
Zafar Rasheed
Department of Biochemistry, Faculty of Medicine, J.N. Medical College, A.M.U., Aligarh-202002, India
Rizwan Ahmad
Department of Biochemistry, S.B.S.P.G.I., Balawala, Dehradun-248161, India
Naila Rasheed
Division of Pharmacology, Central Drug Research Institute, P.B. No. 173, Lucknow-226001, India
Trivendra Tripathi
Department of Biochemistry, Faculty of Medicine, J.N. Medical College, A.M.U., Aligarh-202002, India
Rashid Ali
Department of Biochemistry, Faculty of Medicine, J.N. Medical College, A.M.U., Aligarh-202002, India
International Journal of Biological Chemistry
Year: 2008 | Volume: 2 | Issue: 1 | Page No.: 1-13







ABSTRACT
The study hypothesized that oxidative by-product like hydrogen peroxide (H2O2), would reacts with hemoglobin (Hb) and help to initiate autoimmunity in patients with type 1 diabetes. To test this hypothesis, Hb was modified by reactive oxygen species (ROS). Modification incurred in Hb was characterized by physico-chemical techniques. ROS-modified Hb was found to be highly immunogenic in rabbits as compare to native Hb. The binding characteristics of circulating autoantibodies in type 1 diabetes patients against native and ROS-modified Hb were assessed. Diabetes patients (n = 39) were examined by direct binding ELISA and their results were compared with healthy age-matched controls (n = 22). High degree of specific binding by 48% of patient`s sera towards ROS-modified Hb, in comparison to its native analogue (p<0.05) was observed. Competitive inhibition ELISA reiterates the direct binding results. Collectively, ROS caused extensive damage to Hb altering its immunogenicity, the neo-epitopes, thus generated might play a role in the induction of circulating autoantibodies in type 1 diabetes mellitus.
PDF Abstract XML References Citation
How to cite this article
Zafar Rasheed, Rizwan Ahmad, Naila Rasheed, Trivendra Tripathi and Rashid Ali, 2008. Reactive Oxygen Species Damaged Hemoglobin Presents Unique Epitopes for Type 1 Diabetes Autoantibodies
. International Journal of Biological Chemistry, 2: 1-13.
URL: https://scialert.net/abstract/?doi=ijbc.2008.1.13
URL: https://scialert.net/abstract/?doi=ijbc.2008.1.13
INTRODUCTION
Oxidative stress has been implicated in a wide range of diseases including atherosclerosis, hypertension, diabetes, ischemia reperfusion injury and a range of neurodegenerative disorders (Flora, 2007). Metal interactions with hydrogen peroxide have been frequently implicated in catalyzing free radical-mediated damage. Redox-active transition metals, containing unpaired electrons in their d-orbital, are able to generate, or remove, reactive species with unpaired electrons. In particular free ferrous iron (Gutteridge, 1986) has been suggested to catalyze tissue damage by reacting with hydrogen peroxide to generate the highly reactive hydroxyl radical (OH•). However, the relevance of this Fenton chemistry to systems where strong redox-inactive chelaters exist to bind iron is not clear.
There has been accumulating evidence that an additional mechanism to generate reactive oxygen species occurs in vivo (Moore et al., 1998). This involves the interaction of the ferric state of heme proteins with hydrogen peroxide:
R + Fe3+ + H2O2<—> R•+ + Fe4+ = O2–+ H2O | (1) |
| Where: | ||
| R | = | Represents the polypeptide chain. |
| Fe3+ | = | Iron bound to the heme group. |
Both the ferryl iron (Fe4+ = O2–) and the protein-bound free radical (R•+) are able to initiate a range of oxidative chemistry with similar reactivity to the hydroxyl radical (Sutton and Winterbourn, 1989). Heme proteins like hemoglobin (Hb) and myoglobin (Mb) contain redox-active transition metal iron that makes them susceptible to causing oxidative damage. Although the structure of the globin chain allows heme to bind oxygen with minimal oxidation of ferrous to ferric iron, autoxidation is not entirely prevented; low concentrations of ferric heme are normally present in vivo (methemoglobin, metmyoglobin). These can then react with the peroxides formed during the autoxidation process itself or elsewhere in the protein`s vicinity. It is well documented that oxidative damage of protein and DNA presents unique neoepitopes, which is one of the factors in antigen-driven autoimmune response in various diseases (Scofield et al., 2005; Habib et al., 2006).
Recent studies from our laboratory have demonstrated that after modification with ROS, proteins became highly immunogenic in experimental animals and oxidative by-products with proteins could lead to neoantigens which would initiate autoimmunity in various diseases (Rasheed and Ali, 2006; Rasheed et al., 2006, 2007a,b). In view of these, the present study was hypothesized that oxidative by-products like hydrogen peroxide, would react with Hb and help to initiate autoimmunity in patients with type 1 diabetes mellitus. To test this hypothesis, Hb was modified by reactive oxygen species, generated by in vitro reaction of hemoglobin with hydrogen peroxide. Immunogenicity of ROS modified and unmodified Hb was investigated in rabbits and found that autoimmunity was established faster and more vigorously in the animals that were immunized against ROS-modified Hb. We have studied the presence of circulating autoantibodies in type 1 diabetes sera directed against native and damaged Hb.
