
Baby Joseph
Interdisciplinary Research Unit, Department of Biotechnology, Malankara Catholic College, Mariagiri, Kaliakkavilai Post, Kanyakumari District, Tamil Nadu, 629153, India
D. Jini
Interdisciplinary Research Unit, Department of Biotechnology, Malankara Catholic College, Mariagiri, Kaliakkavilai Post, Kanyakumari District, Tamil Nadu, 629153, India
International Journal of Botany
Year: 2010 | Volume: 6 | Issue: 4 | Page No.: 456-464







ABSTRACT
Salinity toxicity is a worldwide agricultural and eco-environmental problem. Salt stress is one of the major abiotic stresses faced by plants, which adversely affect their productivity. Salt stress causes reduction of crop yield and alterations in plant metabolism, including a reduced water potential, ion imbalances and toxicity and sometimes severe salt stress may even threaten survival. Salinity also leads to oxidative stress in plants due to the production of Reactive Oxygen Species (ROS) such as the super oxide radical, hydrogen peroxide and hydroxyl radical. Oxidative stress is one of the major limiting factors in plant productivity. Reactive Oxygen Species (ROS) generated during metabolic processes damage cellular functions and consequently lead to disease, senescence and cell death. Plants have evolved an efficient defense system by which the ROS is scavenged by antioxidant enzymes such as superoxide dismutase (SOD), calatase (CAT), peroxidase (POX), Polyphenoloxidase (PPO) and Glutathione Reductase (GR). Attempts to reduce oxidative damages under the salt stress conditions have included the manipulation of ROS scavenging enzymes by gene transfer technology. It is important to maintain and/or increase the productivity (photosynthetic capacity) under stressful environment by developing plants that have well adapted to environmental stress through manipulating antioxidant system. In this study, we discuss the role of antioxidant enzymes for salt tolerance in plants and this study will help to improve the tolerability of plants to salt by enhancing the expression of antioxidant enzymes.
PDF Abstract XML References Citation
Received: August 13, 2010;
Accepted: August 21, 2010;
Published: November 27, 2010
How to cite this article
Baby Joseph and D. Jini, 2010. Insight into the Role of Antioxidant Enzymes for Salt Tolerance in Plants. International Journal of Botany, 6: 456-464.
DOI: 10.3923/ijb.2010.456.464
URL: https://scialert.net/abstract/?doi=ijb.2010.456.464
DOI: 10.3923/ijb.2010.456.464
URL: https://scialert.net/abstract/?doi=ijb.2010.456.464
INTRODUCTION
Many arid and semi-arid regions in the world contain soils and water resources that are too saline for most of the common economic crops, which affect plants through osmotic effects, ion specific effects and oxidative stress (Munns, 2002; Pitman and Lauchli, 2002). High concentrations of salt in soil are causing large decreases in yields for a wide variety of crops all over the world (Sekmen et al., 2007). Currently, 20% of the world's cultivated land is affected by salinity, which results in the loss of 50% of agricultural yield (Zhu, 2001; Bartels and Sunkar, 2005). Many crop plants such as barley, maize and rice, are often subject to salinity stress (Sairam and Tyagi, 2004).
Abiotic pressures like salt stress and chemical insultance can impose limitations on crop productivity and also limit land available for farming, often in regions that can ill afford such constraints, thus highlighting a greater need for understanding how plants respond to adverse conditions with the hope of improving tolerance of plants to environmental stress (Joseph et al., 2010). The effects of various environmental stresses in plants are known to be mediated, at least in part, by an enhanced generation of Reactive Oxygen Species (ROS) including ·O2¯, H2O2 and ·OH¯ (Hernandez et al., 2000; Benavides et al., 2000). These ROS are highly reactive and can alter normal cellular metabolism through oxidative damage to membranes, proteins and nucleic acids; they also cause lipid peroxidation, protein denaturation and DNA mutation (Imlay, 2003). To prevent damage to cellular components by ROS, plants have developed a complex antioxidant system. The primary components of this system include carotenoids, ascorbate, glutathione and tocopherols, in addition to enzymes such as superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPX), peroxidases and the enzymes involved in ascorbate-glutathione cycle (Foyer and Halliwell, 1976), such as ascorbate peroxidase (APX) and glutathione reductase (GR). Many components of this antioxidant defense system can be found in various subcellular compartments (Hernandez et al., 2000).
A comparison of the antioxidant responses of plant genotypes exhibiting differential tolerance to salt stresscould contribute to the understanding of the physiological and biochemical mechanisms of protection against salt-induced oxidative damage. Therefore, the present study was conducted to investigate the role of the antioxidant-enzymatic system in relation to salt tolerance in plants.
