
Ibrahim Abdullah Alaraidh
Department of Botany and Microbiology, Science College, King Saud University, P.O. Box 2455, 11451 Riyadh, Saudi Arabia
LiveDNA: 966.24958
International Journal of Botany
Year: 2018 | Volume: 14 | Issue: 1 | Page No.: 30-35







ABSTRACT
Background and Objective: Balanites aegyptiaca, belonging to family Zygophyllaceae, is multipurpose tree considered highly drought-tolerant but salt-sensitive and cultivated in several arid and tropical regions in North and west Africa and West Asia. The plant parts contain several secondary metabolites and there is a high oil content in the kernels already used in biodiesel production. This study was aimed at investigating effect of the genotypic variation in B. aegyptiaca plants (collected from different geographical locations, Riyadh, Saudi Arabia and Halayeb, Egypt) on the efficiency of the genetic transformation process for both ERD10 and nptII genes. Materials and Methods: Two B. aegyptiaca genotypes collected from two different geographical regions with different ecological conditions (Halayeb, Egypt and Riyadh, Saudi Arabia) were used in this study. The genetic transformation was performed using Agrobacterium tumefacienss train GV3101 harboring the binary vector pBinAR containing ERD10 (Early Responsive to Dehydration 10) and nptII genes, to produce salt-tolerant B. aegyptiaca plants. Results: The results showed that each genotype collected from the different geographical locations after transformation exhibited significant differences in the number of leaves per regenerated explant and shoot length, where by nodal explants from Halayeb, Egypt exhibited the highest values. Conclusion: Current findings emphasize the impact of the genotypic variation (geographical location) of the used plant on the efficiency of the genetic transformation process.
PDF Abstract XML References Citation
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Ibrahim Abdullah Alaraidh, 2018. Genotypic Variation Affects the Efficiency of the Genetic Transformation Process in Balanites aegyptiaca. International Journal of Botany, 14: 30-35.
DOI: 10.3923/ijb.2018.30.35
URL: https://scialert.net/abstract/?doi=ijb.2018.30.35
DOI: 10.3923/ijb.2018.30.35
URL: https://scialert.net/abstract/?doi=ijb.2018.30.35
INTRODUCTION
Balanites aegyptiaca (L.) Del, commonly called the desert date, is a xerophytic multipurpose tree species belonging to family Zygophyllaceae and is considered a highly drought-tolerant plant species found naturally in arid and semi-arid regions in north and west Africa, west Asia and the Arabian Peninsula1. Chapagain et al.2 reported that oil content represents approximately 47% of the kernel and this oil is effectively analyzed for biodiesel production after esterification. Several parts of B. aegyptiaca (seeds, leaves, roots and fruits) show biological activities, such as antibacterial, anticancer and antifungal activities for medical treatment and oral contraceptive production, owing to the presence of various secondary metabolites, apart from saponins, which are successfully used in detergent production, including foaming agents2-4.
Salinity has a negative impact on plant productivity and food production and salt stress affects 800 million ha of agricultural land worldwide5. Despite the high capacity of B. aegyptiaca as a drought-tolerant plant, B. aegyptiaca seedlings show high sensitivity to salt stress at low levels (12 dS m–1). However, salt stress at high levels (24 dS m–1) also showed harmful effects on seedling growth6. Furthermore, salt stress negatively affects the biomass and growth of three different B. aegyptiaca showing significantly differences in responses to salt stress7. Therefore, there is a demand for improving salt tolerance in B. aegyptiaca to cultivates it in arid and semi-arid regions with various soil types. Kovacs et al.8 reported that under abiotic stress, such as salt and drought stress, accumulation of early responses to dehydration proteins (ERD10), which are members of the Dehydrin family, occurs. The chaperone activity of ERD10 might protect proteins by preventing the inactivation and heat-induced aggregation of various proteins under stress9. Several constraints affect the propagation of woody plant species. Nevertheless, numerous classical techniques were used for propagating B. aegyptiaca using roots, suckers, seeds and cuttings; however, these methods showed limitations for the mass propagation of this plant species, owing to slow growth and low germination rate of seeds10. The use of biotechnological techniques, such as in vitro propagation, for trees has shown considerable potential for large-scale multiplication, apart from the improvement and conservation of elite clones11. Several efforts were made for the in vitro propagation of B. aegyptiaca using nodal explants with axillary buds, nodal and cotyledonary parts10,12,13 somatic embryos through roots segments14 callus induction were obtained through roots and cotyledon explants and apical buds15. Khamis et al.13 reported the first reporton the transformation of B. aegyptiaca using three different Agrobacterium tumefaciens strains (EHA105, GV3101 and LBA4404), harboring the pCAMBIA2301 plasmid containing the nptII marker and gus reporter genes, where strain GV3101 showed the highest transformation efficiency. In addition, they transferred ERD10 gene into B. aegyptiaca producing a salt tolerant transformed plant. However, B. aegyptiaca plants collected from different locations revealed genetic variation with a high percentage of polymorphism16,17. Transformation efficiency to introduce ERD10 gene into B. aegyptiaca plants of different genotypes has not been investigated. This study was aimed at investigating effect of the genotypic variation in B. aegyptiaca plants (collected from different geographical locations, Halayeb, Egypt and Riyadh, Saudi Arabia) on the efficiency of the genetic transformation process for both ERD10 and nptII genes using Agrobacterium tumefaciens containing the pBinAR vector harboring ERD10.