MATERIALS AND METHODS
Materials
Anti-rabbit-IgG-alkaline phosphatase conjugate, p-nitrophenyl phosphate, Tween-20, hemoglobin were purchased from Sigma Chemical Company, USA Protein A-Sepharose CL-4B was purchased from Genei, India. Polystyrene microtitre flat bottom ELISA plates and modules were purchased from NUNC, Denmark. All other chemicals were of analytical grade.
Protein was estimated by the methods of Lowry et al. (1951).
Human Subjects
Blood was collected from voluntary donors with history of type 1 diabetes mellitus (n = 39, male = 23, female = 16, aged 49±16) under treatment in J.N. Medical College Hospital A.M.U., Aligarh. The control samples (n = 22, male = 17, female = 5, aged 43±17) were collected from healthy subjects. The average (±SD) of post prandial blood sugar level for 31 diabetic patients was 354±84 mg dL-1, while it was 110±8 mg dL-1 for 22 healthy subjects (without family history of diabetes). The glycated hemoglobin (HbA1C) was 8.0±1.5 for patients, whereas 4.7±0.4 for healthy subjects. Glutamic Acid Decarboxylase Antibodies (GADA), insulin antibodies (IAA), anti-tyrosine phosphatase antibodies (IA-2A) were found in 43, 74 and 39% of the cases, respectively. Immunopositivity for at least one of these autoantibodies was found in all patients. Fifty one percent male patients were having smoking history. All other information on demographics and cumulative clinical and laboratory manifestations over the course of disease was obtained by both chart review and discussion with the patient and his/her family members. The informed consent was obtained from each patient or from patients family members.
All sera were decomplemented by heating at 56°C for 30 min and stored in aliquots at -20°C with sodium azide as preservative.
Modification of Hemoglobin
Hemoglobin from Sigma was used exactly in a similar way as cited by various investigators (Shimada et al., 2005). Hb (7.8 μM) and hydrogen peroxide (2.1 mM) were mixed and incubated at 37°C for 12 h. After incubation, the excess hydrogen peroxide was removed by extensive dialysis against PBS (10 mM sodium phosphate buffer containing 150 mM NaCl, pH 7.4). Hb incubated without hydrogen peroxide served as control. The UV absorption characteristics of native and modified Hb were recorded on a Shimadzu spectrophotometer.
Fluorescence Measurements
Fluorescence measurements were performed on Hitachi F-200 spectrofluorimeter (Japan). The fluorescence spectra were measured at 25±0.1°C with a cell of 1 cm path length. The excitation and emission slits were set at 5 and 10 nm, respectively. Intrinsic fluorescence was measured by exciting the protein solution at 280 or 295 nm and emission spectra were recorded in the range of 300-400 nm. Increase of fluorescence intensity (F.I.) was calculated using the following equation.
Increase of F.I. (%) = [(F.I.Modified Hb-F.I. Native Hb)/F.I.Modified Hb] x 100 |
Absorption-Temperature Scan
Thermal denaturation profile of protein was performed on a Shimadzu UV-240 spectrophotometer equipped with a temperature programmer and controller assembly (Hasan and Ali, 1990). Protein samples were in PBS, pH 7.4 and absorbance was recorded at a fixed wavelength of 280 nm. Percent denaturation was calculated using the equation:
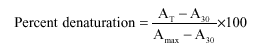 |
| Where: | ||
| AT | = | Absorbance at a temperature T°C. |
| Amax | = | Final maximum absorbance on the completion of denaturation. |
| A30 | = | Initial absorbance at 30°C. |
Assay of Carbonyl Formation
Carbonyl contents of native and ROS-modified Hb were analyzed according to Levine et al. (1990) with slight modifications. The reaction mixture containing 15.6 μM native Hb or ROS-Hb, 0.5 mL of 10 mM 2, 4-dinitrophenylhydrazine (DNPH)/2.5 M HCl was added and thoroughly mixed. After addition of 250 μM TCA (20%) and centrifugation, the pellet was collected and washed three times with 1 mL ethanol: Ethylacetate (1:1) mixture. The pellet was then dissolved in 1 mL of 6 M guanidine solution and incubated at 30°C for 15 min. After centrifugation, the supernatant was collected and carbonyl contents were estimated from the absorbance at 370 nm using a molar absorption coefficient of 22,000 M-1 cm1. Samples were spectrophotometrically analyzed against a blank of 1 mL of guanidine solution (6 M). Protein concentration was determined in the samples by the method of Lowry et al. (1951). Carbonyl contents were expressed as nmol mg-1 protein.
