
Homa Mahmoodzadeh
Department of Biology, Islamic Azad University, Mashhad Branch, Mashhad, Iran
LiveDNA: 98.25020
Elham Esparham
Department of Biology, Islamic Azad University, Fars Science and Research Branch, Iran
International Journal of Botany
Year: 2011 | Volume: 7 | Issue: 2 | Page No.: 195-199







ABSTRACT
The plant hormone abscisic acid (ABA), as a stress signal, plays important roles in the regulation of plant responses to environmental stresses. The relationship among abscisic acid (ABA) accumulation, the generation of Reactive Oxygen Species (ROS) and the activities of several antioxidant enzymes such as Catalase (CAT), Ascorbate Peroxidase (APX) and Glutathione Reductase (GR) was investigated in leaves of detached bean (Phaseolus vulgaris L.) plants exposed to 10,100 and 1000 μM ABA [Detached leaves of bean were cut into leaf segments and floated in solutions containing 0, 10, 100 and 1,000 μM ABA, at 25°C and then used for assay of antioxidant enzymes]. Treatment with 10 and 100 μM ABA significantly increased the levels of O2¯ and H2O2, followed by an increase in activities of CAT, APX and GR in leaves of bean seedlings. Treatment with 1,000 μM ABA led to a more abundant generation of O2¯and H2O2 .The activities of these antioxidative enzymes were reduced when compared with the treatment of 100 μM ABA. These results indicate that treatment with low concentrations of ABA (10 to 100 μM) induced an antioxidative defense response against oxidative damage, but a high concentration of ABA (1,000 μM) induced an excessive generation of ROS and led to an oxidative damage in plant cells.
PDF Abstract XML References Citation
Received: May 31, 2011;
Accepted: August 02, 2011;
Published: August 30, 2011
How to cite this article
Homa Mahmoodzadeh and Elham Esparham, 2011. Changes in Hydrogen Peroxide Content and Antioxidant Enzymes in Abscisic Acid-induced Antioxidant Defense in Leaves of Bean Seedlings. International Journal of Botany, 7: 195-199.
DOI: 10.3923/ijb.2011.195.199
URL: https://scialert.net/abstract/?doi=ijb.2011.195.199
DOI: 10.3923/ijb.2011.195.199
URL: https://scialert.net/abstract/?doi=ijb.2011.195.199
INTRODUCTION
The phytohormone abscisic acid (ABA) regulates many important aspects of plant growth and development and its main function is to regulate plant adaptive responses to various adverse environmental conditions (Saleh, 2007; Cha-Um et al., 2007; Hou et al., 2010; Fatima and Bano, 1999; Ayub et al., 2000; Sivaci et al., 2008; Anjum et al., 1999; Kumari, 2010). How the ABA signal is transduced into a physiological or biochemical response has been an interesting research subject in the recent years.
Increasing evidence indicates that one mode of ABA action may be related to its role in the oxidative stress in plant cells. It has been documented that ABA can cause an increased generation of O2¯(Jiang and Zhang, 2001; Laloi et al., 2004; Hu et al., 2005) and H2O2 (Guan et al., 2000; Pei et al., 2000; Jiang and Zhang, 2001) and CAT (Guan et al., 2000). Meanwhile, ABA also increases the activities of antioxidant enzymes such as SOD, CAT, APX, GR and MDA in plant tissues (Bellaire et al., 2000; Jiang and Zhang, 2001; Hu et al., 2005; Fryer et al., 2003; Park et al., 2004; Khorshidi and Nojavan, 2006).
Although, it has been shown that ABA can result in an oxidative stress, an enhancement in the capacity of oxidative stress tolerance may imply that plants need to mobilize the whole antioxidant defense systems including enzymatic and non-enzymatic constituents to resist oxidative damage in stressed plant tissues, rather than a few enzymes or metabolites. Meanwhile, since ABA causes oxidative stress in plants, it is possible that an oxidative damage might take place in plants exposed to the treatment of high concentrations of ABA. Different concentrations of ABA from 1 to 1,000 μM have been used to different plant tissues and have been shown to be able to induce gene expression and protein synthesis involved in antioxidative defences (Bellaire et al., 2000; Guan et al., 2000). In addition to H2O2, it is not clear whether other Reactive Oxygen Species (ROS), such as O2¯and ·OH, are involved in oxidative stress in plants caused by ABA (Jiang and Zhang, 2001).
In this study, the effects of different concentrations of ABA on the levels of ROS including O2¯ and H2O2, the activities of antioxidant enzymes such as CAT, APX and GR were systematically studied in details. The objective of the present study was to find out a possible relationship between ABA, ROS, antioxidative defences and oxidative damage in bean plants.
