
Kanlaya Kong-ngern
Department of Biology, Faculty of Science, Khon Kaen University, Khon Kaen 40002, Thailand
Sumontip Bunnag
Department of Biology, Faculty of Science, Khon Kaen University, Khon Kaen 40002, Thailand
Piyada Theerakulpisut
Department of Biology, Faculty of Science, Khon Kaen University, Khon Kaen 40002, Thailand
International Journal of Botany
Year: 2012 | Volume: 8 | Issue: 2 | Page No.: 54-65







ABSTRACT
In order to compare differential growth, physiological and biochemical responses of rice cultivars differing in salt tolerance, four rice cultivars; tolerant Pokkali (PK), moderately tolerant Luang Anan (LA) and two sensitive cultivars Khao Dawk Mali 105 (KDML 105) and Pathumthani 60 (PT 60) were grown in hydroponic culture for 2 weeks and then subjected to NaCl stress at 6 or 12 dS m-1 for thirteen days. The tolerant cultivar (PK) exhibited clearly different pattern of responses from the other cultivars in most of the examined aspects including lower growth inhibition, lower inhibition of net photosynthetic rate, lower Na+/K+ ratio in the shoots, less proline accumulation in the shoots, less membrane damage, lower hydrogen peroxide production, higher catalase and lower guaiacol peroxidase activities. Other parameters including Na+/K+ ratio in roots, proline accumulation in roots and osmotic potential in both roots and shoots did not show appropriate association with the level of salt tolerance. The reduction in growth resulted from drastic reduction in net photosynthetic rate which was observed as early as 24 h after exposure to stress and continued to decrease dramatically. Growth retardation was also related to salt-induced accumulation of hydrogen peroxide which caused lipid peroxidation and membrane leakage. Higher catalase activity in the tolerant cultivar played an important role in combating the hydrogen peroxide, leading to less damage and higher tolerance. The activity of peroxidase was lower in the tolerant cultivar than in the sensitive ones. The observed negative relationship between the amount of proline accumulation and the level of salt tolerance did not support the widely advocated role of proline as an osmoprotectant under salt stress. Further investigation needs to be done to determine the role of this compatible osmolyte in salinity stress response in rice. For practical purpose, those parameters which showed differential pattern of responses including lower Na+/K+ ratio in the shoots, less proline accumulation in the shoots, lower reduction in net photosynthesis rate, lower electrolyte leakage, lower hydrogen peroxide accumulation, higher catalase and lower guaiacol peroxidase activities may be regarded as potential biochemical indicators for selection of salt tolerant rice and targets for improvement through transgenic approaches.
PDF Abstract XML References Citation
Received: December 13, 2011;
Accepted: March 29, 2012;
Published: July 11, 2012
How to cite this article
Kanlaya Kong-ngern, Sumontip Bunnag and Piyada Theerakulpisut, 2012. Proline, Hydrogen Peroxide, Membrane Stability and Antioxidant Enzyme Activity as Potential Indicators for Salt Tolerance in Rice (Oryza sativa L.). International Journal of Botany, 8: 54-65.
DOI: 10.3923/ijb.2012.54.65
URL: https://scialert.net/abstract/?doi=ijb.2012.54.65
DOI: 10.3923/ijb.2012.54.65
URL: https://scialert.net/abstract/?doi=ijb.2012.54.65
INTRODUCTION
Soil salinity is one of the most important abiotic stress factors adversely affecting crop production. In order to develop more salt-tolerant plants using conventional method, molecular breeding or transgenic approaches, it is essential to understand what mechanisms make one cultivar more salt-tolerant than another. The deleterious effects of salinity on plant growth are associated with (1) low osmotic potential of soil, (2) ion imbalance, (3) specific ion effects and (4) accumulation of reactive oxygen species (Ashraf and Harris, 2004). In rice, several authors have shown that Na-K selectivity of plant roots to minimize the entry of Na+ into plants and maintain effective K+ uptake together with the mechanism of low salt transport to expanding leaves is a very important mechanism directly correlated with salt tolerance (Flowers et al., 1991; Munns, 2002). Osmotic adjustment by an accumulation and compartmentation of inorganic ions and organic compatible solutes is another well-characterized mechanism relating to salt tolerance (Flowers and Yeo, 1986; Munns, 2002). Salinity also has an inhibitory effect on chlorophyll synthesis resulting in a reduction in chlorophyll and the severity of the effect vary depending on concentration of salts and duration of stress. Rice plants treated with low salt concentration (<50 mM) for short duration usually have higher chlorophyll content than the non-stressed plants. When the concentration of salt or the duration of treatment increased the chlorophyll content decreased and the extent of the reduction was higher in salt-sensitive varieties (Faustino et al., 1996; Misra et al., 1997). Salt tolerance has also been related to lipid peroxidation and membrane damage (Azooz, 2009). Plants with less salt tolerance ability tended to accumulate more Reactive Oxygen Species (ROS) causing more intense lipid peroxidation which leads to enhanced loss of membrane integrity (Dionisio-Sese and Tobita, 1998; Sairam and Srivastava, 2002; Joseph and Jini, 2010).
Various studies on plant stress responses have seen some contradictory or inconclusive results regarding the differential responses in certain physiological parameters between salt-tolerant and salt-sensitive cultivars. The magnitude of proline accumulation was either positively (Hare et al., 1999; Hien et al., 2003; Demiral and Turkan, 2006) or inversely related (Lutts et al., 1996; Lutts et al.,1999) to salt tolerance. Overexpression of genes leading to more proline synthesis in transgenic tobacco (Kishor et al., 1995) and rice (Zhu et al., 1998) resulted in an enhancement of salt tolerance. In contrast, some authors suggested that intensive accumulation of proline was a symptom of salt-stress injury and a consequence of proteolysis and senescence (Lutts et al., 1999). Maggio et al. (2002) suggested that using transgenic approaches to overexpress proline-synthesising enzymes and hence increased proline concentration in some cases resulted in plants with reduced growth in the absence of stress. In response to salt stress, the activities of superoxide dismutase (SOD) and peroxidase (POX) in the sensitive rice cultivars (Hitomebore and IR 28) changed in a different direction from those of the tolerant cv. PK. However, the patterns of changes in the activities of both enzymes were similar to those of another tolerant cv. Bankat (Dionisio-Sese and Tobita, 1998).