Immunization Schedule
The immunization of random bred, New Zealand white rabbits was performed as described previously (Rasheed et al., 2006). Briefly, rabbits (n = 4; two each for native and ROS-Hb antigens) were immunized intramuscularly at multiple sites with 100 μg of antigen, emulsified with an equal volume of Freund`s complete adjuvant. The animals were boosted in Freund`s incomplete adjuvant at weekly intervals for 6 weeks with the same amount of antigen. Test bleeds were performed 7 days post boost, which gave appropriate titre of the antibody. The animals were bled and the serum separated from the blood was heated at 56°C for 30 min to inactivate complement proteins and stored at -20°C.
Purification of IgG
Immunoglobulin G (IgG) was isolated from immune and preimmune rabbit sera on a protein A-sepharose CL-4B column (Rasheed and Ali, 2006). The homogeneity of isolated IgG was checked by polyacrylamide gel electrophoresis.
Enzyme Linked Immunosorbent Assay
Enzyme linked immunosorbent assay (ELISA) was carried out on polystryrene plates with slight modification (Rasheed and Ali, 2006; Rasheed et al., 2006, 2007a, b). Polystryrene polysorp immunoplates were coated with 100 μL of native or modified Hb (10 μg mL-1) in carbonate-bicarbonate buffer (0.05 M, pH 9.6). The plates were coated for 2 h at 37°C and overnight at 4°C. Each sample was coated in duplicate and half of the plates served as control devoid of only antigen coating. Unbound antigen was washed thrice with TBS-T (20 mM Tris, 150 mM NaCl, pH 7.4 containing 0.05% Tween-20) and unoccupied sites were blocked with 2% fat free milk in TBS (10 mM Tris, 150 mM NaCl, pH 7.4) for 4-6 h at 37°C. After incubation the plates were washed four times with TBS-T. The test serum serially diluted in TBS-T or affinity purified IgG in TBS (100 μL well-1) was adsorbed for 2 h at 37°C and overnight at 4°C. Bound antibodies were assayed with anti-rabbit alkaline phosphatase conjugate using p-nitrophenyl phosphate as substrate. The absorbance (A) of each well was monitored at 410 nm on an automatic microplate reader. Each sample was run in duplicate. The control wells were treated similarly but were devoid of antigen. Results were expressed as a mean of Atest-Acontrol.
Competition ELISA
The antigenic specificity of the antibodies was determined by competition ELISA (Rasheed et al., 2006; Rasheed and Ali, 2006). Varying amounts of inhibitors (0-20 μg mL-1) were mixed with a constant amount of antiserum or purified IgG. The mixture was incubated at room temperature for 2 h and overnight at 4°C. The immune complex thus formed was coated in the wells instead of the serum/IgG. The remaining steps were the same as in direct binding ELISA. Percent inhibition was calculated using the formula:
Percent inhibition = 1-(Ainhibited/Auninhibited) x 100 |
Statistical Analysis
Data are presented as mean±SD. Significance of differences from control values were determined with the Student`s t-test (Statgraphics, Origin 6.1). A value of p<0.05 was considered to indicate statistical significance.
RESULTS
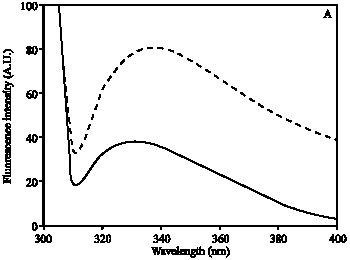
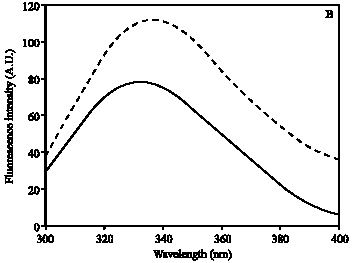
Hb was modified by ROS and the induced changes in Hb were analyzed by UV-visible absorption spectroscopy. The absorption spectra of ROS damaged Hb showed 64% hypochromicity at 280 nm. A sharp peak at 410 nm was observed, which is characteristic of native Hb. Upon ROS modification, peak at 410 nm was completely disappeared (Fig. 1). The oxidation of tryptophan residues on damaged Hb was evident by the increase of 51% fluorescence intensity at 330 nm using an excitation wavelength of 295 nm (Fig. 2A). The damage to tryptophan residues were confirmed by the increase of 28% fluorescence intensity at 330 nm using an excitation wavelength of 280 nm (Fig. 2B). Stability of native and modified Hb was checked by temperature-induced denaturation experiments, the protein absorbance changes with temperature until the process of unfolding is complete. Thermal melting profile of native and ROS-damaged Hb showed melting temperature (Tm) of native and modified HAS as 59 and 65.5°C, respectively (Fig. 3). The complete unfolding of native and ROS-damaged Hb was occurred at 75 and 80°C, respectively. After these temperatures both protein samples showed precipitation or aggregation as abnormal increase in their absorbance was noticed. Oxidation of protein typically results in an increase in carbonyl contents, a known biomarker of oxidative stress. The oxidation of ROS-Hb was further evident by the estimation of carbonyl contents. The average carbonyl contents (±SD) of five independent assays of ROS-Hb and native Hb were 28.7±1.2 and 10.4±0.7 nmol mg-1 protein, respectively, a p-value of<0.001 indicates significant difference in the carbonyl contents of ROS-Hb and native Hb.
