate and jasmonate-dependent defense pathways in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA., 97: 8711-8716.
Direct Link - Verberne, M.C., R. Verpoorte, J.F. Bol, J. Mercado-Blanco and H.J.M. Linthorst, 2000. Overproduction of salicylic acid in plants by bacterial transgenes enhances pathogen resistance. Nat. Biotech., 18: 779-783.
Direct Link - Vleeshouwers, V.G.A.A., W. van Dooijeweert, F. Govers, S. Kamoun and L.T. Colon, 2000. Does basal PR gene expression in Solanum species contribute to nonspecific resistance to Phytophthora infestans. Physiol. Mol. Plant Pathol., 57: 35-42.
Direct Link - Ward, E.R., S.J. Uknes, S.C. Williams, S.S. Dincher and D.L. Wiederhold et al., 1991. Coordinate gene activity in response to agents that induce systemic acquired resistance. Plant Cell., 3: 1085-1094.
Direct Link - Weyers, J.B.B. and N.W. Paterson, 2001. Plant hormones and the control of physiological processes. New Phytol., 152: 375-407.
CrossRefDirect Link - Wicks, T.J., B. Morgan and B. Hall, 1995. Chemical and biological control of Rhizoctonia solani on potato seed tubers. Aust. J. Exp. Agric., 35: 661-664.
Direct Link - Wyatt, S.E. and J. Kuc, 1992. The accumulation of β-ionone and 3-hydroxy esters of β-ionone in tobacco immunized by foliar inoculation with tobacco mosaic virus. Phytopathology, 82: 580-582.
Direct Link - Yan, Z., M.S. Reddy, C.M. Ryu, J.A. McInroy, M. Wilson and J.W. Kloepper, 2002. Induced systemic protection against tomato late blight elicited by plant growth-promoting rhizobacteria. Phytopathology, 92: 1329-1333.
CrossRefPubMedDirect Link - Yao, H. and S. Tian, 2005. Effects of pre- and post-harvest application of salicylic acid or methyl jasmonate on inducing disease resistance of sweet cherry fruit in storage. Postharvest Biol. Technol., 35: 253-262.
CrossRefDirect Link - Yu, D., Y. Liu, B. Fan, D.F. Klessig and Z. Chen, 1997. Is the high basal level of salicylic acid important for disease resistance in potato? Plant Physiol., 115: 343-349.
CrossRefDirect Link - Kloepper, J.W., S. Tuzun and J.A. Kuc, 1992. Proposed definitions related to induced disease resistance. Biocontrol Sci. Technol., 2: 349-351.
CrossRef