The objective of this study was to compare growth and physiological responses, among four rice cultivars differing in salt tolerance ability in order to gain more understanding of the mechanisms of salt tolerance at the cellular levels which in turn determined growth and yield. The parameters examined included those relating to photosynthesis, ion and proline accumulation, osmotic potential, membrane damage, the accumulation of hydrogen peroxide and the activity of antioxidative enzymes. The parameters which showed highly different pattern of responses to salinity between the salt-tolerant and salt-sensitive cultivars are expected to be useful as potential targets for improvement of salt-tolerant rice cultivars by conventional and transgenic approaches.
MATERIALS AND METHODS
Plant materials and salinity treatments: Seeds of rice (Oryza sativa L. cvs. Pokkali; PK, Luang Anan; LA, Khao Dawk Mali 105; KDML 105 and Pathumthani 60; PT 60) were kindly provided by Pathumthani Rice Research Institute, Thailand. PK is a tall indica variety from India, well-known for its tolerance to salinity. LA is a local Thai variety classified as moderately salt tolerant (Kanawapee et al., 2011). KDML 105 and PT 60 are commercial cultivars widely grown in Thailand and are known to be salt sensitive. Seeds were surfaced sterilized, imbibed for 48 h and then sown on plastic grids placed above 4 L black plastic pots containing distilled water. When seedlings were 5 days old, distilled water was replaced with nutrient solution (Yoshida et al., 1976). When the plants were 14 days old, the nutrient solutions were replaced by the ones containing NaCl at the salinity level of 0, 6 or 12 dS m-1. The plants were grown in the salinized solutions for thirteen days, during which the pH of the solutions were adjusted daily to 5.0 and renewed every five days. All growth and physiology measurements were determined after the plants were grown in the salinized solutions for thirteen days, except for the measurement of photosynthetic rates which was performed on days 1, 6 and 12 after the beginning of salinity treatment.
Measurement of growth and net photosynthesis rate: Thirteen days after salinization, five randomly sampled seedlings of each treatment were harvested and separated into shoots and roots. Fresh weight of each sample was recorded, then the samples were oven-dried at 60°C for 72 h for determination of dry weight. Growth Rate (GR) was determined following the method outlined by Dionisio-Sese and Tobita (1998). GR was calculated from the increase in dry weight of plants at the beginning and the end of salt treatment, using the equation:
where, DW is the plant dry weight, t is the time and subscripts denote initial and final sampling, that is, 0 and 13 days after salinity treatment. Net photosynthesis rate (μmol CO2 m-1 sec-1) of the second leaf from the top of each plant (five plants for each treatment) was measured between 900-1500 using portable photosynthesis system (Li-cor 6400).
Determination of proline, Na+ and K+ content and osmotic potential: Free proline content of shoots and roots was determined using 0.1 g sample using acid-ninhydrin method based on Bates et al. (1973). For measurement of Na+ and K+ content, shoots and roots were dried at 70°C for 48 h, accurately weighed, digested with perchloric acid and nitric acid (1:2) and the volume of extracts were adjusted to 100 mL. The extracts were diluted to a ratio of 1:100 and Na+ and K+ content determined by atomic absorption spectrophotometer (Model GBC 932 AA). For osmotic potentials, shoot and root samples (1.0 g) were ground with small tissue grinder in microcentrifuge tubes and centrifuged at 13,000 g for 15 min. The osmotic concentration of the supernatant was determined in an osmometer (KNAUER automatic semi-microosmometer A0300 version 0291). The osmotic potential of the extract was determined according to van’t Hoff’s equation (Salisbury and Ross, 1992). Five replicates were measured for each treatment.
Determination of electrolyte leakage, lipid peroxidation and hydrogen peroxide: Electrolyte Leakage (EL) test of leaf samples was determined by comparing the initial Electrical Conductivity (EC) of the medium bathing the fresh plant materials at 32°C for 2 h (EC1) with the final EC (EC2) measured after the plant materials had been killed by autoclaving at 121°C for 20 min to release all electrolytes. The electrolyte leakage was expressed following the formula EL = EC1/EC2x100 (Dionisio-Sese and Tobita, 1998). Lipid peroxidation was determined by measuring the amount of malondialdehyde (MDA) in leaf tissues using thiobarbituric acid method described by Stewart and Bewley (1980). Hydrogen peroxide content was determined according to Sergiev et al. (1997). Hydrogen peroxide was extracted from leaf tissues (0.5 g) on an ice bath with 5 mL of 0.1% (w/v) trichloroacetic acid (TCA). Crude extract was centrifuged at 12,000 g for 15 min and 0.5 mL of the supernatant was transferred to a 15 mL test tube. The supernatant was added with 0.5 mL of 10 mM potassium phosphate buffer (pH 7.0), 1 mL of 1 M potassium iodide (KI) and mixed by vortexing briefly. The absorbance of mixture was measured at 390 nm. A mixture without the supernatant served as a blank. Each parameter was evaluated using five plants for each treatment.
Catalase and peroxidase enzymes assay: For enzyme assays and estimation of total protein, 0.5 g of frozen leaf tissue (from five plants for each treatment) was ground to a fine powder with liquid nitrogen and extracted with 400 μL of ice-cold extraction buffer [50 mM phosphate buffer (pH 7.0)]. The extracts were centrifuged at 4°C for 30 min at 10,000 g and the resulting supernatants were used as the crude extracts. Soluble protein content of the crude extract was determined using the Bio-Rad assay kit (Bio-Rad Laboratories, USA) with Bovine Serum Albumin (BSA) as a calibration standard. The activity of Peroxidase (POX) was determined using the guaiacol oxidation method (Chance and Maehly, 1955) in a 3 mL reaction mixture containing 10 mM phosphate buffer (pH 6.4), 8 mM guaiacol, 100-200 μL crude extract and 2.75 mM H2O2. The increase in absorbance was recorded at 470 nm within 30 sec (linear phase) after H2O2 was added. One unit of POX activity was expressed as the change in absorbance per minute and specific activity as ΔA470 min-1 mg-1 protein. The activity of catalase was assayed by measuring the initial rate of disappearance of H2O2 by the method of Kato and Shimizu (1987). Three mL of catalase reaction mixture contained 10 mM potassium phosphate buffer, pH 7.0, 0.1 mL enzyme extract and 0.035 mL 3% H2O2. The decrease in H2O2 concentration was followed as the decline in optical density at 240 nm and the activity was calculated using the extinction coefficient of 40 mM-1 cm-1 for H2O2. The specific activity was expressed as units mg-1 protein min-1.
