
B. Dhir
Department of Genetics, University of Delhi South Campus, New Delhi-110021, India
S.A. Nasim
Environmental Biotechnology Laboratory, Department of Botany, Hamdard University, New Delhi, 110062, India
S. Samantary
Department of Botany, Sailabala Women�s College, Cuttack, Odisha, India
S. Srivastava
Department of Genetics, University of Delhi South Campus, New Delhi-110021, India
International Journal of Botany
Year: 2012 | Volume: 8 | Issue: 3 | Page No.: 153-158







ABSTRACT
Accumulation of osmolytes in terrestrial plants in response to environmental stresses is well reported and information about aquatic plants is limited. Present study aimed to investigate if the aquatic weed, Salvinia natans accumulates osmolytes/compatible solutes on exposure to various heavy metals. Plants exposed to heavy metals viz. Cd, Cu, Ni, Cr, Mn, Fe, Co, Pb and Zn, were harvested after 48 h and various osmolytes including sucrose, mannitol, proline, glycine betaine and polyamines were estimated using biochemical methods. Results suggested that heavy metal stress does trigger the accumulation of osmolytes such as sucrose, mannitol and glycine betaine. In contrast proline accumulation was not observed. Studies of heavy metal stress on the endogenous levels of polyamines showed presence of free polyamines, while conjugated and bound forms were not detected. Among free polyamines, Putrescine (Put) and Spermidine (Spd) did not show significant decrease in heavy metal exposed Salvinia except Pb and Fe exposed plants. Spermine (Spm) content showed decline in heavy metal exposed Salvinia. The decrease in polyamine levels indicated their possible role in combating oxidative stress induced by heavy metals. Studies suggest that accumulation of osmolytes under heavy metal stress might help in imparting tolerance in Salvinia.
PDF Abstract XML References Citation
Received: February 17, 2012;
Accepted: June 25, 2012;
Published: October 10, 2012
How to cite this article
B. Dhir, S.A. Nasim, S. Samantary and S. Srivastava, 2012. Assessment of Osmolyte Accumulation in Heavy Metal Exposed Salvinia natans. International Journal of Botany, 8: 153-158.
DOI: 10.3923/ijb.2012.153.158
URL: https://scialert.net/abstract/?doi=ijb.2012.153.158
DOI: 10.3923/ijb.2012.153.158
URL: https://scialert.net/abstract/?doi=ijb.2012.153.158
INTRODUCTION
Accumulation of compatible organic osmolytes in plants in response to various environmental stresses viz., drought, salinity, extreme temperatures, UV radiation and heavy metals is well reported in literature (Serraj and Sinclair, 2002; Ashraf and Foolad, 2007). These are low molecular weight, highly soluble compounds. The major contribution of these osmolytes lies in protection of plants from abiotic stresses via different mechanisms including adjustment of cellular osmoticum, detoxification of reactive oxygen species, maintenance of membrane integrity and stabilization of enzymes/proteins. Apart from this, they are also known to protect cellular components from dehydration and injury (Sharma and Dietz, 2006; Ashraf and Foolad, 2007). These solutes include sugars (glucose, fructose, sucrose, trehalose, raffinose), sugar alcohols, nitrogen-containing compounds such as proline, Quaternary Amino Compounds (QACs) such as glycine betaine, alanine betaine, proline betaine and polyamines (Mudgal et al., 2010).
Potential of aquatic plants for accumulation of heavy metals is well studied (Mishra and Tripathi, 2008; Peng et al., 2008; Verma et al., 2008; Dhir et al., 2009a; Ndimele and Jimoh, 2011). The accumulation of toxic heavy metals in plants induces osmotic stress that in turn may initiate synthesis of metabolites that play an important role in metal binding, antioxidant defence and signaling (Sharma and Dietz, 2006; Joseph and Jini, 2010; Bhat and Khan, 2011). Accumulation of osmolytes in response to heavy metal stress has been documented for terrestrial species but information regarding aquatic species is lacking (Sivaci et al., 2008). Therefore, present investigations were carried out with an aim to investigate if aquatic plants also show accumulation of osmolytes in response to heavy metal stress. Salvinia natans, a free-floating aquatic weed having high growth rate and tolerance to high element concentrations was chosen for the present Previous findings by our group proved that Salvinia natans possesses significant potential to accumulate various heavy metals (Dhir et al., 2008, 2009b). Therefore, further investigations were carried out with an aim to: (1) check if Salvinia accumulate osmolytes under heavy metal stress and (2) investigate the alterations in the level of endogenous polyamines in heavy metal exposed Salvinia.
