
Sherif M. Sharawy
Department of Botany, Faculty of Science, Ain Shams University, Abbaseya 11566, Cairo, Egypt
International Journal of Botany
Year: 2013 | Volume: 9 | Issue: 1 | Page No.: 18-29







ABSTRACT
Taxonomic status of some members of the subfamily Asclepiadoideae (Apocynaceae) was analysed using variation of morphological features, seed protein electrophoretic pattern and molecular RAPD-PCR markers. The relationships between the examined taxa have been expressed as UPGMA trees, based on the coefficient of similarity using the NTSYS-pc software program. In all trees, the species of tribe Ceropegieae have been delimited together as one group from another major group that comprises species of tribe Asclepiadeae. The present work supports the earlier classification that transferred the genus Ceropegia from subtribe Stapeliinae to subtribe Ceropegiinae. The delimitation of species of tribe Asclepiadeae in four groups supports their previous subtribal delimitation.
PDF Abstract XML References Citation
Received: November 15, 2012;
Accepted: March 09, 2013;
Published: June 27, 2013
How to cite this article
Sherif M. Sharawy, 2013. Taxonomic Relationships of Some Taxa of Subfamily Asclepiadoideae (Apocynaceae) as Reflected by Morphological Variations and Polymorphism in Seed Protein and RAPD Electrophoretic Profile. International Journal of Botany, 9: 18-29.
DOI: 10.3923/ijb.2013.18.29
URL: https://scialert.net/abstract/?doi=ijb.2013.18.29
DOI: 10.3923/ijb.2013.18.29
URL: https://scialert.net/abstract/?doi=ijb.2013.18.29
INTRODUCTION
Robert Brown (Brown, 1810, 1811) separated the Asclepiadeae (Asclepiadaceae) from the Apocineae (Apocynaceae) of Antoine-Laurent de (1789). That separation was based on the morphological characters of androecium, pollen transfer system, gynoecium and the more or less presence of extra whorl of corona attached to petal which is present in Asclepiadaceae and lacking in Apocynaceae (Al Nawaihi et al., 2006). On the other hand, new evidence from more detailed and extensive morphological and palynological evidences as well as the rapidly growing body of molecular information (Nilsson et al., 1993; Struwe et al., 1994; Endress and Albert, 1995; Sennblad and Bremer, 1996; Civeyrel et al., 1998; Sennblad et al., 1998; Potgieter, 1999; Livshultz, 2010), suggests that Brown's delimitation does not reflect natural relationships and support the recognition of a single entity. Endress and Bruyns (2000) concluded that the Asclepiadaceae is an apomorphic derivative of the Apocynaceae; thus making the latter family monophyletic and are better considered Asclepiadaceae as a subfamily of the latter; a view which was also held by Angiosperm Phylogeny Group (2003).
In the subfamily Asclepiadoideae Liede and Albers (1994) recognized five tribes: Fockeeae H. Kunze, Meve and Liede, Marsdenieae Benth., Stapelieae Decne., Gonolobeae Reichb. ex Don and Asclepiadeae (R. Br.) Duby. Six years later, Endress and Bruyns (2000) reduced the tribes to three by abandoning Fockeeae and placing it into the tribe Marsdenieae and abandoning Gonolobeae and placing it within tribe Asclepiadeae. Further, the name Ceropegieae was adopted for the tribe formerly known as Stapelieae. The current three tribes in the subfamily Asclepiadoideae, in their treatment are thus: Marsdenieae, Ceropegieae and Asclepiadeae.
The tribal subdivision of subfamily Asclepiadoideae is based largely on the organization of the androecium (Huber, 1983; Endress and Bruyns, 2000); the pollinaria are directly attached to the corpusculum as in Marsdenieae or attached to the corpusculum via caudicles, additional arm-like appendages of the translator that are synapomorphic for the other two tribes of the subfamily (Asclepiadeae and Ceropegieae). The pollinia in the pollen sacs are oriented upwardly in tribes Ceropegieae and Marsdenieae or horizontally to pendulous in tribe Asclepiadeae in relation to translator (Endress and Bruyns, 2000). In addition to the orientation of the pollen sacs, the morphology of the anther (whether or not embedded in the tissue of the anther wings) and the position of anther wings with respect to the anther sacs were suggested as supplementary characters for tribal classification of the family. The characters of the gynoecium, particularly the presence or absence of true styles and the sharp constriction between stigma-head and ovaries have also been suggested as useful in differentiating Asclepiadeae and Ceropegieae (Swarupanandan et al., 1996).