Statistical analysis: The experimental design was completely randomized with five replicates per treatment. Statistical analysis was performed using one-way ANOVA and the difference between the mean values was compared using Duncan’s Multiple Range Test (DMRT) at the p<0.01 level.
RESULTS
Effect of NaCl on root and shoot growth: The effects of NaCl stress on growth of the tolerant (PK) and moderately tolerant (LA) cultivars were notably different (Fig. 1a, b) from those on the sensitive ones (KDML 105 and PT 60). Dry weights of roots and shoots of PK and LA seedlings treated with low (6 dS m-1) and high (12 dS m-1) levels of salinity were not significantly different from those of control seedlings, whereas those of the stressed KDML 105 and PT 60 were significantly decreased compared to the controls. At 12 dS m-1 salinity, PK showed the lowest percentage of reduction in root and shoot dry weight (29.63 and 23.73%) while KDML 105 the highest (62.50 and 53.93%). The highest level of tolerance of PK was also revealed by the insignificant difference in GR between both stressed groups compared with the control (Fig. 1c). At 12 dS m-1 salinity, the reduction in GR rates of PK, LA, KDML 105 and PT 60 were 39.76, 51.72, 80.21 and 66.27%, respectively.
Effect of NaCl on net photosynthetic rate: The effects of NaCl on net photosynthetic rate of the second leaf from the top of each rice seedling at days 1, 6 and 12 after the introduction of NaCl are shown in Fig. 2. The concentration of NaCl as well as the stress durations led to significant inhibition of net photosynthetic rates in all cultivars tested. The effect was remarkable when higher concentration was confounded with increasing stress duration in all cultivars tested. At low salinity level and all stress durations, salinity caused a significant decrease in the net photosynthetic rate of all cultivars.
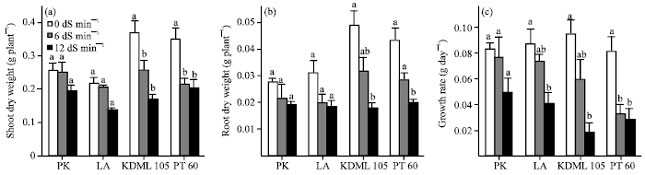 | |
| Fig. 1(a-c): | Effect of salinity on dry weight (g/plant) of (a) Shoots, (b) Roots and (c) Growth rate (g/day) of 27-day-old seedlings of four rice cultivars which were subjected to 0, 6 and 12 dS m-1 salinity for 13 days. Values represent the Mean±SD, For each cultivar, means with different letters are significantly different at p<0.01 |
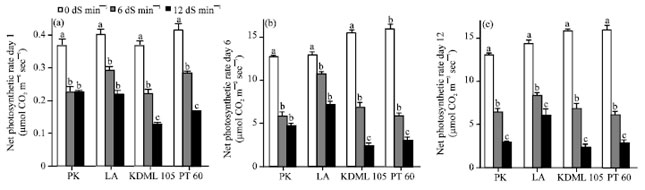 | |
| Fig. 2(a-c): | Effect of salinity on the net photosynthetic rate (μmol CO2 m-2 sec-1) of the second leaf from top of seedlings of four rice cultivars which were subjected to 0, 6 and 12 dS m-1 salinity levels for (a) 1 day, (b) 6 days and (c) 12 days, Values represent the Mean±SD. For each cultivar, means with different letters are significantly different at p<0.01 |
Under high salinity level at days 1 and 6, net photosynthetic rates were more severely affected in the salt-sensitive cultivars than in the salt-tolerant ones. However, under high salinity treatment at day 12, salinity caused a strong inhibition in all cultivars tested. The net photosynthetic rates under high salinity treatment at day 12 were reduced by 4.51 and 2.35 folds in the salt-tolerant PK and LA, respectively whereas, in the salt-sensitive KDML 105 and PT 60, they were reduced by 5.31 and 5.66 folds, respectively.
Effect of NaCl on ion and proline content: With increasing salinity, the concentration of Na+ in shoots and roots in all cultivars was dramatically increased, whereas that of K+ decreased (Fig. 3). There were strong relationships between Na+ and K+ concentration and the Na+/K+ ratio in the shoots with the level of salt tolerance. The shoots of more tolerant cultivars had lower amount of Na+ but higher amount of K+ than the less tolerant ones. The results indicated that the Na+/K+ ratio in shoots is strongly related with the degree of salt tolerance (Fig. 3c). However, no relationship was found between the degree of salt tolerance and the Na+/K+ ratio in roots (Fig. 3f). The presence of NaCl resulted in an increase in root proline in all cultivars (Fig. 4a). The salt-sensitive KDML 105 accumulated the highest amount of root proline followed by the salt-sensitive PT 60, the moderately tolerant LA and the tolerant PK. The root proline content in PK, LA, KDML 105 and PT 60 treated with 12 dS m-1 NaCl showed 35.54, 8.97, 32.60 and 41.42% increase, respectively compared with the control plants. Therefore, the amount of root proline in LA was the least and PT 60 the most affected by salt stress. The constitutive amounts of proline in shoots of LA, KDML 105 and PT 60 under non-stressed condition were similar to those found in the roots, except for PK in which the amount of shoot proline was double that in the roots. The accumulation of proline due to salinity stress was much more dramatic in shoots than in roots (Fig. 4b). In response salinity at 12 dS m-1, shoot proline increased 2.7, 11.0, 6.8 and 7.9 folds in PK, LA, KDML 105 and PT 60, respectively.