MATERIALS AND METHODS
Balanites aegyptiaca need 6 months for regeneration and acclimatization. So, this experiment was conducted between June and December, 2017.
Plant material and culture conditions: Seeds of B. aegyptiaca were collected from two different geographical locations, Saudi Arabia (Riyadh) and Egypt (Halayeb). The seeds were mechanically released from the fruits. The seeds were sterilized with sodium hypochlorite (9.4% active chlorine) for 40 min and washed four times with sterilized water. For germination, the seeds were placed on Murashige and Skoog (MS) medium supplemented with 3% (w/v) sucrose and 0.4% Phytagel and pH was adjusted to 5.7 for 4 weeks18 and incubated in a growth chamber at 24°C with a 16:8 h light:dark photoperiod. After germination, shoot segments containing single node were excised for use as nodal explants. Preparation of Agrobacterium tumefaciens and explants for transformation.
The binary vector pBinAR harboring ERD10 13 was transformed by the freeze/thaw shock method of transformation19 into A. tumefaciens strain GV3101. The cells were spread on 2× yeast extract-tryptone (2-YT) plates supplemented with rifampicin (50 mg L–1) and kanamycin (50 mg L–1). On the second day of incubation, the transformed colonies of strain GV3101 were subjected to plasmid isolation and the presence of ERD10 (800 bp) was detected by PCR as described by Khamis et al.13.
Preparation of nodal explants for transformation and regeneration conditions: At the 4th week of germination, the nodal explants were excised from aseptic seedlings of B. aegyptiaca from Halayeb, Egypt (E) and Riyadh, Saudi Arabia (S). Preparation of nodal explants for transformation was conducted according to Khamis et al.13. For acclimatization, healthy plantlets with shoots and roots (Two transformed plantlets from Riyadh, Saudi Arabia and three from Halayeb, Egypt) were transferred under greenhouse conditions to pots filled with soil (natural clay and peat, 2:1) and covered with plastic bags to maintain humidity and watered every two days with half-strength MS medium for two weeks. Then, the plastic bags were removed to allow the plantlets to adapt for another two weeks.
Molecular analysis: Polymerase Chain Reaction (PCR) was conducted according to Khamis et al.13 for detecting the presence of ERD10 and nptii in the transformed plants. Leaves were taken from regenerated, transformed plants and DNA was extracted by the cetyltrimethylammonium bromide (CTAB) method. The following primers were used: nptii5'ATGGCTAAAATGAGAATA3' as the forward primer and nptii 5'CTAAAACAATTCATCCAG3' as the reverse primer. The amplification of the 800 bp PCR fragment of ERD10 through PCR was performed using the following primers: Forward primer, 5'GGTACCATGGCTGAAGAGTACAAG3' and reverse primer, 5'GGATCCTCATCCTTCTAAATCATCGG3'.
Statistical analysis: Approximately, 150 nodal explants were used in the transformation experiment for both genotypes from both locations in Saudi Arabia and Egypt. Different analytical measurements for the biological samples were performed in duplicate and the results were expressed as Means±SE. Independent t-tests were performed to determine whether the differences between the two groups were statistically significant (p<0.05) using SPSS 17 for Windows.
