
E. U. Durugbo
Department of Biological Sciences, Redeemer`s University, Mowe, Ogun State, Nigeria
LiveDNA: 234.15316
O. T. Ogundipe
Department of Botany and Microbiology, University of Lagos, Akoka, Lagos, Nigeria
O. K. Ulu
Earth Sciences and Reservoir Services Division, Chevron Nigeria Limited, Nigeria
International Journal of Botany
Year: 2010 | Volume: 6 | Issue: 4 | Page No.: 351-370







ABSTRACT
Rich and well preserved assemblages of pollen, spores, and organic walled dinoflagellate cysts in 96 and 89 samples of Wells A and B from the Western Niger Delta Nigeria are recorded. The dominance of savanna pollen over wet climate indicators (mangrove, freshwater swamp species, brackish water swamp species and Palmae) and the preponderance of the dinocysts Polysphaeridium zoharyi and Operculodinium centrocarpum, species adapted to very saline and warm waters respectively, with abundant fungal spores dominated by Exesisporites sp., gives credence to a predominantly dry climate and lowered sea level during the Pliocene-Pleistocene (ca. 5.0-1.3 Ma) in the Gulf of Guinea. The most pronounced glacial events were around the 2.0-2.7 Ma, as well as between the 2.7-3.4 Ma.
PDF Abstract XML References Citation
Received: April 16, 2010;
Accepted: July 22, 2010;
Published: September 01, 2010
How to cite this article
E. U. Durugbo, O. T. Ogundipe and O. K. Ulu, 2010. Palynological Evidence of Pliocene-Pleistocene Climatic Variations from the Western Niger Delta, Nigeria. International Journal of Botany, 6: 351-370.
DOI: 10.3923/ijb.2010.351.370
URL: https://scialert.net/abstract/?doi=ijb.2010.351.370
DOI: 10.3923/ijb.2010.351.370
URL: https://scialert.net/abstract/?doi=ijb.2010.351.370
INTRODUCTION
Palynological studies in Nigeria are largely confined to the Tertiary Niger Delta, where numerous studies have been carried out following the discovery of petroleum resources in the Niger Delta in the late 1950s. Most of these reports exist as confidential reports of the oil and gas prospecting companies. The few published papers on the Niger Delta geological studies include those of Allen (1964a, b), Allen and Wells (1962), Short and Stauble (1967), Weber (1971), Weber and Daukoru (1975), Evamy et al. (1978), Doust and Omatsola (1990), Oboh et al. (1992), Haack et al. (2000). Furthermore, most palynological studies on the Niger Delta are concerned with zonation, age determinations and correlation. Only few had dealt with the vegetation vicissitudes and paleoenvironmental reconstruction for which palynological research is known for. Unfortunately, none had treated the global glacial records and the Plio-Pleistocene climatic variations of which much have been written in other parts of Africa and the world at large (Adam, 1993; Adam et al., 1989, 1990; Dupont and Welnelt, 1996; Dupont et al., 1998, 2000; Finch and Hill, 2008; Hooghiemstra, 1993; Hooghiemstra et al., 1986, 2006; Jahns et al., 1998; Lezine and Cazet, 2005; Leroy and Dupont, 1994, 1997; Weigelt et al., 2008; Molnar and Cane, 2004).
Since climatic changes do affect the vegetation, the reconstruction of the past vegetation will help us understand the mechanisms of such changes (Ivanor et al., 2007). Sowunmi (1987) had discussed the close relationship between the vegetation, soil and climate of an area. She opined that the flora of an area offers the clue to the major climatic regimes in such places. The most popular published works on the Niger Delta palynology are those of Sowunmi (1981a, b, 1986, 1987, 1999, 2002, 2004), Evamy et al. (1978), Biffi and Grignani (1983), Legoux (1978), Clarke and Fredriksen (1968), Knapp (1971), Jan du Chene and Salami (1978), Oboh and Salami (1989), Oboh, 1991a, b, 1993a, b, 1995), Oboh et al. (1992), Antolinez et al. (2006), Demchuk and Morley (2004), Ige (2009) and Adekanmbi and Ogundipe (2009). The modern Niger Delta appears, from sedimentological, faunal and floral evidence, to have a configuration today similar to those of the past. The physiography of the recent Niger Delta is governed by several factors, which influence transport, ultimate deposition of the sediment load and shape and growth of the Delta (Short and Stauble, 1967). Different workers have investigated climatic changes using palynomorphs in different parts of the world. The reports are scanty for Africa and the Gulf of Guinea to which the Niger Delta belongs.
Dupont et al. (1998) had used sporomorphs and dinoflagellate cysts in documenting the vegetation history of the West African forest during the last 700 ka in relation to changes in salinity and productivity of the eastern Gulf of Guinea from site GIK16867 in the northern Angola Basin. They found out that during most of the cool and cold periods, the Afromontane forest, rather than the open grass-rich dry forest, expanded to lower altitudes partly replacing the lowland rain forest of the borderlands east of the Gulf of Guinea.