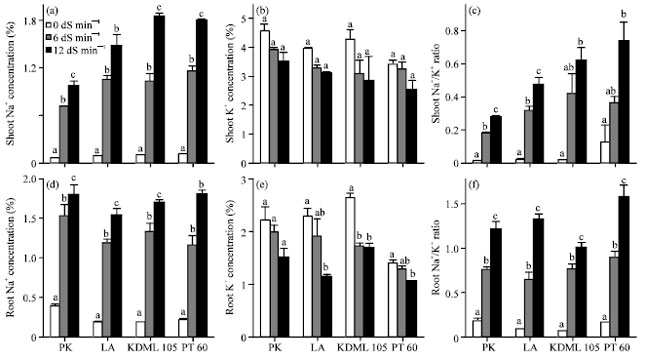 | |
| Fig. 3(a-f): | Effect of salinity on the amount of (a) Na+, (b) K+, (c) Na+/K+ ratio in the shoots and the amount of, (d) Na+, (e) K+ and (f) Na+/K+ ratio in the roots of 27-day-old seedlings of four rice cultivars which were subjected to 0, 6 and 12 dS m-1 salinity levels, The amount of Na+ and K+ in the shoot were determined after 13 days of salinity treatment, Values represent the Mean±SD. For each cultivar, means with different letters are significantly different at p<0.01 |
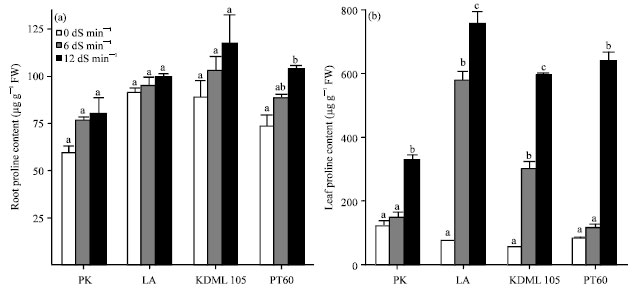 | |
| Fig. 4(a-b): | Effect of salinity on proline content (μg g-1 FW) in the (a) Roots and (b) Leaves of 27-day-old seedlings of four rice cultivars which were subjected to 0, 6 and 12 dS m-1 salinity levels. Leaf and root proline contents were determined after 13 days of treatment, Values represent the Mean±SD, For each cultivar, means with different letters are significantly different at p<0.01 |
Effect of NaCl on osmotic potential in roots and shoots: The osmotic potential of both shoots and roots became more negative with increasing salinity levels. Salinity appeared to cause more drastic change in osmotic potential of shoots than of roots. Salinity level at 12 dS m-1 caused significant decreases in the root osmotic potential of all cultivars (Fig. 5a). The lowest root osmotic potentials were observed in PK and KDML 105. Both levels of salinity caused marked decreases in shoot osmotic potential in all cultivars (Fig. 5b). The reduction in leaf osmotic potential of PK, LA, KDML 105 and PT 60 was 161.05, 220.72, 313.92 and 95.14%, respectively compared with the controlled plants.
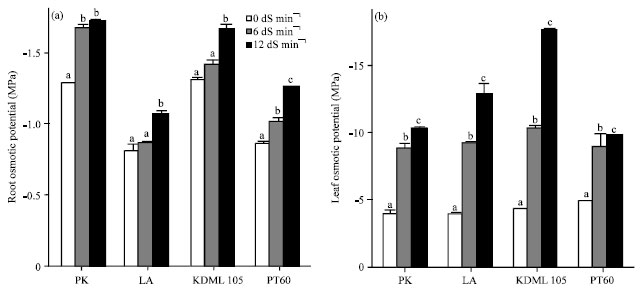 | |
| Fig. 5(a-b): | Effect of salinity on osmotic potential (MPa) in the (a) Roots and (b) Leaves of 27-day-old seedlings of four rice cultivars which were subjected to 0, 6 and 12 dS m-1 salinity levels. Leaf and root osmotic potentials were determined after 13 days of treatment, Values represent Mean±SD, Values represent the Mean±SD, For each cultivar, means with different letters are significantly different at p<0.01 |
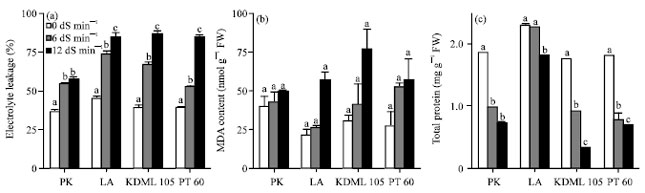 | |
| Fig. 6(a-c): | Effect of salinity on (a) Electrolyte leakage rate (%), (b) MDA content (mmol mg-1 protein) and (c) Total protein content (mg g-1 protein) in the leaves of 27-day-old seedlings of four rice cultivars which were subjected to 0, 6 and 12 dS m-1 salinity levels, Electrolyte leakage, total protein content and MDA content were determined after 13 days of salinity treatment, Values represent Mean±SD, Values represent the Mean±SD, For each cultivar, means with different letters are significantly different at p<0.01 |
The salt-tolerant PK and LA showed a smaller reduction in the leaf osmotic potential than the salt-sensitive KDML 105. However, the sensitive PT 60 showed lowest reduction in the leaf osmotic potential. Therefore, no exact relationship could be drawn between root and leaf osmotic potential and the degree of salt tolerance.
Effect of NaCl on electrolyte leakage, lipid peroxidation and total protein: The effect of NaCl on membrane integrity was monitored by means of EL test in fresh plant samples. The amount of electrolytes leaked out of the cells was assessed indirectly by conductometric measurements. Under controlled conditions, EL of all cultivars tested did not vary greatly i.e. between 37.28% in PK and 45.71% in LA (Fig. 6a). When subjected to salinity treatment, EL increased with increasing NaCl concentrations and almost reached 90% in all cultivars tested, except for PK, in which EL increased to only 60%. At both salt concentrations, the amount of EL in PK was lower than in other cultivars, whereas the moderately-tolerant LA showed comparable amount of EL to the salt-sensitive KDML 105 and PT 60. Membrane of the salt-sensitive KDML 105 tended to be the most severely affected by high level of salinity (12 dS m-1) as indicated by 120.69% increment in EL compared with the controlled plants, followed by PT 60 (112.97%) and LA (86.15%).