MATERIALS AND METHODS
Plant material and growth conditions: Plants of Salvinia natans L. (Salviniaceae) collected from unpolluted water bodies were maintained in cemented pots (~1 m diameter) under natural light in outdoor conditions. The temperature ranged from 30-32°C. The solutions having metal concentration 35 mg L-1 were prepared by using metal salts Cd(NO3)2.4H2O (Cd2+), CuSO4.5H2O (Cu2+), Ni(NO3)2.6H2O (Ni2+), K2Cr2O7 (Cr6+), MnCl2.4H2O (Mn2+), FeSO4.7H2O (Fe2+), CoCl2.6H2O (Co2+), Pb(NO3)2 (Pb2+) and ZnSO4.7H2O (Zn2+). The pH of all the solutions ranged between 4.5-5.0. Distilled water was taken as control. Five plants of Salvinia (each having five nodes with two leaves at each node) were floated in each tub having 750 mL of metal solution. All the measurements were carried out after 48 h of metal exposure.
Sucrose estimation: Sugars were extracted by overnight submersion of dried plant material in 80% (v/v) ethanol at 25°C with periodic shaking. Sucrose content was determined first by degrading reactive sugars present in 0.1 mL extracts with 0.1 mL 5.4 N KOH at 97°C for 10 min. Three mL of freshly prepared anthrone reagent were then added to the cooled reaction product and the mixture was heated at 97°C for 5 min, cooled and absorbance was read at 620 nm (El-Shihaby et al., 2002). Sucrose content was expressed as μg g-1 fresh wt.
Estimation of mannitol: Leaves were ground in liquid nitrogen, lyophilized overnight and stored at -25°C until used. The lyophilized powder was placed in a capped 1.5 mL Eppendorf tube and 1 mL of hot (80°C) distilled water was added. The tube was heated at 80°C for 30 min and then cooled and centrifuged at 12000 g for 15 min twice. Plant extracts were deionized by passage through cationic and anionic resins.
The 0.1 mL of biological extract was dispensed in microcentrifuge tube and 0.5 mL of 0.5 M formate (pH 3.0) was added. To this solution 0.3 mL of 5 mM sodium periodate (reagent 1) was added. The contents were vortexed and left at room temperature for 15 sec and 0.3 mL of a solution containing 0.1 M acetylacetone, 2 M ammonium acetate and 0.02 M sodium thiosulfate (reagent 2), were added. The tube was closed and heated in boiling water for 2 min and cooled under running tap water and the absorbance at 412 nm was measured (Sanchez, 1998.). The mannitol content was expressed in nM g-1 fresh wt.
Proline estimation: Proline was measured in leaf tissues following method described by Bates et al. (1973). The optical density was measured at 520 nm and proline content was expressed as μg g-1 fresh wt.
Glycine betaine estimation: Glycine betaine was estimated in leaf tissues following modified protocol of Desingh and Kanagaraj (2007). The absorbance was measured at 365 nm and glycine betaine content was expressed as μg g-1 fresh weight.
Estimation of polyamines: Polyamines (free, conjugated and bound forms) present in leaf tissue were quantified according to modified protocol of Flores and Galston (1982). The polyamines were extracted in perchloric acid. Supernatant was used for estimation of free polyamines, while supernatant treated with HCl was used for estimation of conjugated polyamines. The pellet was dissolved in 1 N NaOH was used for estimation of bound polyamines. Two hundred microliter each of supernatent (untreated), supernatent hydrolysate and pellet hydrolysate were taken separately. Two hundred microliter of saturated Na2CO3 and 400 μL of dansyl chloride (5 mg mL-1 in acetone) were added to each tube, vortexed and incubated at 25°C in dark (overnight). 300 μL of proline (100 mg mL-1) was added to dansylation mixture and samples were incubated in dark for 30 min at room temperature. To extract polyamines, 50 μL benzene was added to dansylation mixture, vortexed and allow to stand till the layers separate. Equal volumes of polyamine solutions and standards were applied and resolved by Thin-Layer Chromatography (TLC) with cyclohexane and ethylacetate in a ratio of 5:4 (v/v) as the solvent. The polyamines were located with a UV detector, polyamine-containing silica spots were collected and eluted with 4 mL of ethyl acetate. The fluorescence of these solutions was measured with a UV-fluorescence spectrophotometer (Hitachi, F-2000) at 359 nm (excitation) and 495 nm (emission) wavelengths, respectively (Chang et al., 1999).