The variations in the electrophoretic profile of storage seed protein SDS-PAGE have been found useful in the study of systematics and evolution of plant species (Ladizinsky and Hymowitz, 1979; Vaughan, 1983) and may provide an understanding of the phylogenetic relationships of some taxa (Bergner and Jensen, 1989). However, valid assessment of taxonomic relationships among species and higher taxonomic ranks should necessarily be obtained when these data are considered with other lines of evidence particularly from morphology and cytology (Badr, 1995; Badr et al., 1998; Khalifa et al., 1998; Albers and Meve, 2002; Sharawy, 2008; Sharawy and Badr, 2008).
In recent years molecular data have been increasingly applied to resolve taxonomic and phylogenetic problems in plant systematics. In Asclepiadoideae, Meve and Liede (2004) analyzed the relationships of Ceropegieae and Marsdenieae, the two Asclepiadoideae tribes possessing erect pollinia by molecular investigation of non-coding cpDNA markers (trnT-L and trnL-F spacers), and the trnL intron. More recently, the PCR based approach Randomly Amplified Polymorphic DNA (RAPD) was widely used to address taxonomic issues in the subfamily. Goyder et al. (2007) studied relationships in subtribe Asclepiadinae and Jiu-Xiang et al. (2007) studied the affinities of 12 species of Apocynaceae. Mahmood et al. (2010) employed RAPD technique using 10 primers to assess genetic diversity and inter-specific relationships between two species of genus Caralluma that belong to tribe Ceropegieae.
In the present research work variation in morphological characters, polymorphism in seed protein electrophoretic profile as revealed by SDS-PAGE and the DNA molecular characters as generated by RAPD markers are applied to reassess the taxonomic relationships of 27 samples representing 13 genera and 17 species of subfamily Asclepiadoideae (Apocynaceae), in the light of their previous taxonomic treatments.
MATERIALS AND METHODS
The taxa used in this study, were collected from their natural habitats in Egypt and Saudi Arabia and from public gardens in Egypt and Saudi Arabia during the period 2006-2009. The tribe delimitation and localities of the examined materials are given in Table 1. The studied species were identified according to Tackholm and Drar (1974), Mandaville (1990), Moslem (1999), Collenette (1999) and Boulos (1999). Herbarium specimens of the examined taxa are deposited at the Herbarium of Botany Department, Faculty of Science, Ain Shams University, Cairo, Egypt and at the Museum of Biology Department, Faculty of Science, Hail University, Hail, Saudi Arabia.
| Table 1: | A list of the examined taxa of the subfamily asclepiadoideae, their tribe delimitation and their localities |
 | |
A total of 44 morphological characters were considered, the examined characters including 24 two-state characters and 20 multi-state characters. A list of these characters and their states are given in Table 2.
For protein extraction, 0.2 g seeds were powdered and mixed with 2 ml Tris-HCl buffer (Tris-SDS, β-mercaptoethanol and PMSF at pH = 8) for 1 h at room temperature. The mixture was centrifuged for 10 min at 12,000 rpm and 50 μL of supernatant (protein extract) was electrophoresed in 12.5% acrylamide that was prepared as described by Laemmli (1970) in Consort vertical slab gel apparatus. Gels were then stained for 16 h in sufficient amount of Comassie blue and distained for 12 h. the banding profile of the examined species was photographed while the gels were wet. The number of bands was scored by direct observation of gels and photograph. Each band was considered as a character and its presence or absence was coded for analysis.
RAPD-PCR reactions were conducted using 10 random primers from OPC series were tested for the amplification of DNA.
| Table 2: | A list of examined morphological characters and their codes for numerical analysis |
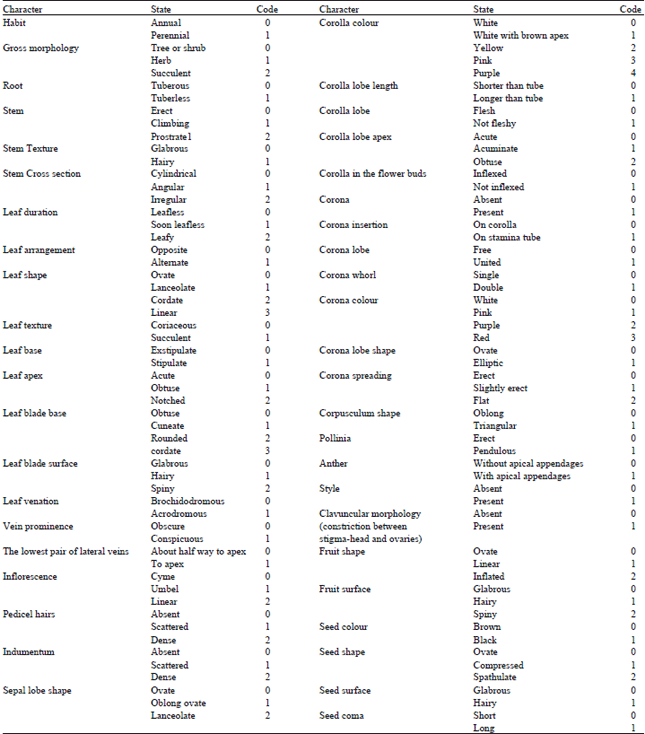 | |
The conditions reported by Khanuja et al. (1999) and Michiels et al. (2003) were used for RAPD-PCR. Amplification was achieved in a Techne (UK) Progene thermocycler programmed as follows: 94°C/4 min (1 cycle); 94°C/1 min, 37°C/1 min, 72°C/2 min (44 cycles); 72°C/7 min (1 cycle) and 4°C (infinitive). After the cycling was completed, 15 μL of the reaction products were analysed alongside small molecular weight markers on a 2% agarose gel in the presence of ethidium bromide and photographed under UV light.