Reactive oxygen species: Reactive Oxygen Species (ROS) are regarded as the main source of damage to cells under biotic and abiotic stresses (Candan and Tarhan, 2003; Gara at al., 2003; Mittler, 2002; Vaidyanathan et al., 2003). ROS’s are partially reduced forms of atmospheric oxygen, which are produced in vital processes such as photorespiration, photosynthesis and respiration (Mittler, 2002; Uchida et al., 2002). To produce water in these processes, four electrons are required for perfect reduction of oxygen. But ROS typically results from the transference of one, two and three electrons, respectively, to O2 to form superoxide (·O2¯), peroxide hydrogen (H2O2) and hydroxyl radical (·OH¯) (Mittler, 2002). These species of oxygen are highly cytotoxic and can seriously react with vital biomolecules such as lipids, proteins, nucleic acid, etc., causing lipid peroxidation, protein denaturing and DNA mutation, respectively (Breusegem et al., 2001; Scandalios, 1993; Quiles and Lopez, 2004). Evidence suggests that membranes are the primary sites of salinity injury to cells and organelles (Candan and Tarhan, 2003) because ROS can react with unsaturated fatty acids to cause peroxidation of essential membrane lipids in plasmalemma or intracellular organelles (Karabal et al., 2003; Stewart and Bewley, 1980).
Oxidative damage by salt stress: Oxygen is essential for the existence of aerobic life, but toxic reactive oxygen species (ROS), which include the superoxide anion radical (•O2¯), hydroxyl radical (•OH¯) and hydrogen peroxide (H2O2), are generated in all aerobic cells during metabolic processes (Foyer et al., 1994; Asada, 1999). Injury caused by these ROS is known as oxidative stress, which is one of the major damaging factors to plants exposed to environmental stress. It is well documented that abiotic stresses exert at least in part of their effects by causing oxidative damage (Smirnoff, 1995).
Oxidative damages are caused by Reactive Oxygen Species (ROS) and excess amounts of ROS are harmful to many cellular components, including membrane lipids. ROS cause peroxidation of polyunsaturated fatty acids in the membranes (Smirnoff, 2005). Production of ROS is increased under saline conditions (Greenway and Munns, 1980; Hasegawa et al., 2000) and ROS-mediated membrane damage has been demonstrated to be a major cause of the cellular toxicity by salinity in rice, tomato and citrus (Gueta-Dahan et al., 1997; Dionisio-Sese and Tobita, 1998; Mittova et al., 2004). The effects of various environmental stresses in plants are known to be mediated, at least in part, by an enhanced generation of reactive oxygen species (ROS) including ·O2¯, H2O2 and ·OH¯ (Hernandez et al., 2000; Benavides et al., 2000).
Regulation of Reactive Oxygen Species (ROS) at normal environment: Under normal growth conditions, low amounts of ROS such as superoxide radical (·O2¯), hydrogen peroxide (H2O2), hydroxyl radical (·OH¯) and singlet oxygen (1O2) are metabolic byproducts of plant cells (Cai-Hong et al., 2005). Plants have developed the scavenging mechanism of ROS categorized as enzymatic and non-enzymatic (Reddy et al., 2004; Demiral and Turkan, 2005). When ROS increases, chain reactions start, in which superoxide dismutase (SOD) catalyzes the dismutation of O2¯ radicals to molecular O2 and H2O2 (Meloni et al., 2003). The H2O2 is then detoxified in the ascorbate-glutathione cycle (Asada, 1999; Mittler, 2002), which involves the oxidation and re-reduction of ascorbate and glutathione through the ascorbate peroxidase (APX) and Glutathione Reductase (GR) action (Noctor and Foyer, 1998). If there is a serious imbalance in any cell compartment between the production of Reactive Oxygen Species (ROS) and antioxidant defense, oxidative stress and damage occurs (Mittler, 2002). When a plant faces harsh conditions, ROS production will overcome scavenging systems and oxidative stress will burst. In these conditions, ROS attack vital biomolecules and disturb the cell metabolism and ultimately the cell causes its own death (Sakihama et al., 2002).
Scavenging of reactive oxygen species by plants under salt stress condition: Salt stress induces cellular accumulation of ROS which can damage membrane lipids, proteins and nucleic acids (Hernandez et al., 1993, 1999; 2000; Mansour et al., 2005; Ben-Amor et al., 2007; Eyidogan and Oz, 2007). Much of the injury to plants exposed to stress is connected with oxidative damage at the cellular level (Foyer and Noctor, 2003). During oxidative stress, the excess production of ROS causes membrane damage that eventually leads to cell death. For protection against ROS, plants contain antioxidant enzymes as well as a wide array of non-enzymatic antioxidants (Blokhina et al., 2003).