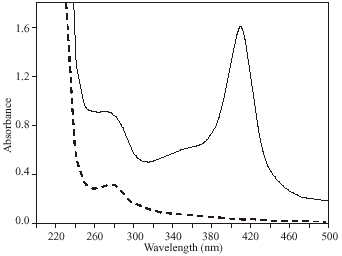 | |
| Fig. 1: | UV-visible spectra of native (—) and ROS-modified (---) Hb. The samples were in PBS, pH 7.4 at a concentration of 7.8 μM. The spectra are the average of three determinations |
| Fig. 2: | Fluorescence emission spectra of native (—) and ROS-modified (---) Hb. Protein was in PBS, pH 7.4 at a concentration of 1.5 μM. The excitation wavelength was 295 nm (A) and 280 nm (B). The spectra are the average of three determinations |
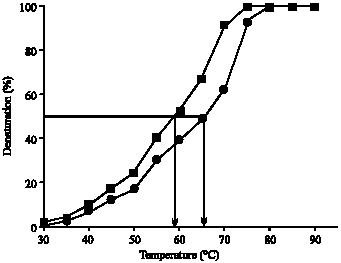 | |
| Fig. 3: | Thermal denaturation profile of native (■) and ROS-modified (●) Hb. The samples were in PBS, pH 7.4 at a concentration of 3 μM |
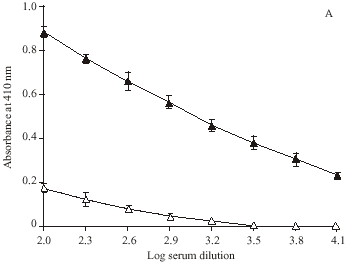
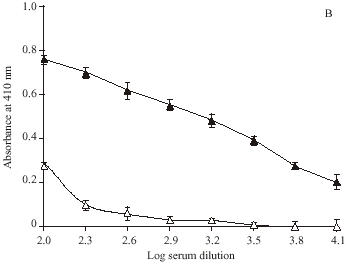
Direct binding ELISA showed ROS-modified Hb as a potent immunogen in rabbits inducing high titre (> 1: 12800) antibodies, whereas with native Hb the titre was low (~ 1: 6400). Preimmune serum serve as negative control, as it did not showed appreciable binding with the respective immunogens under identical experimental conditions (Fig. 4).
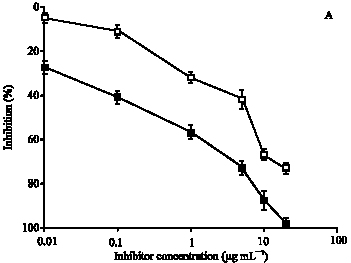
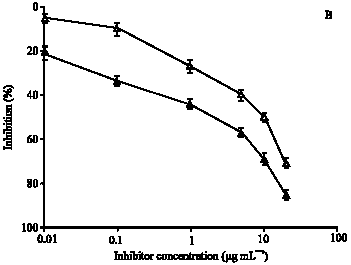
To substantiate the above results, immunoglobulin G (IgG) against ROS-Hb and Hb were isolated on Protein-A Sepharose CL-4B affinity column. IgG from preimmune sera was also purified under identical experimental conditions. The purified IgG was found to elute in a single symmetrical peak. SDS-PAGE under non-reducing conditions of affinity purified IgG showed a single homogenous band (data not shown). Direct binding ELISA of purified anti-ROS-Hb IgG and anti-Hb IgG showed strong binding with their respective immunogens. Preimmune IgG showed negligible binding under identical conditions (data not given). The specificity of the purified IgG was further evaluated by competitive inhibition assays. Affinity purified anti-ROS-Hb IgG showed the average percent inhibition (±SD) of 98±2.2 at 20 μg mL-1 of inhibitor (ROS-Hb) concentration. The concentration of competitor (ROS-Hb) required for 50% inhibition was only 0.5 μg mL-1. However, with native Hb a maximum of 73±2.2 inhibition of the anti-ROS-Hb IgG was observed. The microtitre plates were coated with ROS-Hb (Fig. 5A). Whereas, anti-Hb IgG showed the average percent inhibition (±SD) of 85±2.2 at 20 μg mL-1 of inhibitor (native Hb) concentration. The concentration of competitor (native Hb) required for 50% inhibition was 2 μg mL-1, ROS-Hb showed average percent inhibition 71±2.2 of anti-Hb IgG. The microtitre plates were coated with native Hb (Fig. 5 B).