MATERIALS AND METHODS
Plant material: The experiment was conducted in Faculty of Science, Mashad Branch, Islamic Azad University, Iran (2010).
The seeds of bean were supplied by the Agricultural Research Center of Khorasan, Iran. The plant Seeds were surface-sterilized with 5% NaHClO for 10 min and germinated at 28°C for 1 day. The seedlings were grown in trays of sand in a greenhouse at a temperature of 25-30°C and photoperiod of 14/10 h (day night) and watered daily. When the second leaf was fully expanded, they were collected and used for investigations.
Abscisic acid treatments: Detached leaves were cut into leaf segments (3 cm in length), rinsed with distilled water and then floated in solutions containing 0, 10, 100 and 1,000 μM ABA, at 25°C. Leaf segments were vacuum-infiltrated with these solutions before the incubation and immediately frozen in liquid N2 and then stored at -80°C for further analysis.
Assays of antioxidant enzyme activities: Frozen leaf segments (0.5 g) were crushed into fine powder in a mortar and pestle under liquid N2. Soluble proteins were extracted by homogenizing the powder in 10 mL of 50 mM potassium phosphate buffer (pH 7.0) containing 1 mM EDTA and 1% polyvinylpyrrolidone, with the addition of 1 mM ASC in the case of APX assay. The homogenate was centrifuged at 15,000x g for 20 min at 4°C and the supernatant was used for the following enzyme assays. CAT (EC1.11.1.6) activity was determined by following the consumption of H2O2 (extinction coefficient 39.4 mM-1 cm-1) at 240 nm for 3 min (Aebi, 1984). The reaction mixture contained 50 mM potassium phosphate buffer (pH7.0), 10 mM H2O2 and 200 μL of enzyme extract in a 3 mL volume.
GR (EC1.6.4.2) activity was determined by following the oxidation of NADPH at 340 nm (extinction coefficient 6.2 mM-1 cm-1) for 3 min in 1 mL of an assay mixture containing 50 mM potassium phosphate buffer (pH 7.8), 2 mM Na2 EDTA, 0.15 mM NADPH, 0.5 mM GSSG and 200 μmL of enzyme extract. The reaction was initiated by adding NADPH. Corrections were made for the background absorbance at 340 nm, without NADPH (Shaedle and Bassham, 1977).
APX (EC1.11.1.11) activity was determined by following the decrease in A290 (extinction coefficient 2.8 mM-1 cm-1) for 1 min in 1 mL of a reaction mixture containing 50 mM potassium phosphate buffer (pH 7.0), 0.5 mM ASC, 0.1 mM H2O2 and 200 μL of enzyme extract. The reaction was started by adding enzyme extract. Correction was done for the low, non-enzymatic oxidation of ASC by H2O2 (Nakano and Asada, 1981).
Determination of superoxide radical and hydrogen peroxide: O2¯ was measured as described by Erich and Heupel (1976) by monitoring the nitrite formation from hydroxylamine in the presence of O2¯, with some modifications. One g of frozen leaf segments was homogenized with 3 mL of 65 mM potassium phosphate buffer (pH 7.8) and centrifuged at 5,000x g for 10 min. The incubation mixture contained 0.9 mL of 65 mM phosphate buffer (pH 7.8), 0.1 mL of 10 mM hydroxylamine hydrochloride and 1 mL of the supernatant. After incubation at 25°C for 20 min, 17 mM sulfanilamide and 7 mM-naphthylamine were added to the incubation mixture. After reaction at 25°C for 20 min, ethyl ether in the same volume was added and centrifuged at 1,500x g for 5 min. The absorbance in the aqueous solution was read at 530 nm. A standard curve with NO2¯ was used to calculate the production rate of O2¯ from the chemical reaction of O2¯ and hydroxylamine.
The content of H2O2 was measured by monitoring the A415 of the titanium-peroxide complex following the method described by Brennan and Frenkel (1977). Absorbance values were calibrated to a standard curve generated with known concentrations of H2O2.
RESULTS
Effects of different concentrations of ABA on levels of O2¯ and H2O2 in leaves of bean seedlings were investigated. Treatment with 10, 100 and 1,000 μM ABA caused a significant increase in levels of O2¯ (Fig. 1) and H2O2 (Fig. 2) compared with the control. At 12 h, O2¯ levels were 0.7, 0.8 and 1.4 (μmol g-1 dry wt. min) respectively, for the three concentrations of ABA used, that increased by 23, 40 and 63% in comparison with the control. H2O2 levels were 0.8, 1 and 1.5 (μmol g-1 fwt.) that increased by 39, 51 and 66% to the control.