RESULTS
Sterilized seeds (150) of each B. aegyptiaca genotype were placed on MS medium without PGR; after 4 weeks of germination, the seeds from Egypt (Halayeb) showed a higher germination percentage than those from Saudi Arabia (Riyadh). The transformed nodal explants from each location were responsive for shoot formation (Fig. 1a-d), when placed on MS medium supplemented with 8.8 μM BA and kanamycin (100 mg L–1) as the selected marker. The transformed nodal explants from Riyadh exhibited a higher percentage of the number of explants produced shoots (37%) than those from Halayeb (33%).
The responses of the transformed plantlets from each location were significantly different (Table 1) after 4 weeks of inoculation on MS medium supplemented with 8.8 μM BA, 1.3 μM NAA and kanamycin (200 mg L–1). The transformed plantlets from Halayeb showed the highest number of shoots per regenerated explants (3.4±0.4) and highest shoot length (1.6±0.8) and they were significantly higher than those from Riyadh (0.96±0.16 and 0.44±0.04) for the number of shoots per regenerated explants and shoot length, respectively (Table 1). Root induction was observed when the transformed plantlets were moved to MS medium supplemented with 1.2 μM NAA and kanamycin (200 mg L–1). Only three plants from Halayeb and two plants from Riyadh produced roots and survived at a high concentration of kanamycin (200 mg L–1) and then, these plants from each location were transferred to the soil for acclimatization.
 | |
| Fig. 1(a-d): | Multiple shoot formation in B. aegyptiaca from Halayeb, Egypt using nodal explants after transformation with A. tumefaciens, (a) Nodal explant on the first day, (b) Multiple shoot induction on MS+8.8 μM BA after 3 weeks, (c) Elongation and proliferation of multiple shoots on MS+8.8 μM BA+1.3 μM NAA after 8 weeks and (D) In vitro rooted plantlet on MS+1.2 μM NAA |
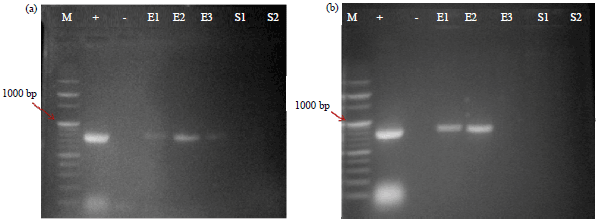 | |
| Fig. 2(a-b): | PCR products of (a) ERD10 (800 bp) and (b) nptII (750 bp) |
| M: One kbp DNA ladder, +: Binary vector pBinAR containing ERD10 and nptII and -: Leaves excised from non-transformed B. aegyptiaca plants. E1, E2, E3: Leaves of B. aegyptiaca from Egypt, S1, S2: Leaves of B. aegyptiaca from Saudi Arabia. Both B. aegyptiaca plant genotypes were transformed using. A. tumefaciens strain GV3101 harboring the binary vector pBinAR containing ERD10 and nptII | |
| Table 1: | Frequencies of shoot regeneration for the two B. aegyptiaca genotypes after transformation with A. tumefaciens |
| Values represent Means±SE, Means followed by *Within a column differed significantly after Tukey’s test (p<0.05) | |
Leaves of the three transformed plants from Halayeb, Egypt (E), two from Riyadh, Saudi Arabia (S) and non-transformed (-) B. aegyptiaca plants were subjected to PCR analysis. The presence of ERD10 was detected in the transformed plants (E1, E2 and E3) from Halayeb, Egypt (Fig. 1a). Furthermore, the presence of nptII was detected in the transformed plants (E1 and E2) from Halayeb, Egypt (Fig. 2b). However, both the transformed plants from Riyadh, Saudi Arabia (S) and non-transformed plants did not show the respective fragments (Fig. 2a, b).