Preimmune IgG with ROS-Hb and Hb showed negligible inhibition under identical experimental conditions (data not given). The results were similar to those obtained with serum and point to that autoimmunity was established faster and more vigorously in the animals that were immunized with ROS-Hb.
| Fig. 4: | Level of induced antibodies against ROS-modified and native Hb. Direct binding ELISA of (A) ROS-Hb and (B) Hb with immune (▲) and preimmune (Δ) sera. Microtitre plates were coated with the respective antigen (10 μg mL-1). Each point represents a mean±SD of three independent assays |
Present study comprised serum samples from patients suffering from type 1 diabetes mellitus (DM). Each sample was obtained after careful clinical examination of patients with proven diagnosis at J.N. Medical College Hospital, A.M.U., Aligarh. Sera from thirty nine type 1 DM and twenty two normal human subjects were tested for binding to native and ROS modified Hb by direct binding ELISA (Table 1). A majority of DM sera (19/39) showed strong binding to ROS-Hb over native Hb at 1:100 serum dilution (p<0.05). No appreciable binding was observed with the normal subjects. The average absorbance at 410 nm (±SD) of 19 DM sera binding to native and ROS-damaged Hb was 0.47±0.06 and 0.83±0.07, respectively. Whereas, the absorbance at 410 nm (±SD) of 22 normal human sera binding to both protein antigens was 0.24±0.04. The binding specificity of antibodies from 19 selected sera of DM patients was evaluated by competition ELISA using native and ROS-Hb as inhibitors. Mircrotitre plates were coated with ROS-Hb. Results point to a higher reactivity of autoantibodies in patient`s sera towards ROS damaged Hb over native Hb. Similarly, rest of the sera showed high percent inhibitions with modified Hb over native Hb. The average percent inhibition (±SD) in the binding of 19 type 1 DM sera to native and ROS-Hb was 27.6±5.6 and 49.3±9.9, respectively. The data reveals striking differences in the recognition of native and oxidized Hb by DM autoantibodies (p<0.001). Table 1 summarizes complete demographics, clinical, biochemical and immunological characteristics of all 19 selected DM sera with their controls. Among 19 DM sera, 11 sera were from those male patients which are having smoking history of 22.4±10.4 years. Direct binding and competition ELISA results point to a strong reactivity of all DM smokers towards ROS-Hb over native Hb. Sera from aging patients with high degree of diabetes showed substantially higher binding to ROS-Hb as compare to native Hb (Table 1). Present results are in full agreement to the view that oxidative damage of protein presents unique epitopes, which help to initiate autoimmunity (Scofield et al., 2005; Rasheed and Ali, 2006; Rasheed et al., 2006, 2007a, b).
| Fig. 5: | (A) Inhibition of anti-ROS-Hb IgG binding to ROS-Hb. The inhibitors were ROS-Hb (■) and native Hb (∼). The microtitre plates were coated with the ROS-Hb (10 μg mL-1). (B) Inhibition of anti-Hb IgG binding to native Hb. The inhibitors were native Hb (▲) and ROS-Hb (Δ). The microtitre plates were coated with the native Hb (10 μg mL-1). Each point represents a mean±SD of three independent assays |
DISCUSSION
ROS are continuously generated in cells by cellular metabolism and exogenous agents but increase in their steady states is attributed to the variety of pathological conditions, including cardiovascular diseases, cancer, SLE and aging (Ahsan et al., 2003). Excess generation of ROS has the ability, either directly or indirectly, to damage proteins, DNA and other cell biomolecules (Rasheed and Ali, 2006). In fact, proteins are major targets for free radical attack. Protein oxidation results in cellular dysfunctions, functional disruption and structural changes (Arutyunova et al., 2003) and contribute to the etiology of many human diseases (Chou, 2002). These oxidative modifications on protein may lead to the formation of neoantigens which could in turn initiate autoimmunity (Scofield et al., 2005; Rasheed and Ali, 2006; Rasheed et al., 2006, 2007a, b).
| Table 1: | Biochemical and immunological detailed of study subjects |
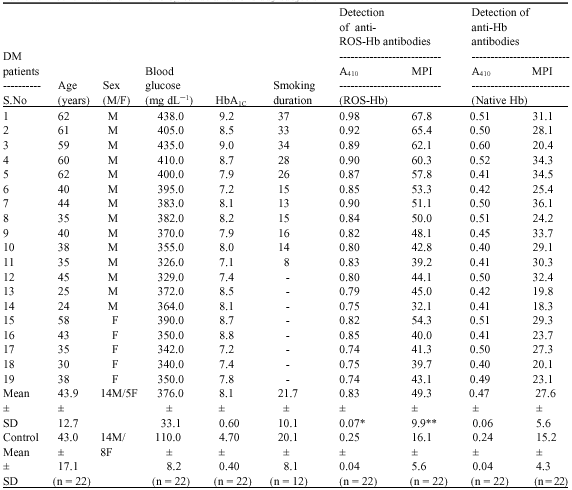 | |
| DM: Type 1 diabetes mellitus; M: Male; F: Female; n: Number of samples tested; DM-serum: Serum from DM patients; A410: Absorbance at 410 nm calculated by direct binding ELISA; MPI: Maximum percent inhibition at 20 μg mL-1 of inhibitor concentration calculated by competitive inhibition ELISA. *: p<0.05 vs. A410 (native Hb); **: p<0.001 vs. MPI (native Hb) | |
The extracellular concentration of hemoglobin in plasma is 3-6 mg L-1 and is usually eliminated by renal filtration or hepatic capture (Faivre et al., 1998). Plasma hemoglobin concentration can increase considerably as a result of hemolysis induced by drugs and environmental hazards or in patients given hemoglobin based blood substitutes, which might saturate these systems (Alayash, 1999). Under these conditions, hemoglobin can react with H2O2 to generate tissue damage (Yoo et al., 1999). Normally, tissue and plasma H2O2 levels are quite low, but it has been estimated that the production of H2O2 by activated macrophages can reach concentrations as high as 100-600 μM (Grisham et al., 1990). In the bloodstream, H2O2 can be generated by autoxidation of oxyHb (Giulivi et al., 1994), platelet activation (Iuliano et al., 1992), macrophage activation, or xenobiotic metabolism (Kindt et al., 1992).