Fortunately, plants have developed various protective mechanisms to eliminate or reduce ROS, which are effective at different levels of stress-induced deterioration (Beak and Skinner, 2003). The enzymatic antioxidant system is one of the protective mechanisms including superoxide dismutase (SOD: EC 1.15.1.1), which can be found in various cell compartments and it catalyses the disproportion of two ·O2¯ radicals to H2O2 and O2 (Scandalios, 1993). H2O2 is eliminated by various antioxidant enzymes such as catalases (CAT: EC 1.11.1.6) (Kono and Fridovich, 1983; Scandalios, 1993) and peroxidases (POX: EC 1.11.1.7) (Gara et al., 2003; Jablonski and Anderson, 1982) which convert H2O2 to water. Other enzymes that are very important in the ROS scavenging system and function in the ascorbate-glutathione cycle are glutathione reductase (GR: EC 1.6.4.2), monodehydro ascorbate reductase (MDHAR: EC 1.6.5.4) and dehydroascorbate reductase (DHAR: EC 1.8.5.1) (Candan and Tarhan, 2003; Yoshimura et al., 2000).
Role of antioxidant enzymes for salt tolerance in plants: To minimize the effects of oxidative stress, plant cells have evolved a complex antioxidant system, which is composed of low-molecular mass antioxidants (glutathione, ascorbate and carotenoids) as well as ROS-scavenging enzymes, such as: superoxide dismutase (SOD), calatase (CAT), ascorbate peroxidase (APX), guaiacol peroxidase (GPX) and glutathione reductase (GR) (Apel and Hirt, 2004).
One of the well-known mechanisms, that how the antioxidants work properly at the onset of oxidative stress is the water-water cycle (Asada, 1999; Fig. 1). The most important function of this cycle is a rapid, immediate scavenging of superoxide anion radical and hydrogen peroxide at the site of generation prior to their interaction with target molecules. SOD, APX (thylakoid-bound and stroma), monodehydro ascorbate reductase (MDHAR), dehydroascorbate reductase (DHAR) and Glutathione Reductase (GR) were participated in this cycle. However, this antioxidative mechanism seems to be not enough to protect plants from the elevated environmental stresses. To maintain the productivity of plants under the stress condition, it is important to fortify the antioxidative mechanism of the chloroplasts by manipulating the antioxidant enzymes and small antioxidant molecules in the chloroplast (Kwon et al., 2001).
Kartashov et al. (2008) studied the activities of principal antioxidant enzymes and the accumulation of sodium ions and proline. Moreover, plants with the high activity of peroxidase and active proline accumulation could acclimate to salts stress (100 mM NaCl, 24 h) independently of SOD activity (Kartashov et al., 2008). Several studies have pointed out that salt-tolerant species increased their antioxidant enzyme activities and antioxidant contents in response to salt treatment, whereas salt-sensitive species failed to do so (Shalata et al., 2001; Demiral and Turkan, 2005). The scavenging of ROS by increased activation of antioxidant enzymes can improve salt tolerance (Alscher et al., 2002). A relationship between salt tolerance and increased activation of antioxidant enzymes has been demonstrated in Plantago (Sekmen et al., 2007), pea (Hernandez et al., 2000), rice (Dionisio-Sese and Tobita, 1998), tomato, maize (Neto et al., 2005), sorghum (Costa et al., 2005; Heidari, 2009), soybean (Cicek and Cakirlar, 2008) and mulberry (Harinasut et al., 2003).
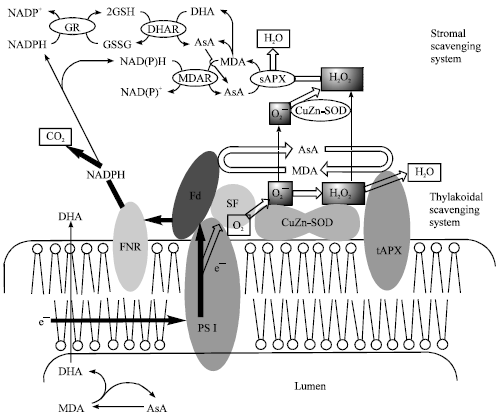 | |
| Fig. 1: | The water-water cycle and microcompartmetalization of the participating enzymes (Asada, 1999) |
The generation of ROS and increased activity of many antioxidant enzymes during salt stress have been reported in cotton (Desingh and Kanagaraj, 2007), mulberry (Sudhakar et al., 2001; Harinasut et al., 2003), wheat (Sairam et al., 2002), tomato (Mittova et al., 2002), rice (Vaidyanathan et al., 2003) and sugar beet (Bor et al., 2003). In general, the activities of antioxidant enzymes were increased in the root and shoot under saline stress. But the increase was more significant and consistent in the root (Kim et al., 2005). The activities of antioxidant enzymes were increased in the root and shoot under saline stress in barley seedlings treated with 200 mM NaCl (Kim et al., 2005).
The quantitative and qualitative aspects of changes are often related to the levels of resistance to salinity. In tomato and citrus, salt-tolerance is attributed to the increased activities of SOD, APX and CAT (Gueta-Dahan et al., 1997; Mittova et al., 2004). Further supporting evidence on the involvement of antioxidant enzymes in salt tolerance has been provided by transgenic plants with a reduced or an increased expression of antioxidant enzymes (Willekens et al., 1997). Increased protection to salt stress has been demonstrated by the over expression of cytosolic APX (Torsethaugen et al., 1997). Jaleel (2009) carried out an experiment with different concentrations of sodium chloride (NaCl) in alba and rosea varieties of Catharanthus roseus (L.) to determine the changes that occurring in antioxidant enzymes in various plant parts and reported that, C. roseus is an ideal plant for cultivation in salt affected areas in order to obtain plants with high antioxidant and medicinal values.