For data analysis, the NTSYS-pc version 2.2 programs (Rohlf, 2000) was used. The most common use of this program is for performing various types of agglomerative cluster analyses of some type of similarity or dissimilarity matrix. The relationships between the examined taxa have been expressed using the coefficient of similarity proposed by Dice (1945) as calculated by the program. Construction of trees illustrating the relationships between the studied taxa was constructed using the Unweighted Pair Group Method using Arithmetic Average (UPGMA) proposed by Sokal and Michener (1958).
RESULTS
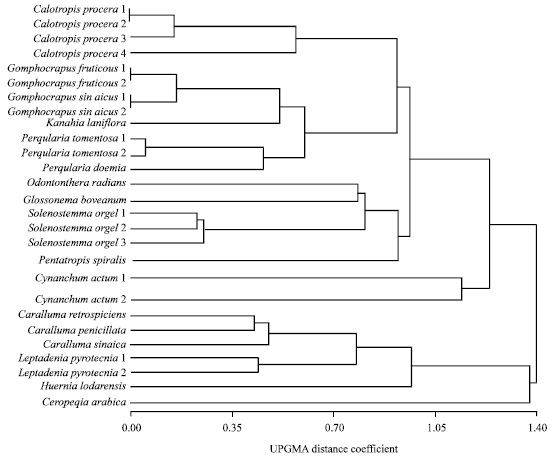
Relationships based on morphological attributes: The UPGMA cluster analysis illustrating the relationships based on morphological attributes is shown in Fig. 1. In this tree, seven taxa representing the six species of tribe Ceropegieae are delimited together as a separate group from another major group that comprises the 20 taxa representing 11 species of tribe Asclepiadeae at a UPGMA distance coefficient of about 1.40. In the former group, the Ceropegia arabica is clearly distinguished from the other three species of the same tribe (Ceropegieae) at distance coefficient about 1.04. At distance coefficient of 0.90 the two samples of Leptadenia pyrotechnica are separated from the other species of the same tribe. Also, at distance coefficient of about 0.8 the Huernia lodarensis is separated from the three species of Caralluma. The second group comprising the species of tribe Asclepiadeae is divided into two subgroups; the first includes the eight taxa representing five species and the second comprises 12 taxa representing six species. The former subgroup is delimited at distance coefficient of about 1.15 into two clusters; one comprising Pentatropis spiralis, Solenostemma argel, (three samples), Odonthanthera radians and Glossonema boveanum and the second comprising the two samples of Cynanchum acutum. In first cluster, Pentatropis spiralis is clearly differentiated from the other species at taxonomic distance coefficient of about 1.10. The second subgroup includes 12 taxa representing six species and is divided into two clusters at taxonomic distance coefficient of about 0.75, the first comprising Prgularia daemia, P. tomentosa (2 samples), Kanahia laniflora, Gomphocarpus sinaicus (two samples) and G. fruticosus (two samples) while the second cluster comprising the four samples of Calatropis procera.