In the present study commercially available hemoglobin (Hb) was modified by ROS, generated by in vitro reaction of Hb and H2O2. UV-visible spectra showed extensive damage to Hb. The observed hypochromicity at 280 nm could be due to the modification of chromophoric groups, modification of aromatic amino acid residues of Hb or due to the structural alteration. A sharp peak at 410 nm was observed due to the soret absorption by the heme system. Upon ROS modification the soret peak was completely lost. Oxidation of tryptophan residues on ROS modification was confirmed by the increase in fluorescence intensity when the protein is excited at 295 nm. The changes in tryptophan were further confirmed by exciting the protein at 280 nm. Thermal denaturation profile showed unfolding of ROS-modified Hb as it requires high temperature as compared to native Hb. The result indicates that stability of Hb increases upon ROS modification. Protein carbonyl groups are actually the most general indicator and by far the most commonly used marker of protein oxidation (Beal, 2002). The oxidation of ROS-Hb was further substantiated by the significant (p<0.001) increase in carbonyl contents. This increase is due to the oxidation of lys, arg, pro or many other amino acid residues (Dalle-Dome et al., 2003).
It is well established that oxidative damage of protein or DNA leads to various autoimmune diseases (Scofield et al., 2005). This is because of oxidative modification of protein or DNA that might result in the formation of chemical adducts which could serve as neoantigens that the immune system has probably not been exposed to. There are other examples of an immune response against oxidatively modified self-antigens. Aldehyde-modified proteins were found to be highly immunogenic and autoantibodies directed against epitopes. In malondiadehyde– (MDA) and HNE-modified Low Density Lipoprotein (LDL). These have been demonstrated in the plasma of rabbits and mice immunized with oxidized LDL (ox-LDL) (Palinski et al., 1990). Antibodies against ox-LDL or MDA-LDL have been demonstrated in atherosclerotic plaques (Bergmark et al., 1995). Itabe et al. (1998) have shown that MDA-cross linked proteins are actually present in vivo and that their production is stimulated by oxidative stress. Antigens modified by oxidative by-products have been shown to induce immune responses in alcoholic liver diseases (Mottaran et al., 2002). Oxidative stress occurs when antioxidant mechanisms are not overwhelmed in some ways, possible due to pathology in the disease. Hydrogen peroxide is generated by number of ways (Grisham et al., 1990); these peroxides are normally eliminated by glutathione which acts as a reducing agent. Moreover, hemoglobin sulfhydryl groups can no longer be maintained in the reduced form when glutathione reductase becomes inactive, Hb molecules then cross-link with one another to form aggregates called Heinz bodies and reactive oxygen species become deformed and the cell is likely to undergo lysis. Reduced glutathione is essential to maintain normal structure of red blood cells and for keeping Hb in the ferrous state. The reduced form also plays a role in detoxification by reacting with hydrogen peroxide and organic peroxide. Cells with a lowered level of reduced glutathione are more susceptible to hemolysis (Berg et al., 2002).
In this study, ROS modified Hb was found to be a potent antigenic stimulus inducing high titre antibodies in rabbits, whereas native Hb results in low titre antibodies. The antigenic specificity of Protein-A purified anti-ROS-Hb IgG and anti-Hb IgG reiterated that induced antibodies are immunogen specific. Our results are in full agreement to the view that the substantially enhanced immunogenicity of ROS-Hb in comparison to native Hb could possibly be due to the generation of potential neoepitopes (Rasheed and Ali, 2006). However, the induced antibodies against ROS-Hb and native Hb also showed cross-reactivity with native and ROS-modified Hb. The analysis of cross-reactivity indicates that anti-ROS-Hb IgG and anti-Hb IgG are immunogen-specific, showing various extents of cross-reactivity attributable to sharing of common antigenic determinants.