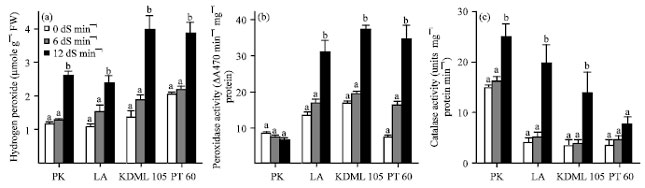 | |
| Fig. 7(a-c): | Effect of salinity on the amount of (a) Hydrogen peroxide concentration (μmol g-1 protein), (b) The activity of peroxidase (ΔA470 min-1 mg-1 protein) and (c) The activity of catalase (units min-1 mg-1 protein) in the leaves of 27-day-old seedlings of four rice cultivars which were subjected to 0, 6 and 12 dS m-1 salinity levels, Hydrogen peroxide, catalase and peroxidase activities were determined after 13 days of salinity treatment. Values represent Mean±SD, Values represent the Mean±SD, For each cultivar, means with different letters are significantly different at p<0.01 |
The results indicated that the increase in EL is associated with the level of salt tolerance. At both NaCl concentrations, MDA content increased with increasing salinity in all cultivars (Fig. 6b). Under non-stressed condition (0 dS m-1), the MDA level in PK was higher than that in other cultivars. However, with increasing concentration of NaCl, PK showed only a small increase in MDA content, whereas the other three cultivars showed larger increases. At 12 dS m-1 salinity level, the increase in MDA content in PK, LA, KDML 105 and PT 60 were 25.38, 159.14, 150.56 and 104.85%, respectively compared with the control plants. A dramatic reduction in leaf total protein content was found in PK, KDML 105 and PT 60 but less reduction was observed in LA (Fig. 6c).
Effect of NaCl on H2O2 content and activities of antioxidative enzymes: The amount of H2O2 content in leaves increased with increasing salinity levels as compared with the control plants (Fig. 7a). In all cultivars, the H2O2 content slightly increased at low salinity treatment, then increased dramatically at high salinity treatment. The increase in the amount of H2O2 in the salt-tolerant cultivars was lower than that in the salt-sensitive ones. In contrast to the other three less tolerant cultivars, the salt-tolerant PK showed a slight decrease in peroxidase activity (Fig. 7b). The peroxidase activity in PK was 8.46 units min-1 mg-1 protein for the controlled plants and dropped to 6.65 units min-1 mg-1 protein (21.37% reduction) for plants treated with NaCl at 12 dS m-1. On the other hand, the pattern of change in peroxidase activity in the moderately tolerant LA was more similar to that of the salt-sensitive KDML 105 and PT 60 than that of the salt-tolerant PK. The increases in peroxidase activity in LA, KDML 105 and PT 60 were 127.97, 121.95 and 353.96%, respectively, as compared with the control plants. Unlike peroxidase, catalase activity increased in response to NaCl in all cultivars with increasing salinity levels (Fig. 7c). The amount of catalase activity in the salt-tolerant PK was much higher than that in other cultivars under non-stressed condition as well as in both NaCl concentrations.
DISCUSSION
The detrimental effects of NaCl on plants are consequences of both water deficit resulting in osmotic stress and the effects of excess sodium ions on critical physiological parameters and biochemical processes. Rice cultivars chosen for this study represented three levels of salt tolerance to compare different pattern of physiological and biochemical responses. According to our preliminary screening test based on the International Rice Research Institute’s protocol (Gregorio et al., 1997), PK was rated as ‘tolerant’, LA ‘moderately tolerant’ and KDML 105 and PT 60 ‘sensitive’. After 13 days in salinized solution, growth of all cultivars, particularly the sensitive ones, was retarded leading to the reduction in fresh and dry weights of shoots and roots with increasing salinity. For short-term exposure, Vaidyanathan et al. (2003) also reported that after the treatment of 250 mM NaCl for 42 h, the salt-tolerant PK showed only 6.0% reduction in fresh weight, whereas the sensitive cv. Pusa Basmati 1 showed a reduction of approximately 17%. Other growth-related parameters, i.e., GR and net photosynthetic rate were also reduced by salt stress and the more sensitive cultivars were more affected than the more tolerant ones.
The reduction in growth was directly related to the inhibitory effects of salinity on leaf photosynthesis and leaf conductance (Myers et al., 1990). The inhibition of net photosynthetic rate of the second leaf from the top increased with increasing salt concentration and duration of stress. A dramatic reduction in the net photosynthetic rate was observed as early as 24 h after the addition of NaCl. The net photosynthetic rates of the more tolerant cultivars tended to be less affected than those of the more sensitive ones. In addition to net photosynthetic rate, other photosynthetic parameters including maximum quantum yield of PSII, non-photochemical quenching and photon yield of PSII were reduced in rice subjected to high salt stress (Amirjani, 2011).
The most well-established mechanism by which plants could survive salt stress is to compartmentalize ions away from the cytosol (Apse et al., 1999). The results showed that the increase of Na+ concentration and the reduction of K+ concentration in rice shoots were related to the level of salt tolerance, while no clear relationship could be observed in the roots. The most tolerant PK showed the lowest shoot Na+ and the highest shoot K+ concentration which resulted in the lowest Na+/K+ ratio in the shoot. In contrast, the salt-sensitive PT 60 had the highest shoot Na+, the lowest shoot K+ concentration and the highest Na+/K+ ratio in the shoot. The moderately tolerant LA exhibited the values in between those of the tolerant PK and the sensitive KDML 105 and PT 60. This data supported the claim that Na+/K+ ratio in the shoot is the most important factor closely associated with the degree of salt tolerance (Flowers et al., 1991; Munns, 1993; Gregorio et al., 1997). The relationship between the degree to which plants tolerate salt stress and their capacity to maintain a high K+/Na+ ratio (or low Na+/K+) has been noted by several authors (Omielan and Dvorak, 1991; Dvorak et al., 1994; Goudarzi and Pakniyat, 2008).