Figure 3-5 show the effects of different concentrations of ABA on several antioxidative enzymes such as CAT (Fig. 3), the enzyme mainly responsible for eliminating H2O2 in the peroxisomes and APX (Fig. 4) and GR (Fig. 5), the two key enzymes of the Halliwell-Asada pathway for the removal of H2O2 in the chloroplasts. Treatment with 10 and 100 μM ABA caused a continuous increase in the activities of the three antioxidative enzymes .
 | |
| Fig. 1: | Effects of ABA at different concentrations on levels of O2¯in leaves of bean seedlings. Values are the means of three different experiments. Error bars represent SE with n = 3 |
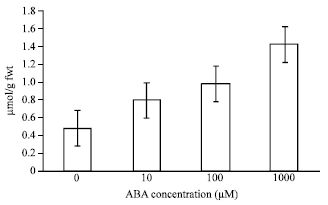 | |
| Fig. 2: | Effects of ABA at different concentrations on levels of H2O2 in leaves of bean seedlings. Values are the means of three different experiments. Error bars represent SE with n = 3 |
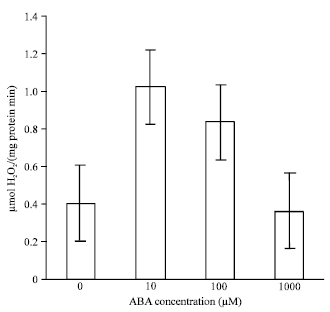 | |
| Fig. 3: | Effects of ABA at different concentrations on activities of, CAT in leaves of bean seedlings. Values are the means of three different experiments. Error bars represent SE with n =3 |
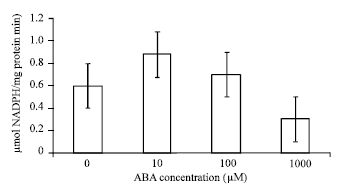 | |
| Fig. 4: | Effects of ABA at different concentrations on activities of GR in leaves of bean seedlings. Values are the means of three different experiments. Error bars represent SE with n = 3 |
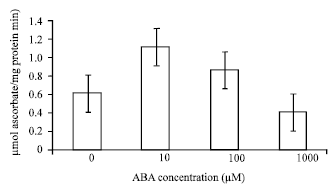 | |
| Fig. 5: | Effects of ABA at different concentrations on activities of APX in leaves of bean seedlings. Values are the means of three different experiments. Error bars represent SE with n = 3 |
The leaves treated with 10 and 100 μM ABA enhanced the activities of CAT by 1 and 0.8 μmol H2O2/(mg protein min) i.e, 60 and 52%, respectively; when compared with the control (Fig. 3); GR by 0.9 and 0.7 μmol NADPH/(mg protein min) i.e, 35 and 16% (Fig. 4); APX by 1.15 and 0.9 μmol ascorbate/ (mg protein min) i.e, 45 and 29% (Fig. 5). Activities of these antioxidative enzymes reduced in leaves treated with 1000 μM ABA in comparison with the control; CAT by 0.35 (10%), GR by 0.3 (23%) and APX by 0.4 (33%).
DISCUSSION
It has been shown that ABA can cause an increased production of H2O2, induce the expression of some antioxidant genes and enhance the activities of antioxidative enzymes such as SOD, CAT, GPX, APX and GR in plants (Bellaire et al., 2000; Guan et al., 2000). These suggest that ABA can result in an oxidative stress in plants. Our results not only have shown that ABA induced an increase in the content of H2O2 and the activities of CAT, APX and GR in leaves of bean seedlings but also further indicated that treatment of ABA led to an increase in the level of O2¯. These results further support that ABA results in an oxidative stress in plants.
ROS are generally believed as the by-products of oxygen metabolism and can have adverse effects such as lipid peroxidation, denaturation of proteins and mutation of DNA, all considered as cellular oxidative damage. Abundant evidence, however, has shown that ROS, especially H2O2 and O2¯, are involved in cellular signaling process as secondary messengers. These induce a number of genes and proteins involved in stress defences, including CAT, GPX, GP, GR, GST, APX, SOD and pathogenesis-related (PR) protein (Bellaire et al., 2000; Guan et al., 2000). Although, ABA and ROS are thought to act as secondary messengers for the induction of antioxidant defences in stressed plants, the relationship between ABA and ROS in stress signal transduction cascades is still unclear. One possible model is that ABA-mediated metabolic changes might lead to an increase in endogenous ROS levels, which in turn induce antioxidative gene expression (Guan et al., 2000). Present results showed that a significant increase in levels of O2¯ and H2O2 occurred under ABA treatment and the increase in activities of antioxidative enzymes took place; meanwhile, the enhancement in the activities of these antioxidative enzymes was closely related to the levels of ROS induced by different concentrations of ABA in leaves of bean seedlings. These findings support the hypothesis mentioned above and suggests further that the generation of ABA-induced ROS may trigger the response of whole antioxidative defense systems against oxidative stress.