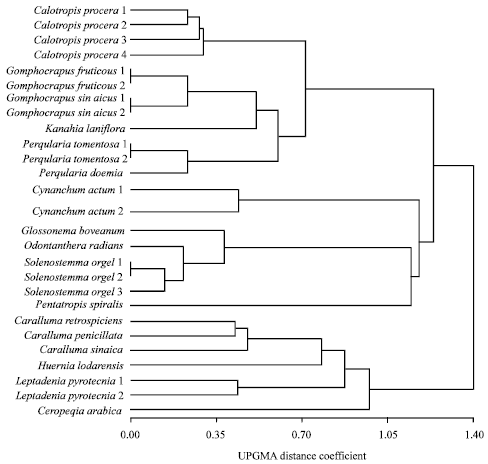 | |
| Fig. 1: | UGMA tree illustrating the relationship between the studied samples of subfamily Asclepiadoideae, based on variation in morphological characters |
Relationships based on attributes of seed protein electrophoresis: A total of 52 protein bands have been revealed in the electrophoretic profiles of the examined taxa of subfamily Asclepiadoideae (Plate 1). The molecular size of the bands scored ranges between over 100 KDa to about 10 KDa; the majority of the bands have been found polymorphic across the examined taxa. A UPGMA tree illustrating the relationships between the examined species based on the polymorphism in the seed protein electrophoretic profile is shown in Fig. 2. The topology of this tree generally resembles that of the tree based on morphological criteria. The examined species are delimited in two groups, one comprising seven taxa representing six species (Leptadenia pyrotechnica, Ceropegia arabica, Huernia lodarensis, Caralluma sinaica, C. retrospiciens and C. penicillata) and the other group includes 20 taxa representing the remaining 11 species. In the former cluster the two samples of Leptadenia pyrotechnica are clearly distinguished from the other species at a UPGMA distance coefficient of about 1.20. Ceropegia arabica and Hurenia lodarensis are separated from the three species of Caralluma at taxonomic distance coefficient of about 1.00 and 0.95 spontaneously. The second group is divided into two subgroups; the first includes eight taxa representing five species delimited in one small cluster comprised of the samples of Cynanchum acutum groups and another cluster comprising Pentatropis spiralis, Solenostemma argel (three samples), Odontanthera radians and Glossonema boveanum. The second subgroup cluster comprises 12 taxa representing six species and is divided into two clusters, a small one comprising the 4 samples of Calotropis procera. The second cluster comprises Pergularia daemia, P. tomentosa (two samples), Kanahia laniflora, Gomphocarpus sinaicus (two samples) and G. fruticosus (two samples).
Relationships based on attributes of RAPD fingerprinting: The bands produced by 10 of the 20 used random primers ranges in size from 500-2000 bp. Some of the bands were monomorphic but the majority of bands are polymorphic (Plate 2).
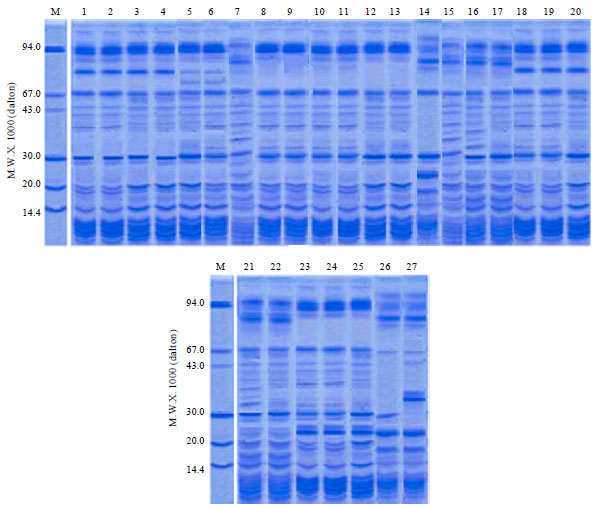 | |
| Plate 1: | Photographs of polyacrylamide gel illustrating electrophoretic band profiles of seed proteins of the studied taxa extracted in Tris-HCl buffer (Numbers of taxa are as in (Table 1) |
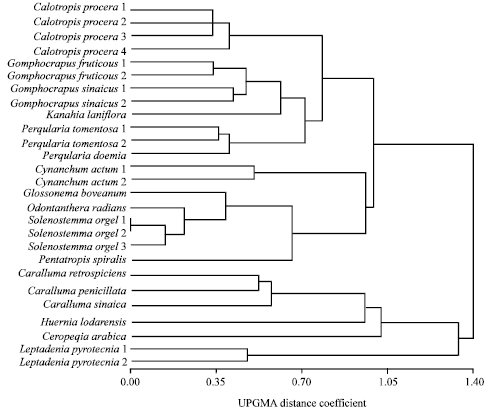 | |
| Fig. 2: | UGMA tree illustrating the relationship between the studied samples of subfamily Asclepiadoideae, based on polymorphism in seed protein electrophoretic profiles |
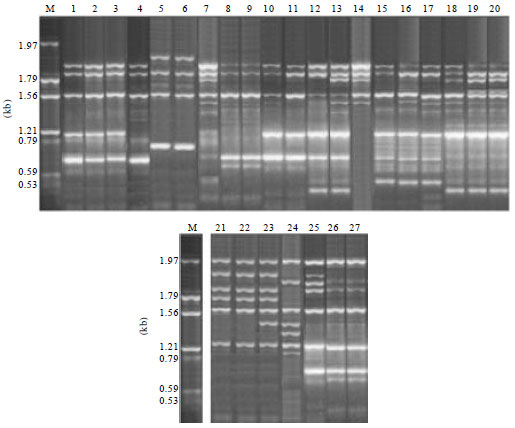 | |
| Plate 2: | PCR-RAPD pattern of the studied taxa using different primers, (No. of taxa are as in Table 1) |