Proline has been widely considered to be a compatible solute that accumulates in plants in response to a wide variety of environmental stresses and confers stress tolerance by contributing to osmotic adjustment, protecting proteins and membranes and quenching reactive oxygen species (Matysik et al., 2002; Heidari, 2009; Mudgal et al., 2010). However, the effective role of proline accumulation on salt tolerance has been very controversial. All four rice cultivars showed an increase in proline levels in both roots and leaves with increasing salinity levels as compared with the controlled plants and the leaf proline content showed a much higher degree of increase than that of the root. Similar finding was observed in six citrus cultivars that proline content in leaves was higher and was more useful than that in roots as a determinant of the level of salt stress experienced by plants (Ghotb Abadi et al., 2010). The most tolerant PK showed the lowest amount of proline in both roots and leaves and the percentages of increase in response to salt were much lower than those in LA and the two salt-sensitive cultivars. Thus, it may be implicated that the high level of salt tolerance in PK did not depend on the amount of root and leaf proline. Theerakulpisut et al. (2005) also found that PK accumulated the lowest amount of leaf and root proline, the most sensitive cultivar (IR 29) the highest and the three moderately tolerant cultivars displayed the moderate values. Similarly, several authors reported that the increase in proline was more pronounced in the sensitive cultivars than in the tolerant rice cultivars (Heuer, 1994; Demiral and Turkan, 2006; Vaidyanathan et al., 2003). In contrast, some authors found that salt-tolerant cultivars showed stronger accumulation of proline than sensitive ones in rice (Lutts et al., 1999; Hien et al., 2003) and barley (Fedina et al., 2002). Therefore, the significance of proline accumulation in salt-tolerant and salt-sensitive rice during stress has yet to be fully understood.
Overproduction of proline in transgenic tobacco (Kishor et al., 1995) and rice (Zhu et al., 1998) enhanced plant biomass and tolerance under water and salinity stress. Several studies, however, have indicated that proline levels did not necessarily correspond to the tolerance of plants for salt and may be induced only after damage has been sustained (Bhaskaran et al., 1985; Chandler and Thorpe, 1987; Moftah and Michel, 1987). In an in vitro study, Sivakumar et al. (1998) reported that proline, at a concentration as low as 100 mM, suppressed activity of Rubisco in seedlings of Brassica juncea, Sesbania sesban and Oryza sativa. Moreover, the cases of negative relationship between proline and salt tolerance have been reported in several studies. Lin and Kao (1996) investigated the effects of NaCl on changes in proline level in rice roots and found that accumulation of proline promoted root growth inhibition caused by NaCl. Garcia et al. (1997) found that by comparing the effects of trehalose and proline on growth and physiology of rice seedlings, proline had no effect or, in some cases, exacerbated the effect of NaCl on growth inhibition and chlorophyll loss whereas trehalose offered much better protection. Moreover, proline and trehalose displayed an opposite effect on the expression of salT, a salt-sensitive marker gene. Proline enhanced, whereas trehalose suppressed the expression of salT which negatively correlated with Na accumulation. Lutts et al. (1999) suggested that proline accumulation in salt-sensitive rice leaf was likely a symptom of salt-stress injury and the result of the increase in the endogenous pool of its precursor glutamate. Recently, Saleethong et al. (2011) also found that salt-sensitive KDML 105 rice cultivar accumulated higher proline than the salt-tolerant PK under salinity stress. Transgenic plants engineered to overproduce proline were found to exhibit impaired growth in the absence of stress (Maggio et al., 2002).
The reduction in osmotic potential is an essential adaptive mechanism in plants growing in saline soil. The osmotic potential of roots and leaves in all cultivars decreased with increasing NaCl concentrations. The percentage of reduction in the osmotic potential in roots tends to be lower than those in leaves. The reduction in leaf osmotic potential in all cultivars (except PT 60) was related to the levels of salt tolerance. However, no exact relationship could be drawn between the levels of salt tolerance and the level of reduction in root osmotic potential. Similar trend was also found in the previous study involving six rice cultivars representing four levels of salt tolerance (Theerakulpisut et al., 2005). In the present study, the most tolerant PK showed the highest (least negative) leaf osmotic potential, whereas the sensitive KDML 105 the lowest (most negative). Higher leaf osmotic potential in the more tolerant rice may indicate that salt tolerance ability in the tolerant cultivars may be more attributed to physiological parameters other than leaf osmotic potential. However, the reduction in osmotic potential may play more important roles in sensitive cultivars and this reduction may result from the accumulation of ions and proline. Many reports have demonstrated that turgor maintenance which is an important mechanism in salt stress tolerance, is obtained by means of increasing cell solute concentration i.e., osmotic adjustment. Proline is known to be an important component of the process. A close relationship between the decrease in the leaf osmotic potential and the accumulation of proline has been reported by Heuer and Nadler (1998). They reported that an accumulation of proline occurred concomitantly with a decline in leaf osmotic potential in potato.
The amount of electrolyte leakage from leaf cells was greatly affected by salinity indicating that salinity leads to an increase in cell membrane permeability. The EL of all cultivars increased with increasing NaCl concentrations. Although under non-stressed condition, EL of leaves did not vary among cultivars. However, in the 12 dS m-1 salinity treatment, most membrane damage occurred in the moderately tolerant LA and the salt-sensitive KDML 105 and PT 60 but less damage was observed in PK. The moderately tolerant LA exhibited similar values of EL to KDML 105 and PT 60. PK also showed the lowest percentage of increment in MDA content, an indicator metabolite of lipid peroxidation reactions. Thus, it may be implicated that lower MDA content and less EL are associated with the salt tolerance in PK. Similar changes in the level of lipid peroxidation and electrolyte leakage in response to NaCl stress have been reported by several authors. Dhindsa et al. (1981) reported that there are similar changes in the level of lipid peroxidation and electrolyte leakage in tobacco leaves. In rice Lutts et al. (1996) reported that MDA content was lowest in salt-tolerant cultivars (Nona Bokra and IR 4630) and highest in salt-sensitive cultivars (I Kong Pao and IR 31785) and MDA content was positively correlated with EL. (Dionisio-Sese and Tobita, 1998) found that EL increased with increasing NaCl concentration in the medium and that it was higher in salt-sensitive than in salt-resistant rice cultivars.
Effects of salinity on the induction of ROS production including H2O2 and the activity of antioxidant enzymes have been studied by several authors. NaCl-induced H2O2 accumulation was non-localized and severely reduced overall plant growth and productivity (Uchida et al., 2002; Vaidyanathan et al., 2003). The result showed that the leaf H2O2 content under salt stress was lower in the more tolerant (PK and LA) than in the more sensitive cultivars (KDML 105 and PT 60). In contrast, the activity of catalase was higher in the more tolerant than the more sensitive cultivars. Both parameters may be regarded as potential indicators for salt tolerance. The action of catalase activity is vital for detoxification of H2O2. This result supported the previous report of Vaidyanathan et al. (2003), who studied the effect of NaCl stress (100-300 mM) on two rice cultivars differing in salt tolerance. They found that the salt-tolerant PK showed higher activity of catalase and lower levels of H2O2 than the salt-sensitive Pusa Basmati 1. The present results also showed that the most tolerant cultivar (PK) which had a lower level of H2O2 also had less MDA content and membrane damage. A similar finding was reported by Lin and Kao (1998) that H2O2 in detached rice leaves is related to membrane damage.