Although, ROS may act as secondary messengers to regulate gene expression and protein biosynthesis involved in the stress defences, these may only occur under subtoxic condition. ROS at higher concentrations are still cytotoxic. Our data showed that 10 to 100 μM ABA treatment significantly induced the increase in the levels of O2¯ and H2O2 and enhanced the capacity of protective systems in leaves of maize seedlings. Such an enhancement in the antioxidant defences is sufficient to scavenge these increased ROS. Treatment with 1,000 μM ABA led to a more abundant generation of O2¯ and H2O2. The capacity of antioxidative defense systems was not maintained at a higher level when compared with the treatment of 100 μM ABA. A marked oxidative damage occurred in leaves of bean seedlings. These suggest that the superoptimal concentration of 1,000 μM ABA may have induced a severe oxidative stress that cannot be controlled by the enhanced antioxidative defense systems.
There is only limited information about the mechanism with which ABA leads to the generation of ROS in plant cells. One speculation is that ABA-induced stomatal closing can cause the reduction in the availability of CO2 for photosynthesis, which may lead to the generation of ROS from the misdirecting of electrons in the photosystems (Bowler et al., 1992). However, in guard cells of Arabidopsis, ABA-induced H2O2 production and the H2O2-activated Ca2+ channels are important mechanisms for ABA-induced stomatal closing (Pei et al., 2000). Diphenyleneiodonium chloride (DPI), a well-known inhibitor of trans-plasma membrane NAD (P)H oxidase (O2¯ synthase) (Orozco-Cardenas et al., 2001), blocked the production of ROS and partially inhibited ABA-induced stomatal closing. These results indicate that an oxidative burst occurred prior to the stomatal closing in the guard cells exposed to ABA treatment and suggest that a trans-plasma membrane NAD(P)H oxidase (O2¯ synthase) might be involved in the ABA signaling.
CONCLUSION
In conclusion, treatment with lower concentrations (10 to 100 μM) of ABA led to a relatively slight increase in the levels of O2G and H2O2 and induced the capacity of whole antioxidant defense systems against oxidative stress. Although, treatment with higher concentration of ABA (1,000 μM) no maintained the capacity of antioxidant defense at a higher level, a marked oxidative damage occurred.
REFERENCES
- Aebi, H., 1984. Catalase in vitro. In: Methods in Enzymology, Packer, L., Academic Press, Cambridge, Massachusetts, United States, ISBN: 9780121820053, pp: 121-126.
CrossRefDirect Link - Anjum, N., A. Bano and M. Sanaullah, 1999. Effects of spermine and abscisic acid on growth and biochemical contents of Vigna mungo L. under high temperature and salt-stress. Pak. J. Biol. Sci., 2: 1375-1377.
CrossRefDirect Link - Ayub, N., A. Bano, S. Ramzan and M. Usman, 2000. Effect of VAM on drought tolerance and growth of plant in comparison with the effect of growth regulators. Pak. J. Biol. Sci., 3: 957-959.
CrossRefDirect Link - Bellaire, B.A., J. Carmody, J. Braud, D.R. Gosset, S.W. Banks and M.C. Lucas, 2000. Involvement of abscisic acid dependent and independent pathways in the upregulation of antioxidant enzyme activity during NaCl stress in cotton callus tissue. Free Radical Res., 33: 531-545.
PubMed - Bowler, C., M.V. Montagu and D. Inze, 1992. Superoxide dismutase and stress tolerance. Annu. Rev. Plant Physiol. Mol. Biol., 43: 83-116.
CrossRefDirect Link - Brennan, T. and C. Frenkel, 1977. Involvement of hydrogen peroxide in the regulation of senescence in pear. Plant Physiol., 59: 411-416.
Direct Link - Cha-Um, S., S. Roytakul, T. Sathung, A. Maijandang, S. Tengsiriwattana and C. Kirdmanee, 2007. Exogenous glucose and abscisic acid pre-treatment in indica rice (Oryza sativa L. spp. indica) responses to sodium chloride salt stress. J. Plant Sci., 2: 141-152.