The dendogram based on RAPD-PCR divided the studied taxa into two main groups; the first includes seven taxa representing the six species of tribe Ceropegieae and the second comprises the 20 taxa representing 11 species of tribe Asclepiadeae (Fig. 3). In the first group Ceropegia arabica and Huernia lodarensis are separated from the remaining species at high distance coefficient of 1.40 and 1.00, respectively. The remaining species in this group are divided into two subgroups the first comprising the two samples of Leptadenia pyrotechnica and the second group including the three species of Caralluma. In the second group, the two samples of Cynanchum acutum are separated from the remaining species at a distance of about 1.10. The remaining species are then separated into two subgroups at a distance coefficient of about 1.00. In the first subgroups Pentatropis spiralis is separated from the other species in this cluster at a distance of 0.85. The species in the first subgroups is divided into clusters; one is comprised of three samples of Solenostemma argel and the second includes the two species Odontanthera radians and Glossonema boveanum. The second subgroups comprises 12 taxa representing six species and is divided into two clusters; the first comprises Pergularia daemia, P. tomentosa (2 samples), Kanahia laniflora, Gomphocarpus sinaicus (two samples) and G. fruticosus (two samples) and comprises the four samples of Calotropis procera.
Relationships based on morphological variations and polymorphism in seed protein electrophoretic profile and RAPD fingerprinting: The overall relationship between the examined taxa based on variation in morphological criteria and polymorphism in seed protein electrophoretic profile and RAPD fingerprinting is illustrated by the UPGMA tree shown in Fig. 4. In this tree, the studied taxa are also divided in two groups; one comprising the seven taxa representing the six species of tribe Ceropegieae and the second comprises the 20 taxa representing 11 species of tribe Asclepiadeae at UPGMA distance coefficient of about 1.37. In the first group Ceropegia arabica is separated from the remaining species at high distance coefficient of about 1.20. The remaining species in this group are divided into two subgroups; the first comprising the two samples of Leptadenia pyrotechnica and the second includes the three species of Caralluma and Huernia lodarensis. In the second group, the two samples of Cynanchum acutum are separated from the remaining species at a distance of about 1.06.
 | |
| Fig. 3: | UPGMA tree illustrating the relationships between the studied species based on analysis of RAPD fingerprinting |
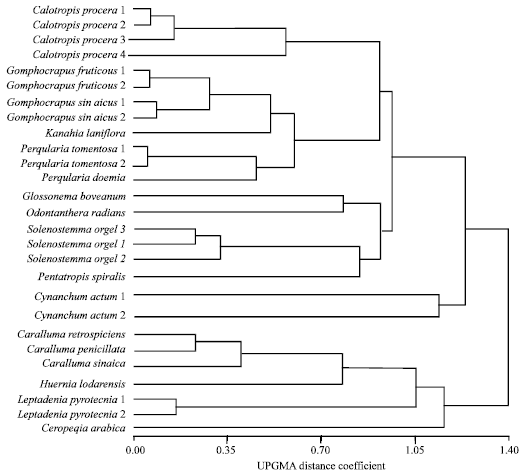 | |
| Fig. 4: | UPGMA tree illustrating the relationships between the studied species based on variation in morphological characters, polymorphism in seed protein electrophoretic profiles and RAPD analysis |
The remaining species are separated into one small subgroups comprised of six taxa representing four species and a major subgroups comprised of 12 taxa representing six species. The former subgroup comprises Pentatropis spiralis, Solenostemma argel (three samples), Odontanthera radians and Glossonema boveanum. The latter larger subgroup includes the remaining species; Pergularia daemia, P. tomentosa (two samples), Kanahia lanifolia, Gomphocarpus sinaicus (two samples), G. fruticosus and the four samples of Calotropis procera.