Superoxide Dismutase (SOD): The Superoxide Dismutase (SOD) (EC 1.15.1.1) is a metalloprotein that catalyzes the initial step in the water-water cycle in chloroplasts, the dismutation of superoxide to H2O2 and molecular oxygen (Scandalios, 1993; Bowler et al., 1991; Allen et al., 1997; Asada, 1999). The subsequent reduction of H2O2 to water through the cycle in the chloroplast uses reducing equivalents from NADPH (Foyer et al., 1994). The increased SOD activities in leaves of Ulmus pumila presented an increased trend at salt concentrations less than 1.5%, the growth of seedlings did not decline and tress and leaves had no symptom of injury, while the salt concentrations exceeded 1.5%, SOD activities sharply decreased (Song et al., 2006). In rice, the salt-tolerant varieties have higher SOD activity and lower lipid peroxidation than the salt-sensitive varieties (Dionisio-Sese and Tobita, 1998). It was suggested that the ratio between superoxide dismutase and H2O2-scavenging enzyme activities could be used as a working hypothesis for a biochemical marker for salt tolerance in sorghum (Costa et al., 2005).
Catalase (CAT): Catalase is a common enzyme found in nearly all living organisms that are exposed to oxygen, where it functions to catalyze the decomposition of hydrogen peroxide to water and oxygen. It is a tetramer of four polypeptide chains, each over 500 amino acids long (Chelikani et al., 2004). Salt stress was associated with an increase of the antioxidant enzyme response and induction of new CAT isoforms in rice (Srivalli et al., 2003). Salt-tolerant cotton cultivars exhibited significantly greater in CAT, APX and SOD activities as compared to the salt-sensitive ones (Gossett et al., 1994). Nagamiya et al. (2007) introduced katE, a catalase gene of Escherichia coli, into japonica rice cultivar. Catalase activity in the transgenic rice plants was 1.5 to 2.5 fold higher than non-transgenic rice plants.
Peroxidase (POX) and Polyphenol Oxidase (PPO): Meratan et al. (2008) studied the effects of salinity on antioxidant enzymes in three Acanthophyllum species. A. glandulosum showed a remarkable increase in POX and PPO between 50 to 200 mM NaCl. In A. sordidum, POX and PPO activities increased at 50 mM NaCl and then decreased at higher salinities. Their results showed that the differences in the antioxidant enzyme activities of seedling may, at least in part explain the greater tolerance of A. glandulosum comparing to A. sordidum and A. laxiusculum. Sreenivasulu et al. (1999) studied the effect of NaCl on total peroxidase activity, induction of isoperoxidases and lipid peroxidation in 5-day-old seedlings of two contrasting genotypes of Setaria italica L. Total peroxidase activity increased under NaCl salinity and the degree of elevation in the activity was salt concentration dependent.
Aspartate Peroxidase (APX): The H2O2 produced via the disproportionation of O2·G catalyzed with SOD is reduced to water by APX (E.C. 1.11.1.11), which use ascorbate as the electron donor in chloroplasts. APX is a heme peroxidase and uses two molecules of ascorbate to reduce H2O2 to water, with the generation of two molecules of monodehydroascorbate (MDHA). It distributed in at least four distinct cell compartments, the stroma (sAPX) and thylakoid membrane (tAPX) in chloroplasts, the microbody (mAPX) and the cytosol (cAPX) (Yoshimura et al., 2000). In addition, Zhang et al. (1997) identified an APX associated with the glyoxysomal membranes. Allen et al. (1997) reported that transgenic tobacco plants expressing gene constructs for either cytosolic APX or a chimeric chloroplast-targeted cytosolic APX from pea have the increased protection from photo oxidative stress.
Role of antioxidant enzymes in rice plants: Rice is one of the most important crops all over the world. But the rice agriculture is often threatened by abiotic stresses, including salinity. Many researches have focused on this problem, with the aim to find the mechanism of salt tolerance and the critical elements that may confer the tolerance to salinity. Some studies have shown that salt stress arouses many aspects of protective reaction, such as accumulation of small osmotic molecules, enhanced selective ion transport, increased reactive oxygen species (Csonka and Hanson, 1991). The responses of the antioxidant enzymes in the resistance to salt stress were determined in the rice (Oryza sativaL. cv. Dongjin) plant. In the leaves of the rice plant, salt stress preferentially enhanced the content of H2O2 as well as the activities of the superoxide dismutase (SOD), ascorbate peroxidase (APX) and peroxidase specific to guaiacol, whereas it induced the decrease of catalase activity (Lee et al., 2001). The superoxide dismutase activity in roots and glutathione reductase activity in both shoots and roots were decreased significantly under high salinity levels in rice plant (Kumar and Shriram, 2009).