The pattern of change in leaf peroxidase activity in response to salinity differed between PK and the other three cultivars. The activity of peroxidase in PK showed a slight non-significant decrease in both salinity levels whereas peroxidase activity in the other three cultivars exhibited a marked increase especially in high salinity treatment. Similar results were reported in which the salt-tolerant rice cultivar (PK) showed a slight decrease in peroxidase activity in the leaves but the moderately tolerant (Bankat) and the sensitive (Hitomebore and IR 29) ones showed increasing peroxidase activity with increasing salinization (Dionisio-Sese and Tobita, 1998). Peroxidase was known to function in ROS scavenging as well as in the biosynthesis of cell wall materials including lignifications and suberization which leads to cell wall stiffening and restricted cell growth. The correlation between high peroxidase activity and the reduction of growth, observed in sensitive rice varieties in this study, was earlier observed by Lee and Lin (1995) in anoxia-treated rice seedlings.
CONCLUSION
In comparison to the sensitive rice cultivars, higher salt tolerance (less growth reduction) in the tolerant rice was evidently related to lower shoot Na+ concentration, lower inhibition of photosynthesis, lower level of salt-induced H2O2 as a result of higher catalase activity and less lipid peroxidation (low MDA) leading to greater ability to maintain the membrane integrity (lower EL). Proline accumulation did not play a major role in combating salinity stress since the tolerant cultivar accumulated relatively small amount as compared to the sensitive cultivars in which high concentration of proline did not seem to offer much protection. It is concluded that low shoot Na+/K+ ratio, low shoot proline content, lower reduction of net photosynthesis rate, low H2O2 accumulation, high catalase and low peroxidase activity are potentially useful biochemical indicators for selection of salt tolerant rice cultivars.
ACKNOWLEDGMENTS
The authors are grateful to Khon Kaen University for a research grant to the Research Group for Genomics and Proteomics for Improvement of Salt-tolerant Rice and the Higher Education Commission for financial support to KK. The generous gifts of rice seeds from Pathumthani Rice Research Institute, Thailand is also gratefully acknowledged.
REFERENCES
- Amirjani, M.R., 2011. Effect of salinity stress on growth, sugar content, pigments and enzyme activity of rice. Int. J. Bot., 7: 73-81.
CrossRefDirect Link - Apse, M.P., G.S. Aharon, W.A. Snedden and E. Blumwald, 1999. Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science, 285: 1256-1258.
PubMed - Ashraf, M. and P.J.C. Harris, 2004. Potential biochemical indicators of salinity tolerance in plants. Plant Sci., 166: 3-16.
CrossRefDirect Link - Azooz, M.M., 2009. Foliar application with riboflavin (Vitamin B2) enhancing the resistance of Hibiscus sabdariffa L. (deep red sepals variety) to salinity stress. J. Biol. Sci., 9: 109-118.
CrossRefDirect Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Bhaskaran, S., R.H. Smith and R.J. Newton, 1985. Physiological changes in cultured sorghum cells in response to induced water stress: I. free proline. Plant Physiol., 79: 266-269.
Direct Link - Chance, B. and A.C. Maehly, 1955. Assay of catalases and peroxidases. Methods Enzymol., 2: 764-775.
CrossRefDirect Link - Chandler, S.F. and T.A. Thorpe, 1987. Proline accumulation and sodium sulphate tolerance in callus cultures of Brassica napus L. cv westar. Plant Cell Rep., 6: 176-179.
Direct Link - Demiral, T. and I. Turkan, 2006. Exogenous glycinebetaine affects growth and proline accumulation and retards senescence in two rice cultivars under NaCl stress. Environ. Exp. Bot., 56: 72-79.
CrossRefDirect Link - Dhindsa, R.S., P. Plumb-Dhindsa and T.A. Thorpe, 1981. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation and decreased levels of superoxide dismutase and catalase. J. Exp. Bot., 32: 93-101.
CrossRefDirect Link - Dionisio-Sese, M.L. and S. Tobita, 1998. Antioxidant responses of rice seedlings to salinity stress. Plant Sci., 135: 1-9.
CrossRefDirect Link - Dvorak, J., M.M. Noaman, S. Goyal and J. Gorham, 1994. Enhancement of the salt tolerance of Triticum turgidum L. by the Kna1 locus transferred from the Triticum aestivum L. chromosome 4D by homoeologous recombination. Theor. Applied Genet., 87: 872-877.
CrossRefDirect Link - Faustino, F.C., H.S. Lips and E.P. Pacardo, 1996. Physiological and biochemical mechanisms of salt tolerance in rice: 1: Sensitivity thresholds to salinity of some physiological processes in rice (Oryza sativa L.). Philip. J. Crop Sci., 21: 40-50.
Direct Link - Flowers, T.J. and A.R. Yeo, 1986. Ion relations of plants under drought and salinity. Aust. J. Plant Physiol., 13: 75-91.
CrossRefDirect Link - Garcia, A.B., J.A. Engler, S. Iyer, T. Gerats, M. van Montagu and A.B. Caplan, 1997. Effects of osmoprotectants upon NaCl stress in rice. Plant Physiol., 115: 159-169.
Direct Link - Abadi, F.S.G., M. Mostafavi, A. Eboutalebi, S. Samavat and A. Ebadi, 2010. Biomass accumulation and proline content of six citrus rootstocks as influenced by long-term salinity. Res. J. Environ. Sci., 4: 158-165.
CrossRefDirect Link - Goudarzi, M. and H. Pakniyat, 2008. Comparison between salt tolerance of various cultivars of wheat and maize. J. Applied Sci., 8: 2300-2305.
CrossRefDirect Link - Hare, P.D., W.A. Cress and J. van Staden, 1999. Proline synthesis and degradation: A model system for elucidating stress-related signal transduction. J. Exp. Bot., 50: 413-434.
CrossRefDirect Link - Heidari, M., 2009. Antioxidant activity and osmolyte concentration of sorghum (Sorghum bicolor) and wheat (Triticum aestivum) genotypes under salinity stress. Asian J. Plant Sci., 8: 240-244.