DISCUSSION
The relationship between the examined species of the subfamily Asclepiadoideae (Apocynaceae) are discussed in the present work through the numerical analysis of all attributes viz. morphological characters, seed protein electrophoresis and RAPD-PCR analysis (Fig. 4). The six species of tribe Ceropegieae are delimited together as a separate group from another major group that comprises 11 species of tribe Asclepiadeae. The separation of species in the two tribes is in agreement with previous taxonomic classification by Endlicher (1938), Cronquist (1968), Markgraf (1972), Spellman (1977), Takhtajan (1980) and Rosatti (1989) based on the morphological characters. In addition Sundell (1980), Kunz (1995), Swarupanandan et al. (1996) and Endress and Bruyns (2000) recognized these two tribes based on the organization of the androecium, where in tribe Asclepiadeae the pollinia in pollen sacs are horizontally or pendulous while in tribe Ceropegieae are oriented upwardly. Liede and Albers (1994) and Goyder (2006) supported the separation between the two tribes (Asclepiadeae and Ceropegieae) by the characters of gynoecium, particularly the presence or absence of true styles and the sharp constriction between stigma-head and ovaries. Phylogenetic studies based genetic variations have also confirmed the delimitation between the two tribes as observed in this study (Liede, 1996, Fishbein, 2001; Goyder et al., 2007; Mahmood et al., 2010).
The six species of tribe Ceropegieae in all analyses used in this study are delimited into 3 groups. The three species of Caralluma (C. penicillata, C. retrospiciens and C. sinaica) and Huernia lodarensis of subtribe Stapeliinae are grouped together in the most analyses in this study. The relatively high distance between H. lodarensis and the three species of Caralluma as indicated in Fig. 4 do not supports the grouping of H. lodarensis in section Caralluma as was proposed by Audissou (2005), and Ramachandran et al., (2011). Moreover, the three species of genus Caralluma are clustered at a low distance coefficient (about 0.25) indicating close morphological resemblance between the genotype of Caralluma species. The two samples of Leptadenia pyrotechnica were also separated from the other studied species of tribe Ceropegieae in the all analyses. This is congruent with their delimitation in subtribe Leptadeniinae by Endlicher (1938) and Meve and Liede (2002).
The Ceropegia arabica of tribe Ceropegieae is separated from the other species of same tribe in all analyses used in this study. Forster and Bruyns (1992), Kunz (1995) and Swarupanandan et al. (1996) included the genus Ceropegia in the subtribe Stapeliinae of tribe Ceropegieae referring to common characters of latex, lack of anther appendage and mode of attachment of the pollinia to the translator in addition to the stylar characters. Also, by using molecular evidence Liede (1997) and Meve and Liede (2004) pointed to the same direction. As early as, Bentham and Hooker (1876) separated the genus Ceropegia in subtribe Ceropegiinae according to the stem morphological characters. Recently the phylogenetic studies carried by Fishbein (2001), Bensusan (2009), Bartosz et al. (2010) and Kullayiswamy et al. (2012) confirmed the separation of Ceropegia arabica of tribe Ceropegieae from the other species of the same tribe; the present results support this separation.
In tribe Asclepiadeae; the separation of Calotropis procera (4 samples) Gomphocarpus fruticosus (2 samples), G. sinaicus (2 samples), Kanahia laniflora, Pergularia daemia, and P. tomentosa (2 samples) in a separate group is in agreement with their previous delimitation in subtribe Asclepiadinae by Sundell (1980), Li et al. (1995), Swarupanandan et al. (1996) Liede (1997, 1999) and Fishbein (2001) based on the vegetative and corona characters as well as style structure and mode of pollinia attachment to the translator. Otherwise, the grouping of the genus Pergularia in subtribe Asclepiadinae in the present work is congruent with its transfer from the subtribe Cynanchinae to subtribe Asclepiadinae by Liede and Meve (1996) based on the analysis of ITS region of the nuclear genome.
Except for Pentatropis spiralis of subtribe Astephaninae the grouping of Glossonema boveanum, Odonthera radians, and Solenostema argel together is in agreement with their previous delimitation in subtribe Glossonematinae by Liede et al. (2002). The grouping of Pentatropis spiralis (Subtribe Astephaninae) with the species of subtribe Glossonematinae is ascertained with the analyses based on seed protein characters (Fig. 2). However, it contradicts the analyses based on morphological criteria and RAPD-PCR (Fig. 1, 3). Meanwhile only a few morphological similarities exist between P. spiralis and the other four species of the same tribe. All of them are characterized by brochidodromous leaf venation, white colour of both corolla and corona and brown ovate seeds.