Strategies to improve salt stress tolerance: Recent advances in plant genome mapping and molecular biology techniques offer a new opportunity for understanding the genetics of salt stress-resistance genes and their contribution to plant performance under salt stress. These biotechnological advances will provide new tools for breeding in salt stress environment. Molecular genetic maps have been developed for major crop plants, including rice, wheat, maize, barley, sorghum and potato, which make it possible for scientists to tag desirable traits using known DNA landmarks. Molecular genetic markers allow breeders to track genetic loci controlling stress resistance without having to measure the phenotype, thus reducing the need for extensive field-testing over time and space. Moreover, gene pyramiding or introgression can be done more precisely using molecular tags. Together, molecular genetic markers offer a new strategy known as marker assisted selection. Another molecular strategy which depends on gene cloning and plant transformation technology, is genetic engineering of selected genes into elite breeding lines. What makes a particular goal attainable or unattainable in genetic engineering experiments is the availability of the following three inputs: (1) the gene of interest, (2) an effective technique for transferring the desired gene from one species to another and (3) promoter sequences for regulated expression of that gene. Amongst these, the first is considered a rate-limiting factor. Arrays of salt stress-induced genes have been isolated. Salt stress-responsive genes can be analysed following targeted or non-targetted strategy. The targetted approach relies upon the availability of relevant biochemical information (i.e., in terms of defined enzyme, protein, a biochemical reaction or a physiological phenomenon). The non-targetted strategy to obtain a desired gene is indirect. This strategy, for instance, includes differential hybridization and shotgun cloning. The list of genes, whose transcription is upregulated in response to stress, is rapidly increasing. Understanding of the mechanisms which regulate gene expression and the ability to transfer genes from other organisms into plants will expand the ways in which plants can be utilized. To exploit the full potential of these approaches, it is essential that the knowledge is applied to agriculturally and ecologically important plant species.
CONCLUSION AND FUTURE PROSPECTS
Salinity in the soil and irrigation water is an environmental problem and a major constraint for crop production. Salt stress (NaCl) has both osmotic (cell dehydration) and toxic (ion accumulation) effects on plants. Although, a wide range of genetic adaptations to saline conditions has been observed and a number of significant physiological responses have been associated with tolerance, underlying mechanisms of salt tolerance in plants are still poorly understood.
The ROS, especially hydrogen peroxide, have been proved as a central component of plant adaptation to biotic and abiotic stresses (Mittler, 2002; Karpinski et al., 1999). Under the stress conditions, ROS may play in damaging the cellular components or signaling the activation of defense responses. To allow for these different roles, cellular levels of ROS must be tightly controlled. In first, precise understanding the roles of each ROS scavenging enzyme and small molecular antioxidants in stress adaptation and accurate characterization of the complex stress tolerance phenotypes is necessary to develop stress tolerant plants. However, this antioxidative mechanism seems to be not enough to protect plants from the elevated environmental stresses such as salinity. To maintain the productivity of plants under the salt stress condition, it is important to fortify the antioxidative mechanism of the chloroplasts by manipulating the antioxidant enzymes and small antioxidant molecules in the chloroplast.
Cloned plant genes and transgenic plants have become a standard tool in plant-stress biology. These technologies have mainly been applied to model systems and have greatly enlarged the knowledge of mechanisms of tolerance. The various abiotic stresses cause changes in plant processes at all levels of organization (morphological, physiological, biochemical and molecular). In recent years, attention has focused on alterations in gene expression. The list of genes whose transcription is upregulated in response to stress is rapidly increasing. Functions for some of these polypeptides are close to being identified and their likely role in stress physiology is being determined. The understanding of mechanisms that regulate gene expression and the ability to transfer genes from other organisms into plants will expand the ways in which plants can be utilized. The exploitation of cloned genes to alter the function of gene products in transgenic plants provides novel opportunities to assess their biological role in a salt stress response.
ACKNOWLEDGMENTS
Authors gratefully acknowledged to our Malankara Catholic College Correspondent Fr. Prem Kumar (M.S.W) given encouragement and support for preparation of this research manuscript. We wish to express the sincere thanks to Dr. Sujatha (ICBM), Mr. K. Suresh and the research scholars in the Interdisciplinary Research centre for updated article collection and all the efficient supports of this review preparation.
REFERENCES
- Allen, R.D., R.P. Webb and S.A. Schake, 1997. Use of transgenic plants to study antioxidant defenses. Free Radic. Biol. Med., 23: 473-479.
CrossRefDirect Link - Apel, K. and H. Hirt, 2004. Reactive oxygen species: Metabolism, oxidative stress and signal transduction. Annu. Rev. Plant Biol., 55: 373-399.
CrossRefPubMedDirect Link - Asada, K., 1999. The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Physiol. Plant Mol. Biol., 50: 601-639.