Statistical analysis: Analysis of Variance (ANOVA) for all measured variables was performed by using software new MSTAT-C (version 2.1). The level of significance was measured using Duncan’s Multiple Range Test (DMRT) taking p≤0.05 as significant.
RESULTS AND DISCUSSION
Sucrose and mannitol levels: Overproduction of compatible organic solutes is one of the most common stress responses in plants (Serraj and Sinclair, 2002). Exposure to heavy metal stress induced a substantial increase in osmolyte content (Table 1). Sucrose levels recorded enhancement in plants exposed to heavy metals, though the response varied for each metal. The increase was significant in Cr, Cd and Zn exposed Salvinia. Cadmium (352), Cr (317)and Zn (293) exposed plants showed ~3 fold increase, while Ni (268), Pb(250), Fe(239), Mn(202), Co(214) and Cu(184) exposed plants showed ~2 fold enhancement in sucrose level in comparison to control (108).
| Table 1: | Alterations in various solutes measured after 48 h of metal exposure |
 | |
| Values are Mean±SEM of three independent experiments with three replicates each, Different letters in a row are significantly different at p≤0.05 | |
Sucrose functions as an osmoprotectant under stress conditions and has been postulated to possess ROS-scavenging capacity (Ende and Valluru, 2008). The increase in sucrose content can be correlated to its reduced utilization during stress and/or block in its transport rather than overproduction (El-Shihaby et al., 2002). Heavy metal induced increase in soluble sugars particularly sucrose has been reported (Alaoui-Sosse et al., 2004; Rosa et al., 2009).
Mannitol, a sugar alcohol plays an important role in storage of carbon and energy, regulation of coenzymes, osmoregulation and free radical scavenging (Prabhavathi and Rajam, 2007). Heavy metal exposed Salvinia showed increase in mannitol content except Cu, Pb and Mn exposed plants where no change was observed. Plants exposed to Co (796) and Ni (547) showed ~4 and ~2.5 fold increase, while Cd, Cr (471, 443) exposed plants showed ~2 fold enhancement in mannitol level in comparison to control (208). The increase in mannitol content suggests its possible role as a free radical scavenger thus preventing cells from lipid peroxidation (Sickler et al., 2007).
Glycine betaine and proline levels: Glycine betaine levels increased in plants exposed to metal stress except for Cr and Mn, where no change was observed. The increase was more significant in Cu, Fe and Co exposed plants. Glycine betaine levels increased by ~1.9-1.6 fold in Cu, Fe and Co (342, 322, 293) exposed plants, while ~1.2 to 1.3 fold increase was noted in Pb, Zn, Ni and Cd (209, 219, 222, 237) exposed plants in comparison to control (179). Similar response in terms of glycine betaine accumulation under environmental stress has been reported both in terrestrial and aquatic plant species such as Spartina and Phragmites (Zhu et al., 2003; Al-Garni, 2006; Islam et al., 2009). Glycine betaine is shown to confer tolerance in Cd exposed tobacco by protecting cellular components and increasing activities of antioxidant enzymes (Islam et al., 2009). Glycine betaine (GB), a quaternary ammonium compound, plays a vital role in protection of enzyme like RuBisCo, maintenance of membrane integrity and osmotic potential. All these roles thus protect photosynthetic efficiency (Chen and Murata, 2002; Shirasawa et al., 2006; Kattab, 2007).
Proline, an amino acid is reported to play role in osmotic adjustment and maintenance of cellular integrity (e.g. membranes and proteins) via scavenging free radicals, maintaining cellular redox potential and NADP+/NADPH ratios (Sharma and Dietz, 2006; Ashraf and Foolad, 2007). Salvinia exposed to heavy metal stress showed decline in proline accumulation. Decline was more significant in plants exposed to Zn, Fe and Ni. Proline levels noted decline of ~3 fold in plants exposed to Ni, Fe, Zn, (25, 24, 22) while exposure to Cd and Co (34, 30) led to ~2 fold decrease in comparison to control (Table 1). This is in accordance with earlier studies where aquatic species viz. Ceratophyllum, Wolffia, Hydrilla and Lemna polyrhiza exhibited decline in proline levels in response to Cd stress (Dhir et al., 2004; John et al., 2008). Similar response of decrease in proline content has been documented for Cu exposed Spirodela polyrrhiza (Xing et al., 2010).