Liede et al. (2002) showed the subtribe Glossonematinae of the tribe Asclepiadeae, hitherto composed of Arabian and North African genera Glossonema, Odonthanthera and Solenostemma, not to be monophyletic. Also, he suggested that Glossonema and Odontanthera are closely allied to Pentarrhinum, an African genus of five species belonging to subtribe Cynanchinae based on molecular, karyological and morphological evidences. This is supported by the relationships as expressed in the tree based on RAPD analysis. In the meantime Cynanchum acutum of subtribe Cynanchinae is delimited from all taxa of tribe Asclepiadeae in all analyses. This species is morphologically characterized by often growing from rhizomes, opposite petiolated leaves, gynostegial origin of corona and the fusion of staminal and intrastaminal parts. This delimitation is congregant with previous results based on molecular evidences carried by Sennblad and Bremer (1996) and Liede et al. (2002).
CONCLUSION
Relationships between the studied species of subfamily Asclepiadoideae (Apocynaceae), based on morphological variations and polymorphism in seed protein electrophoretic profile and RAPD fingerprinting support the delimitation of the examined species in the two tribes Asclepiadeae and Ceropegieae. Moreover, the present study supports the transfer of the genus Ceropegia from subtribe Stapeliinae to subtribe Ceropegiinae and does not support the grouping of Huernia lodarensis in section Caralluma. Although, the present study separates the two samples of Cynanchum acutum from all taxa of tribe Asclepiadeae, the results are in agreement with the delimitation of the remaining studied species of tribe Ceropegieae in two subtribes Stapeliinae and Leptadeniinae.
ACKNOWLEDGMENT
Thanks are due to Dr. Abdelfattah Badr, Professor of Genetics and Biosystematics, Botany Department, Faculty of Science, Tanta University for the final revision of the present Study.
REFERENCES
- The Angiosperm Phylogeny Group, 2003. An update of the angiosperm phylogeny group classification for the orders and families of flowering plants: APG II. Bot. J. Linn. Soc., 141: 399-436.
CrossRefDirect Link - Badr, A., 1995. Electrophoretic studies of seed proteins in relation to chromosomal criteria and the relationships of some taxa of Trifolium. Taxon, 44: 183-191.
Direct Link - Bartosz, J.P., P. Swiatek and G. Szymczak, 2010. Can a stench be beautiful? - Osmophores in stem-succulent stapeliads (Apocynaceae-Asclepiadoideae-Ceropegieae-Stapeliinae). Flora-Morphol. Distribution Functional Ecol. Plants, 205: 101-105.
CrossRef - Bensusan, K., 2009. Taxonomy and conservation status of Morocan stapeliads (apocynaceae-asclepiadaceae-ceropegieae-stapeliinae). Bulletin de'l Institute Scientifique, Rabat, Section Sciences de la Vie, 31: 67-77.
Direct Link - Bergner, I. and U. Jensen, 1989. Phytoserological contribution to the systematic placement of the Typhales. Nordic J. Bot., 8: 447-456.
CrossRef - Civeyrel, L., A. Le Thomas, K. Ferguson and M.W. Chase, 1998. Critical reexamination of palynological characters used to delimit Asclepiadaceae in comparison to molecular phylogeny obtained from plastid mark sequences. Mol. Phylog. Evol., 9: 517-527.
CrossRef - Collenette, S., 1999. Wild Flowers of Saudi Arabia. 1st Edn. National Commission for Wildlife Conservation and Development (NCWCD), Riyadh, Pages: 799.
Direct Link - Dice, L.R., 1945. Measures of the amount of ecologic association between species. Ecology, 26: 297-302.
CrossRefDirect Link - Endress, M.E. and P.V. Bruyns, 2000. A revised classification of the Apocynaceae s.l. Bot. Rev., 66: 1-56.
Direct Link - Fishbein, M., 2001. Evolutionary innovation and diversification in the flowers of asclepiadaceae. Anal. Missouri Botanical Gardens, 88: 603-623.
Direct Link - Forster, P.I. and P.V. Bruyns, 1992. Clarification of synonymy for the common moth-vine Araujia sericifera (asclepiadaceae). Taxon, 41: 746-749.
Direct Link - Goyder, D., A. Nicholas and S.L. Schumann, 2007. Phylogenetic relationships in subtribe Asclepiadinae (Asclepiadoideae). Annals Missouri Bot. Garden, 94: 423-434.
Direct Link - Jiu-Xiang, H., Z. Shou-zhou, Z. Xue-ying and L. Bing-Tao, 2007. RAPD analysis of twelve species of Apocynaceae. J. Fujian Forestry Sci. Technol.
Direct Link - Khanuja, S.P.S., A.K. Shasany, M.P. Darokar and S. Kumar, 1999. Rapid isolation of DNA from dry and fresh samples of plants producing large amounts of secondary metabolites and essential oils. Plant Mol. Biol., Rep., 17: 1-7.