Sowunmi (2004) had studied an 11 m terrestrial core from Ahanve in Badagry area of the coastal South-Western Nigeria as part of the Dahomey Gap project. The area was a Typha dominated freshwater swamp annually flooded by the Badagry Creek. She recognized 141 pollen and spore species out of which 104 were identified. She reported that the dominant vegetation communities that prevailed in the coastal South-Western Nigeria was made up of abundant forests, Rhizophora dominated mangrove forest in the tidal zone, freshwater swamp forest in the swampy regions under freshwater influence and drier, semi-deciduous forest on the well drained areas with some woody, savanna species and sparse grass cover at some intervals from ca. 8576±48 BP to sometime prior to ca. 3109±26 BP. Furthermore, between ca. 5780 and ca. 5682±32 BP, there was a marked decrease in all the three forest types and a corresponding marked increase in monolete fern spores Lygodium and Alchornea which she opined indicated a short-lived, drier climate. Again, just prior to ca. 3109±26 BP, Rhizophora declined sharply and later disappeared, it got replaced by freshwater swamp, an expanded freshwater swamp forest and coastal savannas made up of abundant grasses, Alchornea and Elaeis guineensis. The drier forest later became more opened leading to the development of derived savanna. This mosaic of more open forest and savannas established just prior to ca. 3109±26 BP she said constituted the eastern-most extension of the Dahomey Gap. She reasoned that from palynological evidence that from sometime after ca. 3109±26 BP, anthropogenic effects had caused the phenomenal rise in Elaeis guineensis pollen and concluded that though climatic and geomorphological factors appeared the main causes of the late Holocene vegetation changes, information from palynology and archeology indicated outstanding human impact.
Leroy and Dupont (1994) studied a 200 m long marine pollen record from ODP Site 658 (21°N, 19°W) which revealed cyclic fluctuations in vegetation and continental climate in northwestern Africa from 3.7 to 1.7 Ma. These cycles they found paralleled oxygen isotope stages. Prior to 3.5 Ma, the distribution of tropical forests and mangrove swamps reached Cape Blanc, 5°N of the present distribution. Between 3.5 and 2.6 Ma, forests occurred at this latitude during irregular intervals and nearly disappeared afterwards. Likewise, a Saharan paleoriver flowed continuously until isotope Stage 134 (3.35 Ma). When river discharge ceased, wind transport of pollen grains prevailed over fluvial transport. They observed that pollen indicators of trade winds gradually increased between 3.3 and 2.5 Ma. Again, strong aridification of the climate of northwestern Africa occurred during isotope Stage 130 (3.26 Ma). Humid conditions were reestablished followed by another aridification around 2.7 Ma. Afterwards repetitive latitudinal shifts of vegetation zones ranging from wooded savanna to desert flora dominated for the first time between 2.6 and 2.4 Ma as a response to the glacial stages 104, 100 and 98. They reported that as climatic conditions recorded in the Pliocene were not as dry as those of the middle and Late Pleistocene, latitudinal vegetation shifts near the end of the Pliocene resembled those of the interglacial-glacial cycles of the Brunhes chron.
Leroy and Dupont (1997) further carried out a high resolution pollen analysis on ODP site 658, off Cape Blanc, N-W Africa. They observed that the periods before 3.5 Ma were marked by warmer and wetter climates from northwestern Europe as far south as equatorial West Africa. They opined that the long term variation denoted the onset of a drier climate between 3.5 and 3.2 Ma. At 3.2 Ma, their pollen record revealed the aridification of Northwestern Africa probably in connection with enhanced Trade winds. After a period of re-established humidity, a second and stronger step to aridification started at ca. 2.6 Ma. They documented a reduction of the savannah vegetation from 2.8 Ma which they suggested could have resulted in the development of a desert in West Africa.
Jahns et al. (1998) had reconstructed the West African vegetation history for the last 400 ka from pollen analysis of a sediment core from the Atlantic Ocean off Liberia. They noted that an arid climate was indicated by the cold oxygen isotope stages 12, 10, 8, 6, 4, 3 and 2, which resulted in a southward shifting of the southern border of the savanna. They inferred that the Late Pleistocene glacial stages were more arid than those of the Middle Pleistocene. They recorded the persistence of the rain forest even during the glacial stages which they attributed to a glacial refuge of rain forest situated in the Guinean mountains.