Polyamines: Polyamines including spermidine (Spd), spermine (Spm) and their obligate precursor, putrescine (Put), are polybasic aliphatic amines that influence variety of growth and development processes in plants (Liu et al., 2007). They play role in wide range of basic cellular regulatory processes including DNA replication, transcription, translation, cell division, modulation of enzyme activities, cellular cation anion balance and membrane stability due to their potent binding ability to negatively charged macromolecules and membranes. They function as signalling molecules, antioxidants and act as second messengers (Verma and Mishra, 2005). Though, specific role of polyamines in plants under metal stress is not yet known but there is a strong possibility that they can effectively stabilize and protect the membrane systems against the toxic effects of metal ions particularly the redox active metals. Their role of metal chelators has been postulated (Sharma and Dietz, 2006; Groppa et al., 2007; Gill and Tuteja, 2010). Being polycationic in nature, they bind to negatively charged groups in the cell membrane. Spermine, in particular has been noted to acts as a free radical scavenger and is capable of quenching chemically generated singlet oxygen (Groppa et al., 2007; Wen et al., 2010).
| Table 2: | Alterations in level of free polyamines observed after 48 h of metal exposure. |
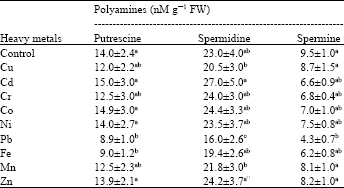 | |
| Values are Mean±SEM of three independent experiments with three replicates each, Different letters in a column are significantly different at p≤0.05 | |
Spermine has been suggested to play a protective role against the oxidative damage produced by metals. Moreover, role of other polyamines in reducing ROS formation by inhibiting NADPH oxidase activity has been reported (Wen et al., 2010). Role of spermidine as a stress-protecting compound and stress-signaling regulator has been noted (Kasukabe et al., 2004).
Polyamines have been considered to act as antioxidants and reduce oxidative damage produced by metals. Salvinia showed accumulation of free polyamines, while conjugated and bound polyamines were not detected (Table 2). Among free polyamines, putrescine and spermidine was present in higher amount, while spermine accumulated to a lesser extent. Polyamine levels showed a variation in response to each metal exposure. Putrescine levels did not show any significant difference in comparison to control. A slight increase was noted in Co and Cd exposed Salvinia, while no difference was observed in Ni and Zn exposed plants in comparison to control. In contrast, decline in putrescine levels was noted in plants exposed to other metals (Cr, Cu, Pb, Mn and Fe). Spermidine levels showed increase in Cd, Cr, Co and Zn exposed plants, though increase was significant in Cd (27) exposed Salvinia (Table 2). In contrast, a decline was noted in plants exposed to Ni, Pb, Cu, Mn and Fe. Spermine levels showed a general decline in plants exposed to metal stress. Stress conditions can affect polyamine metabolism in different manner that includes an increase or decrease of endogenous polyamines (Liu et al., 2007). Increase in free putrescine and spermidine levels noted in Cd and Co exposed Salvinia could be due to enhanced de novo synthesis resulting from increased activity of enzymes involved in putrescine biosynthesis [Arginine Decarboxylase (ADC) or Ornithine Decarboxylase (ODC)] or reduced degradation, although the exact mechanism remains unclear (Liu et al., 2007). Similar increase in putrescine and decrease in spermine has been observed in wheat, Hydrocharis dubia exposed to Cd, Ni (Groppa et al., 2007; Zhao et al., 2008). Ni treatment significantly increased the putrescine (Put) level and lowered spermidine (Spd) and spermine (Spm) levels, thereby significantly reducing the ratio of free (Spd+ Spm)/Put in leaves, which has been considered as the signal under stress (Zhao et al., 2008). Significantly increase in free putrescine (Put) level and alterations in other PAs levels under Pb treatment has been noted in Potamogeton crispus (Xu et al., 2011). In contrast, heavy metal induced decrease in putrescine and spermine level has also been reported in plant species (Groppa et al., 2007; Zhao et al., 2008).