Oxidative stress plays a central role in the onset of Diabetes Mellitus (DM) as well as in diabetic associated complications (Schleicher and Friess, 2007). The role of oxidative protein damage in the pathogenesis of the diabetic state is being investigated extensively (Telci et al., 2000). Hb has close relation with DM, several reports have been made on glycation-induced by structural and functional modification of Hb (Thomas et al., 2007). In type 1 DM, autoantibodies are present in patient`s sera that bind to oxidatively modified islet cell protein (Telci et al., 2000). We have recently reported the recognition of free radical damaged human serum albumin by type 1 DM autoantibodies (Rasheed and Ali, 2006). It was thought worthwhile to investigate the binding characteristics of naturally occurring autoantibodies of type 1 DM to ROS-modified Hb. Autoantibodies from thirty nine different type 1 DM sera were screened by direct binding and competitive inhibition ELISA. Out of these, 48% sera showed preferentially high binding towards ROS-Hb as compared to native Hb (p<0.001). No detectable antibody activity was found in normal human sera. Our results demonstrate the presence of ROS-induced Hb damage in type 1 DM patients which may be due to the failure of reductase system in disease state, this might play an active part in the progression of disease. The present study clearly demonstrates structural perturbation in Hb by reactive oxygen species, rendering it immunogenic. The neo-epitopes thus generated might play a role in the induction of circulating autoantibodies in autoimmune diseases like type 1 diabetes mellitus.
ACKNOWLEDGMENTS
Author thanks Prof. Jamal Ahmad (Department of Medicine, J.N. Medical College, A.M.U. Aligarh) for providing most of the type 1 diabetes mellitus samples. This study was supported in part by a research grants from Department of Biochemistry, J.N.M.C. Aligarh Muslim University, Aligarh-202002, India.
REFERENCES
- Ahsan, H., A. Ali and R. Ali, 2003. Oxygen free radicals and systemic autoimmunity. Clin. Exp. Immunol., 131: 398-404.
Direct Link - Alayash, A.I., 1999. Hemoglobin-based blood substitutes: Oxygen carriers, pressor agents, or oxidants? Nat. Biotechnol., 17: 545-549.
Direct Link - Arutyunova, E., I., P.V. Danshina, L.V. Domnina, A.P. Pleten and V.I. Muronetz, 2003. Oxidation of glyceraldehyde-3-phosphate dehydrogenase enhances its binding to nucleic acids. Biochem. Biophys. Res. Commun., 307: 547-552.
CrossRefDirect Link - Beal, M.F., 2002. Oxidatively modified proteins in aging and disease. Free Radic. Biol. Med., 32: 797-803.
CrossRefDirect Link - Bergmark, C., R. Wu, U. de Faire, A.K. Lefvert and J. Swedenborg, 1995. Patients with early-onset peripheral vascular disease have increased levels of autoantibodies against oxidized LDL. Arterioscler. Thromb. Vasc. Biol., 15: 441-445.
Direct Link - Chou, C., 2002. Binding of rheumatoid and lupus synovial fluids and sera-derived human IgG rheumatoid factor to degalactosylated IgG. Arch. Med. Res., 33: 541-544.
CrossRefDirect Link - Dalle-Dome, I., R. Rossi, D. Giustarini, A. Milzani and R. Colombo, 2003. Protein carbonyl groups as biomarkers of oxidative stress. Clin. Chim. Acta, 29: 23-38.
CrossRefDirect Link - Faivre, B., P. Menu, P. Labrude and C. Vigneron, 1998. Hemoglobin autooxidation/oxidation mechanisms and methemoglobin prevention or reduction processes in the bloodstream literature review and outline of autooxidation reaction. Art Cells Blood Subs. Immob. Biotechnol., 26: 17-26.
CrossRefDirect Link - Flora, S.J., 2007. Role of free radicals and antioxidants in health and disease. Cell Mol. Bio., 53: 1-2.
PubMed - Giulivi, C., P. Hochstein and K.J.A. Davies, 1994. Hydrogen peroxide production by red blood cells. Free Radic. Biol. Med., 16: 123-129.
CrossRefDirect Link - Grisham, M.B., T.S. Gaginella, C. Von Ritter, H. Be. Tamai and D.N. Granger, 1990. Effects of neutrophil-derived oxidants on intestinal permeability, electrolyte transport and epithelial cell viability. Inflammation, 14: 531-542.
CrossRefPubMedDirect Link - Gutteridge, J.M.C., 1986. Iron promoters of the Fenton reaction and lipid peroxidation can be released from haemoglobin by peroxides. FEBS Lett., 201: 291-295.
CrossRefDirect Link - Habib, S., Moinuddin and R. Ali, 2006. Peroxynitrite-modified DNA: A better antigen for systemic lupus erythematosus anti-DNA autoantibodies. Biotechnol. Applied Biochem., 43: 65-70.
PubMedDirect Link - Hasan, R. and R. Ali, 1990. Antibody recognition of common epitopes on Z-DNA and native DNA brominated under high salt. Biochem. Int., 20: 1077-1188.