CrossRefPubMedDirect Link - Bartels, D. and R. Sunkar, 2005. Drought and salt tolerance in plants. Crit. Rev. Plant Sci., 24: 23-58.
CrossRefDirect Link - Beak, K.H. and D.Z. Skinner, 2003. Alteration of antioxidant enzyme gene expression during cold acclimation of near-isogenic wheat lines. Plant Sci., 165: 1221-1227.
CrossRefDirect Link - Benavides, M.P., P.L. Marconey, S.M. Gallego, M.E. Comba and M.I. Tamaro, 2000. Relationship between antioxidant defence systems and salt tolerance in Solano tobarasum. Aust. J. Plant Physiol., 27: 273-278.
Direct Link - Blokhina, O., E. Virolainen and K.V. Fagerstedt, 2003. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot., 91: 179-194.
CrossRefPubMedDirect Link - Bor, M., F. Ozdemir and I. Turkan, 2003. The effect of salt stress on lipid peroxidation and antioxidants in leaves of sugar beet Beta vulgaris L. and wild beet Beta maritima L. Plant Sci., 164: 77-84.
CrossRefDirect Link - Bowler, C., L. Slooten, S. Vandenbranden, R. de Rycke and J. Botterman et al., 1991. Manganese superoxide dismutase can reduce cellular damage mediated by oxygen radicals in transgenic plants. EMBO J., 10: 1723-1732.
Direct Link - Van Breusegem, F., E. Vranova, J.F. Dat and D. Inze, 2001. The role of active oxygen species in plant signal transduction. Plant Sci., 161: 405-414.
CrossRefDirect Link - Candan, N. and L. Tarhan, 2003. The correlation between antioxidant enzyme activities and lipid peroxidation levels in Mentha pulegium organs grown in Ca2+, Mg2+, Cu2+, Zn2+ and Mn2+ stress conditions. Plant Sci., 163: 769-779.
CrossRefDirect Link - Chelikani, P., I. Fita and P.C. Loewen, 2004. Diversity of structures and properties among catalases. Cell. Mol. Life Sci., 61: 192-208.
CrossRefDirect Link - Cicek, N. and H. Cakirlar, 2008. Changes in some antioxidant enzyme ectivities in six soybean cultivars in response to long-term salinity at two different temperatures. Gen. Applied Plant Physiol., 34: 267-280.
Direct Link - Costa, P.H.A., A. Neto, M. Bezerra, J. Prisco and E. Filho, 2005. Antioxidant-enzymatic system of two sorghum genotypes differing in salt tolerance. Braz. J. Plant Physiol., 17: 353-361.
Direct Link - Csonka, L.N. and A.D. Hanson, 1991. Prokaryotic osmoregulation: Genetics and physiology. Annu. Rev. Microbiol., 45: 569-606.
Direct Link - Demiral, T. and I. Turkan, 2005. Comparative lipid peroxidation, antioxidant defense systems and proline content in roots of two rice cultivars differing in salt tolerance. Environ. Exp. Bot., 53: 247-257.
CrossRefDirect Link - Desingh, R. and G. Kanagaraj, 2007. Influence of salinity stress on photosynthesis and antioxidative systems in two cotton varities. Genet. Appl. Plant Physiol., 33: 221-234.
Direct Link - Dionisio-Sese, M.L. and S. Tobita, 1998. Antioxidant responses of rice seedlings to salinity stress. Plant Sci., 135: 1-9.
CrossRefDirect Link - Eyidogan, F. and M.T. Oz, 2007. Effect of salinity on antioxidant responses of chickpea seedlings. Acta Physiol. Plant, 29: 485-493.
CrossRef - Foyer, C.H. and B. Halliwell, 1976. The presence of glutathione and glutathione reductase in chloroplasts: A proposed role in ascorbic acid metabolism. Planta, 133: 21-25.
CrossRefPubMedDirect Link - Foyer, C.H., P. Descourvieres and K.J. Kunert, 1994. Protection against oxygen radicals: An important defence mechanism studied in transgenic plants. Plant Cell Environ., 17: 507-523.
CrossRefDirect Link - Foyer, C.H. and G. Noctor, 2003. Redox sensing and signalling associated with reactive oxygen in chloroplasts, peroxisomes and mitochondria. Physiol. Plant., 119: 355-364.
CrossRefDirect Link - De Gara, L., M.C. de Pinto and F. Tommasi, 2003. The antioxidant systems vis-a-vis reactive oxygen species during plant-pathogen interaction. Plant Physiol. Biochem., 41: 863-870.
CrossRefDirect Link - Greenway, H. and R. Munns, 1980. Mechanisms of salt tolerance in nonhalophytes. Annu. Rev. Plant Physiol., 31: 149-190.
CrossRefDirect Link - Gueta-Dahan, Y., Z. Yaniv, B.A. Zilinskas and G. Ben-Hayyim, 1997. Salt and oxidative stress: Similar and specific responses and their relation to salt tolerance in citrus. Planta, 204: 460-469.