DISCUSSION
In this study, B. aegyptiaca seeds collected from two different geographic locations, Saudi Arabia (Riyadh) and Egypt (Halayeb) were studied to investigate the responses of the two B. aegyptiaca genotypes to genetic transformation. The transformation process was conducted using A. tumefaciens strain GV3101 harboring the binary vector pBinAR containing ERD10 and nptII to increase the salt tolerance in B. aegyptiaca through axillary shoot formation from nodal explants. The BA was effective in inducing shoot formation in the two genotypes. The obtained results are in agreement with those reported in the previous studies on B. aegyptiaca10, 12,20. Balanites sources collected from different locations showed genetic variation and revealed high percentage of polymorphism17. The relation between genetic variation in B. aegyptiaca and geographical locations was investigated by Chamberlain16, showing that there is a high variability in the response to the expression of peroxidase isozyme between plants from different locations. The results showed that after transformation, the seeds collected from the different geographical locations exhibited no significant differences in the number of explants produced shoots between the two locations. Nevertheless, significant differences between the plants collected from the two locations in the number of shoots per regenerated explant and shoot length and the highest values were recorded in the plants from Halayeb, Egypt. This variation might be related to the genotypic variation between the seeds, which were collected from two different locations. However, B. aegyptiaca is a woody plant species considered partially auto-compatible and exhibited a cross-pollination rate of approximately 37%, which could be related to wind and insects12,21. Moreover, the plant species. Aegyptiaca is classified into varieties (ferox, pallida, quarrei and tomentosa), which are highly variable in their visible characteristics related to attributes such as distribution, flowers, fruits and preferred soil type22. High intraspecific genetic diversity was reported for the woody species with outcrossing breeding systems and they are distributed in wide geographical ranges23. Only three transformed plants from Halayeb and two from Riyadh produced roots and survived. The effects of genotype on multiplication rate were investigated in Juglansregia, where by certain clones showed a high rooting percentage (95%) and few had a low percentage (5%) 24. The presence of both ERD10 and nptII was confirmed via PCR analysis of leaves excised from the plants from Halayeb; however, the transformed plants from Riyadh and non-transformed plants did not exhibit the respective fragments. Variation in genetic transformation among genotypes or varieties within the same plant species was reported for different wheat cultivars, whereby Gemmiza 10 and Gemmiza 9 exhibited 80 and 50% positive PCR results for nptII, respectively25. Liu et al. 26 reported that, there were significant variations in transformation efficiency among five banana (Musa spp.) varieties, wherein the highest transformation efficiency was recorded for the Gongjiao variety (9.81±0.29a) and the Baxi variety exhibited the lowest transformation efficiency (1.16±0.15c). Chateau et al.27 showed that several Arabidopsis ecotypes showed differences in their susceptibility to Agrobacterium-mediated transformation and their types of responses to pre-cultivation. Under abiotic stress, such as cold and salt stress, leaves of Brassica napus L. exhibit high dehydrin ERD10 expression as a mechanism to overcome salt stress28. Moreover, ERD10 plays a substantial role in seed development and increases the capability of plants to cope with stress; furthermore, the dehydrin ERD10 is important for tolerance to dehydration9,29.
CONCLUSION
The obtained findings emphasize the impact of the genotypic variation of the used plant on the efficiency of the genetic transformation process. However, further investigations are required to study the expression of the transformed genes and the ability of the transformed plant to survive under different salinity levels. This study may help in the production of salt-tolerant plants which can be cultivated in arid and semi-arid lands and may help in some biotechnological industries such as the biodiesel production.
SIGNIFICANCE STATEMENT
This study may help in the production of salt-tolerant B. aegyptiaca plants which can be cultivated in arid and semi-arid lands which may help in some biotechnological industries such as the biodiesel production.
ACKNOWLEDGMENT
The plasmid construce ERD10 is kindly provided by Dr. Galal Khamis. Lecturer at National Institute of Laser Enhanced Sciences. Cairo University, Egypt.
REFERENCES
- Chapagain, B.P., Y. Yehoshua and Z. Wiesman, 2009. Desert date (Balanites aegyptiaca) as an arid lands sustainable bioresource for biodiesel. Bioresour. Technol., 100: 1221-1226.
CrossRefPubMedDirect Link - Tahir, A.I., A.M. Ibrahim, G.M.H. Satti, T.G. Theander, A. Kharazmi and A.S. Khalid, 1998. The potential antileishmanial activity of some Sudanese medicinal plants. Phytother. Res., 12: 576-579.
Direct Link - Mohamed, A.M., W. Wolf and W.E.L. Spieβ, 2002. Physical, morphological and chemical characteristics, oil recovery and fatty acid composition of Balanites aegyptiaca Del. Kernels. Plant Foods Hum. Nutr., 57: 179-189.
CrossRefDirect Link - Peleg, Z., M.P. Apse and E. Blumwald, 2011. Engineering salinity and water-stress tolerance in crop plants: Getting closer to the field. Adv. Bot. Res., 57: 405-443.
CrossRefDirect Link - Radwan, U.A., I. Springuel, P.K. Biswas and G. Huluka, 2000. The effect of salinity on water use efficiency of Balanites aegyptiaca (L.) Del. Egypt. J. Biol., 2: 1-7.
Direct Link - Elfeel, A.A., Z.H. Sherif and R.A. Abohassan, 2013. Stomatal conductance, mineral concentration and condensed tannin in three Balanites aegyptiaca (L.) Del. intra-specific sources affected by salinity stress. J. Food Agric. Environ., 11: 466-471.