CONCLUSIONS
The present investigations revealed that Salvinia produced osmolytes that helped in combating osmotic stress induced by heavy metals, though the response varied from metal to metal. In summary, Salvinia possesses effective metabolic machinery that is capable of overcoming the osmotic stress induced by heavy metals. This is evident from enhanced accumulation of metabolites viz. glycine betaine, mannitol and sucrose. Decline in the level of metabolites such as proline and polyamines may not suggest a direct role but they might contribute in imparting tolerance in Salvinia.
REFERENCES
- Al-Garni, S.M.S., 2006. Increasing salt tolerance of a halophytic plant Phragmites australis by mycorrhizal symbiosis. Am. Euras. J. Agric. Environ. Sci., 1: 119-126.
Direct Link - Ashraf, M. and M.R. Foolad, 2007. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot., 59: 206-216.
CrossRefDirect Link - Alaoui-Sosse, B., P. Genet, F. Vinit-Dunand, M.L. Taussaint, D. Epron and P.M. Badot, 2004. Effect of copper on growth in cucumber plants (Cucumis sativus) and its relationships with carbohydrate accumulation and changes in ion contents. Plant Sci., 166: 1213-1218.
CrossRef - Bhat, U.N. and A.B. Khan, 2011. Heavy metals: An ambiguous category of inorganic contaminants, nutrients and toxins. Res. J. Env. Sci., 5: 682-690.
CrossRef - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Chang, S.C., P.B. Kaufman, and B.G. Kang, 1999. Changes in endogenous levels of free polyamines during petiole elongation in the semiaquatic plant Ranunculus sceleratus. Int. J. Plant Sci., 160: 691-697.
Direct Link - Chen, T.H.H. and N. Murata, 2002. Enhancement of tolerance of abiotic stress by metabolic engineering of betaines and other compatible solutes. Curr. Opin. Plant Biol., 5: 250-257.
CrossRefPubMedDirect Link - Desingh, R. and G. Kanagaraj, 2007. Influence of salinity stress on photosynthesis and antioxidative systems in two cotton varities. Genet. Appl. Plant Physiol., 33: 221-234.
Direct Link - Dhir, B., P. Sharmila and P.P. Saradhi, 2004. Hydrophytes lack potential to exhibit cadmium stress induced enhancement in lipid peroxidation and accumulation of proline. Aquat. Toxicol., 66: 141-147.
CrossRefDirect Link - Dhir, B., Sharmila, P. and P.P. Saradhi, 2008. Photosynthetic performance of Salvinia natans exposed to chromium and zinc rich wastewater. Braz. J. Plant Physiol., 20: 61-70.
CrossRefDirect Link - Dhir, B., P. Sharmila, P.P. Saradhi and S.A. Nasim, 2009. Physiological and antioxidant responses of Salvinia natans exposed to chromium-rich wastewater. Ecotoxicol. Environ. Saf., 72: 1790-1797.
CrossRefPubMedDirect Link - Dhir, B., P. Sharmila, P.P. Saradhi and S.A. Nasim, 2009. Heavy metal removal potential of dried Salvinia biomass. Int. J. Phytoremediation, 12: 133-141.
CrossRefPubMedDirect Link - El-Shihaby, O.A., M.M. Alla, M.E. Younis and Z.M. EL-Bastawisy, 2002. Effect of kinetin on photosynthetic activity and carbohydrate content in waterlogged or seawater-treated Vigna sinensis and Zea mays plants. Plant Biosystems, 136: 277-290.
CrossRef - Ende, W.V. and R. Valluru, 2008. Sucrose, sucrosyl oligosaccharides, and oxidative stress: Scavenging and salvaging?. J. Exp. Bot., 60: 9-18.
CrossRefDirect Link - Groppa, M.D., M.L. Tomaro and M.P. Benavides, 2007. Polyamines and heavy metal stress: The antioxidant behavior of spermine in cadmium- and copper-treated wheat leaves. BioMetals, 20: 185-195.
CrossRef - Islam, M.M., M.A. Hoque, E. Okuma, M.N.A. Banu, Y. Shimoishi, Y. Nakamura and Y. Murata, 2009. Exogenous proline and glycinebetaine increase antioxidant enzyme activities and confer tolerance to cadmium stress in cultured tobacco cells. J. Plant Physiol., 166: 1587-1597.