CrossRefDirect Link - Heuer, B. and A. Nadler, 1998. Physiological response of potato plants to soil salinity and water deficit. Plant Sci., 137: 43-51.
CrossRefDirect Link - Hien, D.T., M. Jacobs, G. Angenon, C. Hermans, T.T. Thu, L. van Son and N.H. Roosens, 2003. Proline accumulation and Δ1-pyrroline-5-carboxylate synthetase gene properties in three rice cultivars differing in salinity and drought tolerance. Plant Sci., 165: 1059-1068.
Direct Link - Joseph, B. and D. Jini, 2010. Insight into the role of antioxidant enzymes for salt tolerance in plants. Int. J. Bot., 6: 456-464.
CrossRefDirect Link - Theerakulpisut, P., N. Kanawapee, J. Sanitchon and P. Srihaban, 2011. Genetic diversity analysis of rice cultivars (Oryza sativa L.) differing in salinity tolerance based on RAPD and SSR markers. Electron. J. Biotechnol., Vol. 14.
CrossRefDirect Link - Kato, M. and S. Shimizu, 1987. Chlorophyll metabolism in higher plants. VII. Chlorophyll degradation in senescing tobacco leaves; phenolic-dependent peroxidative degradation. Can. J. Bot., 65: 729-735.
CrossRefDirect Link - Kishor, P.B.K., Z. Hong, G.H. Miao, C.A.A. Hu and D.P.S. Verma, 1995. Overexpression of δ-pyrroline-5-carboxylate synthetase increases proline production and confers osmotolerance in transgenic plants. Plant Physiol., 108: 1387-1394.
Direct Link - Lee, T.M. and Y.H. Lin, 1995. Changes in soluble and cell wall-bound peroxidase activities with growth in anoxia-treated rice (Oryza sativa L.) coleoptiles and roots. Plant Sci., 106: 1-7.
CrossRefDirect Link - Lin, C.C. and C.H. Kao, 1996. Proline accumulation is associated with inhibition of rice seedling root growth caused by NaCl. Plant Sci., 114: 121-128.
Direct Link - Lin, J.N. and C.H. Kao, 1998. Effect of oxidative stress caused by hydrogen peroxide on senescence of rice leaves. Bot. Bull. Acad. Sin., 39: 161-165.
Direct Link - Lutts, S., J.M. Kinet and J. Bouharmont, 1996. NaCl-induced senescence in leaves of rice (Oryza sativa L.) cultivars differing in salinity resistance. Ann. Bot. 78: 389-398.
CrossRefDirect Link - Lutts, S., V. Majerus and J.M. Kinet, 1999. NaCl effects on proline metabolism in rice (Oryza sativa L.) seedlings. Physiol. Plant., 105: 450-458.
CrossRef - Mudgal, V., N. Madaan and A. Mudgal, 2010. Biochemical mechanisms of salt tolerance in plants: A review. Int. J. Bot., 6: 136-143.
CrossRefDirect Link - Matysik, J.A., B. Bhalu and P. Mohanty, 2002. Molecular mechanism of quenching of reactive oxygen species by proline under stress in plants. Curr. Sci., 82: 525-532.
Direct Link - Misra, A.N., S.M. Sahu, M. Misra, P. Singh and I. Meera et al., 1997. Sodium chloride induced changes in leaf growth and pigment and protein contents in two rice cultivars. Biol. Plant., 39: 257-262.
CrossRefDirect Link - Moftah, A.E. and B.E. Michel, 1987. The effect of sodium chloride on solute Potential and Proline accumulation in soybean leaves. Plant Physiol., 83: 238-240.
Direct Link - Munns, R., 1993. Physiological processes limiting plant growth in saline soils: Some dogmas and hypotheses. Plant Cell Environ., 16: 15-24.
CrossRefDirect Link - Munns, R., 2002. Comparative physiology of salt and water stress. Plant Cell Environ., 25: 239-250.
CrossRefDirect Link - Myers, B., T. Neales and M. Jones, 1990. The influence of salinity on growth, water relations and photosynthesis in Diplachne fusca (L.) P. Beauv. Ex Roemer & schultes. Functional Plant Biol., 17: 675-691.
CrossRefDirect Link - Omielan, J.A., E. Epstein and J. Dvorak, 1991. Salt tolerance and ionic relations of wheat as affected by individual chromosomes of salt-tolerant Lophopyrum elongatum. Genome, 34: 961-974.
CrossRefDirect Link - Saleethong, P., J. Sanitchon, K. Kong-ngern and P. Theerakulpisut, 2011. Pretreatment with spermidine reverses inhibitory effects of salt stress in two rice (Oryza sativa L.) cultivars differing in salinity tolerance. Asian J. Plant Sci., 10: 245-254.
CrossRefDirect Link - Stewart, R.R.C. and J.D. Bewley, 1980. Lipid peroxidation associated with accelerated ageing of soybean axes. Plant Physiol., 65: 245-248.
Direct Link - Theerakulpisut, P., S. Bunnag and K. Kong-Ngern, 2005. Genetic diversity, salinity tolerance and physiological responses to control of NaCl of six rice (Oryza sativa L.) cultivars. Asian J. Plant Sci., 4: 562-573.
CrossRefDirect Link - Uchida, A., A.T. Jagendorf, T. Hibino, T. Takabe and T. Takabe, 2002. Effects of hydrogen peroxide and nitric oxide on both salt and heat stress tolerance in rice. Plant Sci., 163: 515-523.
CrossRefDirect Link - Vaidyanathan, H., P. Sivakumar, R. Chakrabarty and G. Thomas, 2003. Scavenging of reactive oxygen species in NaCl-stressed rice (Oryza sativa L.)-differential response in salt-tolerant and sensitive varieties. Plant Sci., 165: 1411-1418.
CrossRefDirect Link - Zhu, B., J. Sua, M. Changa, D.P.S. Verma, Y.L. Fan and R. Wu, 1998. Overexpression of a Δ1 pyrroline-5-carboxylate synthetase gene and analysis of tolerance to water-and salt-stress in transgenic rice. Plant Sci., 139: 41-48.
CrossRef
Haidar Abbas Reply
Respected sir,
I am research scholar in INDIA and i am in need for this article to write my thesis so kindly send me this article on this email.
Thanking you