Direct Link - Kovacs, D., E. Kalmar, Z. Torok and P. Tompa, 2008. Chaperone activity of ERD10 and ERD14, two disordered stress-related plant proteins. Plant Physiol., 147: 381-390.
CrossRefDirect Link - Kim, S.Y. and K.H. Nam, 2010. Physiological roles of ERD10 in abiotic stresses and seed germination of Arabidopsis. Plant Cell Rep., 29: 203-209.
CrossRefDirect Link - Anis, M., A. Varshney and I. Siddique, 2010. In vitro clonal propagation of Balanites aegyptiaca (L.) Del. Agrofor. Syst., 78: 151-158.
CrossRefDirect Link - Ndoye, M., I. Diallo and Y.K. Gassama/Dia, 2003. In vitro multiplication of the semi-arid forest tree, Balanites aegyptiaca (L.) Del. Afr. J. Biotechnol., 2: 421-424.
CrossRefDirect Link - Khamis, G., T. Winkelmann, F. Schaarschmidt and J. Papenbrock, 2016. Establishment of an in vitro propagation and transformation system of Balanites aegyptiaca. Plant Cell Tissue Organ Cult., 125: 457-470.
CrossRefDirect Link - Saharan, V., R.C. Yadav, N.R. Yadav and Z. Wiesman, 2011. Somatic embryogenesis and plant regeneration of Balanites aegyptiaca Del (L.): An industrial important arid tree. J. Cell Tissue Res., 11: 2529-2534.
Direct Link - Gour, V.S., S.K. Sharma, C.J.S.K. Emmanuel and K. Tarun, 2007. A rapid in vitro morphogenesis and acclimatization protocol for Balanites aegyptiaca (L) Del-a medicinally important xerophytic tree. J. Plant Biochem. Biotechnol., 16: 151-153.
CrossRefDirect Link - Khamis, G., F. Schaarschmidt and J. Papenbrock, 2017. Genetic diversity among populations of the xerophytic tree species Balanites aegyptiaca and its morpho-physiological responses to water deficiency. Afr. J. Agric. Res., 12: 3252-3269.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Cohen, S.N., A.C.Y. Chang and L. Hsu, 1972. Nonchromosomal antibiotic resistance in bacteria: Genetic transformation of Escherichia coli by R-factor DNA. Proc. Nat. Acad. Sci. USA., 69: 2110-2114.
Direct Link - Siddique, I. and M. Anis, 2009. Direct plant regeneration from nodal explants of Balanites aegyptiaca L.(Del.): A valuable medicinal tree. New For., 37: 53-62.
CrossRefDirect Link - Sands, M.J.S., 2001. The desert date and its relatives: A revision of the genus balanites. Kew. Bull., 56: 1-128.
Direct Link - Hamrick, J.L., M.J. Godt and S.L. Sherman-Broyles, 1992. Factors influencing levels of genetic diversity in woody plant species. New Forests, 6: 95-124.
CrossRefDirect Link - Moghaieb, R.E.A., N.I. El-Arabi, O.A. Momtaz, S.S. Youssef and M.H. Soliman, 2010. Genetic transformation of mature embryos of bread (T. aestivum) and pasta (T. durum) wheat genotypes. GM Crops, 1: 87-93.
Direct Link - Liu, J., P. Gao, X. Sun, J. Zhang and P. Sun et al., 2017. Efficient regeneration and genetic transformation platform applicable to five Musa varieties. Electron. J. Biotechnol., 25: 33-38.
CrossRefDirect Link - Chateau, S., R.S. Sangwan and B.S. Sangwan‐Norreel, 2000. Competence of Arabidopsis thaliana genotypes and mutants for Agrobacterium tumefaciens‐mediated gene transfer: Role of phytohormones. J. Exp. Bot., 51: 1961-1968.
CrossRefDirect Link - Deng, Z., Y. Pang, W. Kong, Z. Chen and X. Wang et al., 2005. A novel ABA-dependent dehydrin ERD10 gene from Brassica napus. DNA Sequence, 16: 28-35.
CrossRefDirect Link - Caruso, A., D. Morabito, F. Delmotte, G. Kahlem and S. Carpin, 2002. Dehydrin induction during drought and osmotic stress in Populus. Plant. Physiol. Biochem., 40: 1033-1042.
CrossRefDirect Link