Direct Link - Itabe, H., S. Jimi, S. Kamimura, K. Suzuki and N. Uesugi et al., 1998. Appearance of cross linked proteins in human atheroma and rat pre-fibrotic liver detected by a new monoclonal antibody. Biochim. Biophys. Acta, 406: 28-39.
CrossRefDirect Link - Iuliano, L., F. Violi, J.Z. Pedersen, D. Pratico, G. Rotilio and F. Balsano, 1992. Free radical-mediated platelet activation by hemoglobin released from red blood cells. Arch. Biochem. Biophys., 299: 220-224.
CrossRefDirect Link - Kindt, J.T., A. Woods, B.M. Martin, R.J. Cotter and Y. Osawa, 1992. Covalent alteration of the prosthetic heme of human hemoglobin by BrCCl3. Cross-linking of heme to cysteine residue 93. J. Biol. Chem., 267: 8739-8743.
PubMedDirect Link - Levine, R.L., D. Garland, C.N. Oliver, A. Amici and I. Climent et al., 1990. Determination of carbonyl content in oxidatively modified proteins. Meth. Enzymol., 186: 464-478.
CrossRefPubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Moore, K.P., S. Holt, R.P. Patel, D.A. Svistunenko and W. Zackert et al., 1998. Causative role for redox cycling of myoglobin and its inhibition by alkalinization in the pathogenesis and treatment of rhabdomyolysis-induced renal failure. J. Biol. Chem., 273: 31731-31737.
PubMedDirect Link - Mottaran, E., S.F. Stewart, R. Rolla, D. Vay and V. Cipriani et al., 2002. Lipid peroxidation contributes to immune reactions associated with alcoholic liver disease. Free Radic. Biol. Med., 32: 38-45.
CrossRefDirect Link - Palinski, W., S. Yla-Herttuala, M.E. Rosenfeld, S. Butler and S.A. Socher et al., 1990. Antisera and monoclonal antibodies specific for epitopes generated during oxidative modification of low density lipoprotein. Atherosclerosis, 10: 325-335.
PubMedDirect Link - Rasheed, Z. and R. Ali, 2006. Reactive oxygen species damaged human serum albumin in patients with type 1 diabetes mellitus: Biochemical and immunological studies. Life Sci., 79: 2320-2328.
CrossRefDirect Link - Rasheed, Z., M.W.A. Khan and R. Ali, 2006. Hydroxyl radical modification of human serum albumin generated cross reactive antibodies. Autoimmunity, 39: 479-488.
CrossRefPubMedDirect Link - Rasheed, Z., R. Ahmad, N. Rasheed and R. Ali, 2007. Reactive oxygen species damaged human serum albumin in patients with hepatocellular carcinoma. J. Exp. Clin. Cancer Res., 26: 395-404.
Direct Link - Rasheed, Z., R. Ahmad, N. Rasheed and R. Ali, 2007. Enhanced recognition of reactive oxygen species damaged human serum albumin by circulating systemic lupus erythematosus autoantibodies. Autoimmunity, 40: 512-520.
CrossRefDirect Link - Schleicher, E. and U. Friess, 2007. Oxidative stress, AGE and atherosclerosis. Kidney Int. Suppl., 106: S17-S26.
CrossRefPubMedDirect Link - Scofield, R.H., B.T. Kurien, S. Granick, M.J. McClain and Q. Pye et al., 2005. Modification of lupus-associated 60-kDa Ro protein with the lipid oxidation product 4-hydroxy-2-nonenal increases antigenicity and facilitates epitopes spreading. Free Radic. Biol. Med., 38: 719-728.
CrossRefDirect Link - Shimada, S., Y. Tanaka, C. Ohmura, Y. Tamura and T. Shimizu et al., 2005. N-(carboxymethyl) valine residues in hemoglobin (CMV-Hb) reflect accumulation of oxidative stress in diabetic patients. Diabetes Res. Clin. Prac., 69: 272-278.
CrossRefDirect Link - Sutton, H.C. and C.C. Winterbourn, 1989. On the participation of higher oxidation states of iron and copper in Fenton reactions. Free Radic. Biol. Med., 6: 53-60.
CrossRefDirect Link - Telci, A., U. Cakatay, S. Salman, I. Satman and A. Sivas, 2000. Oxidative protein damage in early stage Type 1 diabetic patients. Diabetes Res. Clin. Pract., 50: 213-223.
CrossRefDirect Link - Thomas, L.B., S.J. Agosti, M.A. Man and S.M. Mastorides, 2007. Screening for hemoglobinopaties during routine hemoglobin a1c testing using the tosoh g7 glycohemoglobin analyzer. Ann. Clin. Lab. Sci., 37: 251-255.
PubMedDirect Link - Yoo, Y.M., K.M. Kim, S.S. Kim, J.A. Han, H.Z. Lea and Y.M. Kim, 1999. Hemoglobin toxicity in experimental bacterial peritonitis is due to production of reactive oxygen species. Clin. Diagn. Lab. Immunol., 6: 938-945.
PubMedDirect Link