CrossRefDirect Link - Elstner, E.F. and A. Heupel, 1976. Inhibition of nitrite formation from hydroxylammoniumchloride: A simple assay for superoxide dismutase. Anal. Chem., 70: 616-620.
CrossRefPubMedDirect Link - Fatima, Z. and A. Bano, 1999. The effect of plant growth regulators and temperature shock on 1AA-oxidase activity of roots and endogenous ABA level of leaves. Pak. J. Biol. Sci., 2: 1601-1603.
CrossRefDirect Link - Fryer, M.J., L. Ball, K. Oxborough, S. Karpinski, P.H. Mullineaux and N.R. Baker, 2003. Control of Ascorbate Peroxidase 2 expression by hydrogen peroxide and leaf water status during excess light stress reveals a functional organization of Arabidopsis leaves. Plant J., 33: 691-705.
PubMedDirect Link - Guan, L.M., J. Zhao and J.G. Scandalios, 2000. Cis-elements and trans-factors that regulate expression of the maize Cat1 antioxidant gene in response to ABA and osmotic stress: H2O2 is the likely intermediary signaling molecule for the response. Plant J., 22: 87-95.
CrossRef - Hou, Y.D., Z.F. Guo, Y. Yi, H.N. Li and H.G. Li et al., 2010. Effects of cold acclimation and exogenous pytohormone abscisic acid treatment on physiological indicators of winterness wheat. J. Plant Sci., 5: 125-136.
CrossRefDirect Link - Hu, X., M. Jiang, A. Zhang and J. Lu, 2005. Abscisic acid-induced apoplastic H2O2 accumulation up-regulates the activities of chloroplastic and cytosolic antioxidant enzymes in maize leaves. Planta, 223: 57-68.
CrossRefDirect Link - Jiang, M. and J. Zhang, 2001. Effect of abscisic acid on active oxygen species, antioxidative defence syste m and oxidative damage in leaves of maize seedlings. Plant Cell Physiol., 42: 1265-1273.
CrossRefDirect Link - Khorshidi, M. and A.M. Nojavan, 2006. The effects of abscisic acid and CaCl2 on the activities of antioxidant enzymes under cold stress in maize seedlings in the dark. Pak. J. Biol. Sci., 9: 54-59.
CrossRefDirect Link - Kumari, S., 2010. Cellular changes and their relationship to morphology, abscisic acid accumulation and yield in wheat (Triticum aestivum) cultivars under water stress. Am. J. Plant Physiol., 5: 257-277.
CrossRefDirect Link - Laloi, C., D. Mestres-Ortega, Y. Marco, Y. Meyer and J.P. Reichheld, 2004. The Arabidopsis cytosolic h5 gene induction by oxidative stress and its w-box-mediated response to pathogen elicitor. Plant Physiol., 134: 1006-1016.
CrossRef - Nakano, Y. and K. Asada, 1981. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in Spinach chloroplasts. Plant Cell Physiol., 22: 867-880.
CrossRefDirect Link - Orozco-Cardenas, M.L., J. Narvaez-Vasquez and C.A. Ryan, 2001. Hydrogen peroxide acts as a second messenger for the induction of defense genes in tomato plants in response to woundingsystemin and methyl jasmonate. Plant Cell, 13: 179-191.
PubMed - Park, S.Y., S.H. Ryu, I.C. Jang, S.Y. Kwon , J.G. Kim, S.S. Kwak, 2004. Molecular cloning of a cytosolic ascorbate peroxidase cDNA from cell cultures of sweetpotato and its expression in response to stress. Mol. Genet Genomics, 271: 339-346.
CrossRef - Pei, Z.M., Y. Murata, G. Benning, S. Thomine and B. Klusener et al., 2000. Calcium channels activated by hydrogen peroxide mediate abscisic acid signalling in guard cells. Nature, 406: 731-734.
CrossRefDirect Link - Saleh, A.A.H., 2007. Amelioration of chilling injuries in mung bean (Vigna radiata L.) seedlings by paclobutrazol, abscisic acid and hydrogen peroxide. Am. J. Plant Physiol., 2: 318-332.
CrossRefDirect Link - Shaedle, M. and J.A. Bassham, 1977. Chloroplast glutathione reductase. Plant Physiol., 59: 1011-1012.
Direct Link - Sivaci, A., E. Elmas and F. Gumus, 2008. Changes in abscisic acid contents of some aquatic plants exposed to cadmium and salinity. Int. J. Bot., 4: 104-108.
CrossRefDirect Link