CrossRef - Harinasut, P., D. Poonsopa, K. Roengmongkol and R. Charoensataporn, 2003. Salinity effects on antioxidant enzymes in mulberry cultivar. Sci. Asia, 29: 109-113.
Direct Link - Hasegawa, P.M., R.A. Bressan, J.K. Zhu and H.J. Bohnert, 2000. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol., 51: 463-499.
CrossRefDirect Link - Heidari, M., 2009. Antioxidant activity and osmolyte concentration of sorghum (Sorghum bicolor) and wheat (Triticum aestivum) genotypes under salinity stress. Asian J. Plant Sci., 8: 240-244.
CrossRefDirect Link - Hernandez, J.A., A. Jimenez, P. Mullineaux and F. Sevilia, 2000. Tolerance of pea (Pisum sativum L.) to long-term salt stress is associated with induction of antioxidant defences. Plant Cell Environ., 23: 853-862.
CrossRefDirect Link - Hernandez, J.A., A. Campillo, A. Jimenez, J.J. Alarcon and F. Sevilla, 1999. Response of antioxidant systems and leaf water relations to NaCl stress in pea plants. New Phytol., 141: 241-251.
CrossRefDirect Link - Hernandez, J.A., F.J. Corpas, M. Gomez, L.A. Rio and F. Sevilla, 1993. Salt-induced oxidative stress mediated by activated oxygen species in pea leaf mitochondria. Physiol. Plant., 89: 103-110.
CrossRefDirect Link - Jablonski, P.P. and J.W. Anderson, 1982. Light-dependent reduction of hydrogen peroxide by ruptured pea chloroplasts. Plant Physiol., 69: 1403-1407.
Direct Link - Karabal, E., M. Yucel and H.A. Oktem, 2003. Antioxidant responses of tolerant and sensitive barley cultivars to boron toxicity. Plant Sci., 164: 925-933.
CrossRefDirect Link - Karpinski, S., H. Reynolds, B. Karpinska, G. Wingsle, G. Creissen and P. Mullineaux, 1999. Systemic signaling and acclimation in response to excess excitation energy in Arabidopsis. Science, 284: 654-657.
CrossRefDirect Link - Kartashov, A.V., N.L. Radyukina, Y.V. Ivanov, P.P. Pashkovskii, N.I. Shevyakova and V.V. Kuznetsov, 2008. Role of antioxidant systems in wild plant adaptation to salt stress. Russ. J. Plant Physiol., 55: 463-468.
CrossRef - Kim, S., J. Lim, M. Park, Y. Kim, T. Park, Y. Seo and S. Yun, 2005. Enhanced antioxidant enzymes are associated with reduced hydrogen peroxide in barley roots under saline stress. J. Biochem. Mol. Biol., 38: 218-224.
Direct Link - Kono, Y. and I. Fridovich, 1983. Inhibition and reactivation of Mn-catalase. Implications for valence changes at the active site manganese. J. Biol. Chem., 258: 13646-13668.
PubMedDirect Link - Kumar, V. and V. Shriram, 2009. Antioxidant enzyme activities and protein profiling under salt stress in indica rice genotypes differing in salt tolerance. Arch. Agrono. Soil Sci., 55: 379-394.
CrossRef - Lee, D.H., Y.S. Kim and C.B. Lee, 2001. The inductive responses of the antioxidant enzymes by salt stress in the rice (Oryza sativa L.). J. Plant Physiol., 158: 737-745.
CrossRef - Mansour, M.M.F., K.H.A. Salama, F.Z.M. Ali and A.F. Abou Hadid, 2005. Cell and plant responses to NaCl in Zea mays L. cultivars differing in salt tolerance. Gen. Applied Plant Physiol., 31: 29-41.
Direct Link - Meloni, D.A., M.A. Oliva, C.A. Martinez and J. Cambraia, 2003. Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress. Environ. Exp. Bot., 49: 69-76.
CrossRefDirect Link - Meratan, A.A., S.M. Ghaffari and V. Niknam, 2008. Effects of salinity on growth, proteins and antioxidant enzymes in three Acanthophyllum species of different ploidy levels. JSUT, 33: 1-8.
Direct Link - Mittler, R., 2002. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci., 7: 405-410.
CrossRefPubMedDirect Link - Mittova, V., M. Tal, M. Volokita and M. Guy, 2002. Salt stress induces up-regulation of an efficient chloroplasts antioxidant system in the salt-tolerant wild tomato species Lycopersicon pennellii but not in the cultivated species. Plant Physiol., 115: 393-400.
Direct Link - Mittova, V., M. Guy, M. Tal and M. Volokita, 2004. Salinity upregulates the antioxidative system in root mitochondria and peroxisomes of the wild salt-tolerant tomato species Lycopersicon pennellii. J. Exp. Bot., 55: 1105-1113.