CrossRefPubMedDirect Link - John, R., P. Ahmad, K. Gadgil and S. Sharma, 2008. Effect of cadmium and lead on growth, biochemical parameters and uptake in Lemna polyrrhiza L. Plant Soil Environ., 54: 262-270.
Direct Link - Joseph, B. and D. Jini, 2010. Insight into the role of antioxidant enzymes for salt tolerance in plants. Int. J. Bot., 6: 456-464.
CrossRefDirect Link - Kasukabe, Y., L. He, K. Nada, S. Misawa, I. Ihara and S. Tachibana, 2004. Overexpression of spermidine synthase enhances tolerance to multiple environmental stresses and up-regulates the expression of various stress-regulated genes in transgenic Arabidopsis thaliana. Plant Cell Physiol., 45: 712-722.
CrossRefPubMedDirect Link - Kattab, H., 2007. Role of glutathione and polyadenylic acid on the oxidative defense systems of two different cultivars of canola seedlings grown under saline condition. Aust. J. Basic Applied Sci., 1: 323-334.
Direct Link - Liu, J.H., H. Kitashiba, J. Wang, Y. Ban and T. Moriguchi, 2007. Polyamines and their ability to provide environmental stress tolerance to plants. Plant Biotechnol., 24: 117-126.
Direct Link - Mishra, V.K. and B.D. Tripathi, 2008. Concurrent removal and accumulation of heavy metals by the three aquatic macrophytes. Bioresour. Technol., 99: 7091-7097.
CrossRef - Mudgal, V., N. Madaan and A. Mudgal, 2010. Biochemical mechanisms of salt tolerance in plants: A review. Int. J. Bot., 6: 136-143.
CrossRefDirect Link - Ndimele, P.E. and A.A. Jimoh, 2011. Water hyacinth (Eichhornia crassipes (Mart.) Solms.) in phytoremediation of heavy metal polluted water of Ologe Lagoon, Lagos, Nigeria. Res. J. Environ. Sci., 5: 424-433.
CrossRefDirect Link - Rosa, M., C. Prado, G. Podazza, R. Interdonato, J.A. Gonzalez, M. Hilal and F.E. Prado, 2009. Soluble sugars-Metabolism, sensing and abiotic stress A complex network in the life of plants. Plant Signal Behav., 4: 388-393.
CrossRefDirect Link - Serraj, R. and T.R. Sinclair, 2002. Osmolyte accumulation: Can it really help increase crop yield under drought conditions? Plant Cell Environ., 25: 333-341.
CrossRefPubMedDirect Link - Sharma, S.S. and K.J. Dietz, 2006. The significance of amino acids and amino acid-derived molecules in plant responses and adaptation to heavy metal stress. J. Exp. Bot., 57: 711-726.
CrossRefDirect Link - Shirasawa, K., T. Takabe, T. Takabe and S. Kishitani, 2006. Accumulation of glycinebetaine in rice plants that overexpress Choline monooxygenase from spinach and evaluation of their tolerance to abiotic stress. Ann.Bot., 97: 565-571.
CrossRefDirect Link - Sivaci, A., E. Elmas and F. Gumus, 2008. Changes in abscisic acid contents of some aquatic plants exposed to cadmium and salinity. Int. J. Bot., 4: 104-108.
CrossRefDirect Link - Gill, S.S. and N. Tuteja, 2010. Polyamines and abiotic stress tolerance in plants. Plant Signal Behav., 5: 26-33.
Direct Link - Verma, S. and S.N. Mishra, 2005. Putrescine alleviation of growth in salt stressed Brassica juncea by inducing antioxidative defence system. J. Plant Physiol., 162: 669-677.
CrossRef - Xing, W., W. Huang and G. Liu, 2010. Effect of excess iron and copper on physiology of aquatic plant Spirodela polyrrhiza (L.) Stilled. Environ. Toxicol., 25: 103-112.
CrossRef - Zhu, X.Y., Y. Jing, G.C. Chen, S.M. Wang and C.L. Zhang, 2003. Solute levels and osmoregulatory enzyme activities in reed plants adapted to drought and saline habitats. Plant Growth Reg., 41: 165-172.
CrossRef