Direct Link - Ladizinsky, G. and T. Hymowitz, 1979. Seed protein electrophoresis in taxonomic and evolutionary studies. Theor. Applied Genet., 54: 145-151.
CrossRefDirect Link - Laemmli, U.K., 1970. Cleavage of structure protein during the assamble of the head of bacteriophage T4. Nature, 227: 680-685.
Direct Link - Liede, S., 1999. Subtribes and genera of Asclepiadaceae: Reply to Bruyns's response. Taxon, 48: 27-29.
Direct Link - Liede, S. and F. Albers, 1994. Tribal disposition of genera in the Asclepiadaceae. Taxon, 43: 201-231.
Direct Link - Livshultz, T., 2010. The phylogenetic position of milkweeds (Apocynaceae) subfamilies Secamonoideae and Asclepiadoideae): Evidence from the nucleus and chloroplast. Taxon, 59: 1016-1030.
Direct Link - Mahmood, T., S. Muhammad and Z. Shinwari, 2010. Molecular and morphological characterization of Caralluma species. Pak. J. Bot., 42: 1163-1171.
Direct Link - Meve, U. and S. Liede, 2002. A molecular phylogeny and generic rearrangement of the Stapelioid Ceropegieae (Apocynaceae-Asclepiadaceae). Plant Syst. Evol., 234: 171-209.
CrossRefDirect Link - Michiels, A., W. van den Ende, M. Tucker, L. van Riet and A. van Laere, 2003. Extraction of high quality genomic DNA from Latex containing plants. Anal. Biochem., 315: 85-89.
PubMed - Moslem, M.A., 1999. Utilization of milk powder as a source of carbon and nitrogen by soil Mycoflora of Saudi Arabia. Pak. J. Biol. Sci., 2: 674-678.
CrossRefDirect Link - Nilsson, S., M.E. Endress and E. Grafstrom, 1993. On the relationship of the apocynaceae and periplocaceae. Grana, 2: 3-20.
CrossRefDirect Link - Ramachandran, V.S., S. Joseph, H.A. John and C. Sofiya, 2011. Caralluma bicolor sp. nov. (Apocynaceae, Asclepiadoideae) from India. Nordic J. Bot., 29: 447-450.
CrossRef - Rosatti, T.J., 1989. The genera of suborder Apocynaceae (Apocynaceae and Asclepiadaceae) in the Southeastern United States. J. Arnold Arbor., 70: 307-514.
Direct Link - Sennblad, B. and B. Bremer, 1996. The familial and subfamilial relationships of Apocynaceae and Asclepiadaceae evaluated with rbcl data. Plant Syst. Evol., 202: 153-175.
Direct Link - Sennblad, B., M.E. Endress and B. Bremer, 1998. Morphology and molecular data in phylogenetic fraternity: The tribe wrighteae (Apocynaceae) revised. Am. J. Bot., 85: 1143-1158.
PubMed - Sharawy, S.M., 2008. Polymorphism in seed protein electrophoretic pattern and species relationships in the genus Orobanche L. Aust. J. Basic. Appl. Sci., 2: 1298-1306.
Direct Link - Sharawy, S.M. and A. Badr, 2008. Systematic revision of Erodium species in Egypt as reflected by variation in morphological characters and seed protein electrophoretic profiles. Int. J. Bot., 4: 225-230.
CrossRefDirect Link - Sokal, R.R. and C.D. Michener, 1958. A statistical method for evaluating systematic relationships. Univ. Kansas Sci. Bull., 38: 1409-1438.
Direct Link - Spellman, D., 1977. Four species of asclepiadaceae new to panama. Annals Missouri Bot. Garden, 64: 129-132.
Direct Link - Struwe, L., V.A. Albert and B. Bremer, 1994. Cladistics and family level classification of the Gentiales. Cladistics, 10: 175-206.
Direct Link - Sundell, E., 1980. The subfamilial, tribal and subtribal nomenclature of the Asclepiadaceae. Taxon, 29: 257-265.
Direct Link - Swarupanandan, F.T., K.M. Jose, T.K. Sonny, K. Kishorekumar and S.B. Chand, 1996. The subfamilial and tribal classification of the family Asclepiadaceae. Bot. J. Linn. Soc., 120: 327-369.
CrossRefDirect Link - Kullayiswamy, K.R., s. Sandhyarani and S. Karuppusamy, 2012. Ceropegia pullaiahii sp. nov. (Apocynaceae, Asclepiadoideae) from India. Nordic J. Bot.
CrossRef