Direct Link - Munns, R., 2002. Comparative physiology of salt and water stress. Plant Cell Environ., 25: 239-250.
CrossRefDirect Link - Noctor, G. and C.H. Foyer, 1998. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Mol. Biol., 49: 249-279.
CrossRefPubMedDirect Link - Quiles, M.J. and N.I. Lopez, 2004. Photoinhibition of photosystems I and II induced by exposure to high light intensity during oat plant grown effects on the chloroplastic NADH dehydrogenase complex. Plant Sci., 166: 815-823.
CrossRef - Reddy, A.R., K.V. Chaitanya and M. Vivekanandan, 2004. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol., 161: 1189-1202.
CrossRefPubMedDirect Link - Sairam, R.K., K.V. Roa and G.C. Srivastava, 2002. Differential response of wheat genotypes to long term salinity stress in relation to oxidative stress, antioxidant activity and osmolyte concentration. Plant Sci., 163: 1037-1046.
CrossRefDirect Link - Sairam, R.K. and A. Tyagi, 2004. Physiology and molecular biology of salinity stress tolerance in plants. Curr. Sci., 86: 407-421.
Direct Link - Sakihama, Y.M., F. Cohen, S.C. Grace and H. Yamasaki, 2002. Plant phenolic antioxidant and prooxidant activities: Phenolics-induced oxidative damage mediated by metals in plants. Toxicology, 177: 67-80.
CrossRefDirect Link - Scandalios, J.G., 1993. Oxygen stress and superoxide dismutases. Plant Physiol., 101: 7-12.
CrossRefPubMedDirect Link - Sekmen, A.H., I. Turkana, and S. Takiob, 2007. Differential responses of antioxidative enzymes and lipid peroxidation to salt stress in salt-tolerant Plantago maritime and salt-sensitive Plantago media. Physiol. Plant, 131: 399-411.
CrossRef - Shalata, A., V. Mittova, M. Volokita, M. Guy and M. Tal, 2001. Response of the cultivated tomato and its wild salt-tolerant relative Lycopersicon pennellii to salt-dependent oxidative stress: The root antioxidative system. Physiol. Plant., 112: 487-494.
CrossRefPubMedDirect Link - Song, F., C. Yang, X. Liu and G. Li, 2006. Effect of salt stress on activity of superoxide dismutase (SOD) in Ulmus pumila L. J. For. Res., 17: 13-16.
CrossRef - Srivalli, B., G. Sharma and R. Khanna-Chopra, 2003. Antioxidative defense system in an upland rice cultivar subjected to increasing intensity of water stress followed by recovery. Plant Physiol., 119: 503-512.
Direct Link - Stewart, R.R.C. and J.D. Bewley, 1980. Lipid peroxidation associated aging of soybean axes. Plant Physiol., 65: 245-248.
Direct Link - Sudhakar, C., A. Lakshmi and S. Giridarakumar, 2001. Changes in the antioxidant enzyme efficacy in two high yielding genotypes of mulberry (Morus alba L.) under NaCl salinity. Plant Sci., 161: 613-619.
CrossRefDirect Link - Uchida, A., A.T. Jagendorf, T. Hibino, T. Takabe and T. Takabe, 2002. Effects of hydrogen peroxide and nitric oxide on both salt and heat stress tolerance in rice. Plant Sci., 163: 515-523.
CrossRefDirect Link - Vaidyanathan, H., P. Sivakumar, R. Chakrabarty and G. Thomas, 2003. Scavenging of reactive oxygen species in NaCl-stressed rice (Oryza sativa L.)-differential response in salt-tolerant and sensitive varieties. Plant Sci., 165: 1411-1418.
CrossRefDirect Link - Yoshimura, K., Y. Yabute, T. Ishikawa and S. Shigeoka, 2000. Expression of spinach ascorbate peroxidase isoenzymes in response to oxidative stresses. Plant Physiol., 123: 223-233.
Direct Link - Zhang, H., J. Wang, U. Nickel, R.D. Allen and H.M. Goodman, 1997. Cloning and expression of an Arabidopsis gene coding a putative peroxisomal ascorbate peroxidase. Plant Mol. Biol., 34: 967-971.
CrossRef - De Azevedo-Neto, A.D., J.T. Pitsco, J. Eneas-Filho, C.E.B. de Abreu and E. Gomes-Filho, 2006. Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environ. Exp. Bot., 56: 87-94.
CrossRefDirect Link - Alscher, R.G., J.L. Donahue and C.L. Cramer, 1997. Reactive oxygen species and antioxidants: Relationship in green cells. Physiol. Plant., 100: 224-233.
CrossRefDirect Link
sajjad zare Reply
Hi Dear Baby
I had similar reaserch to your work. Your work is good.
we can relate enzyme activity with yield. i worked on canola.
this point is very important that Breeders could select resistance cultivars basis on enzyme activity in primary growth levels.
Best regard
elmor exclamador Reply
can i have a copy of your paper?