Lezine and Cazet (2005) had carried out a high resolution pollen study of core KW31 off the mouth of Niger River (3° 31' 1N-05° 34'1E), from the Gulf of Guinea. The core was collected at a water depth of 1181m close to the coast and it revealed vegetation changes between 40,000 and 3500 cal yr B.P. in the West African lowlands. The microflora was highly diverse from the studied 69 samples with the counts ranging between 124 and 1026, excluding the mangrove pollen Rhizophora. Fern and herbaceous pollen grains of Poaceae and Cyperaceae dominated the assemblages. The percentages reaching 46, 76 and 27%, respectively. Though trees, shrubs and lianas never exceeded 33%, they were represented by 157 pollen taxa (about 62% of the total microflora). They delineated three main pollen zones based on differences in tree pollen percentages. Pollen zone 1 from the base (38,500 cal year B.P.)-917 m (22,000 cal year B.P.) was characterized by influx of minimum values of pollen and spores which they opined resulted from the reduction in the organic material transport from the continent. However, ferns and Typha were dominant with low arboreal pollen except Podocarpus. Again pollen types from Sahara, Sahelian desert and wooded grasslands also occurred in appreciable amounts together with numerous pollen types from the Guineo-Congolian ecosystems. The results indicated the permanency of rainforests and secondary forests in the Niger river catchment possibly as gallery forests along rivers during the last glacial period when enhanced trade wind conditions led to dry conditions. They pointed out that the increase in forest diversity and the expansion of rain and secondary forests in the adjoining continent could have arisen from the post glacial warming coupled with increase in monsoon fluxes over West Africa. Furthermore, they were able to note migration rates of tropical forest populations throughout North West Africa at the beginning of the Holocene through comparison of the KW31 pollen data and continental pollen data from 5°S to 25°N. They further reported the vegetation response to the shift toward aridity recorded widely at the end of the African Humid Period around 4000 cal yr B.P. This wet period had been proposed by DeMenocal et al. (2000) from the study of ODP site 658C off Cape Blanc and was corroborated by the fresh water algal records of Lezine and Cazet (2005).
Hooghiemstra et al. (2006) had studied the distribution patterns of pollen grains in recent marine sediments off NW Africa. They related the distribution patterns to modern pollen source areas (vegetation belts) and the prevailing transport systems (wind belts and ocean currents). Revealing the positions of these vegetation belts in the adjacent continent were the patterns of Quercus, Artemisia, Chenopodiaceae-Amaranthaceae, Ephedra, Gramineae and wet forest trees. The northeast trade winds and the African Easterly Jet (AEJ) were responsible for wind transport of pollen species. They further observed that rivers also transported pollen and fern spores in the rain forest belt. When they compared the recent pollen rain samples from terrestrial and marine sites between 21 and 12°N, clear latitudinal ranges of the vegetation belts were evident in the pollen samples of both environments. A migration of the Southern border of the Sahara was reflected by the changing ratio between Chenopodiaceae-Amaranthaceae pollen from the desert and Gramineae pollen from the savannah belt.
Based on the foregoing, this present study was undertaken to investigate the effects of the Plio-Pleistocene climatic changes/sea level changes as contained in Western Niger Delta palynomorphs.
MATERIALS AND METHODS
Description of study area: The Tertiary Niger Delta (Fig. 1) is a large arcuate delta of the destructive, wave dominated type, which is found in Southern Nigeria. It covers an area of about 7500 km2 and is composed of an overall regressive clastic sequence, which reaches a maximum thickness of 9,000-12,000 m (30,000-40,000 feet). It covers an area located between longitude 4°-9° E and latitude 4°-9° N. Its development has been dependent on the balance between the rate of sedimentation and the rate of subsidence. The stratigraphic distribution of surface outcrops of Tertiary rocks in Southern Nigeria has been previously established by Reyment (1965) and Adegoke (1969). However, the first information on the subsurface distribution of stratigraphic units in the Delta was provided by Short and Stauble (1967) and Frankl and Cordry (1967). According to Short and Stauble (1967), the recent Niger Delta may be divided into 3 main sedimentary environments: the continental, transitional and the marine environments. The continental environment comprises the alluvial environments including the braided stream and meander belt system of the upper deltaic plain. The sediments deposited in this zone are predominantly sandy. Feldspar grains are fairly common and sandy grains commonly are limonite coated. Fine grained sediments (silt and clay) are deposited in the adjacent fresh water back swamps and ox-bows together with large quantities of plant remains. The transitional environment comprises the brackish water lower deltaic plain (mangrove swamp, flood plain, marsh) and the coastal area with its beaches, barrier bars and lagoons.
The sediment in this environment are distinctly finer-grained than in the continental environment. Feldspar is scarce and brackish water faunas may occur. The marine environment includes part of the delta fringe with its fine sand, silt and clay and the associated marine faunas. The environment grades laterally into the holomarine environments, which is not affected by deltaic activity (Short and Stauble, 1967).
Sample collection: Ninty six and eighty nine ditch cutting samples of the wells A and wells B from the offshore Western Niger Delta (Fig. 1), were donated for the investigation by Chevron Nigeria Ltd. in January 2005.
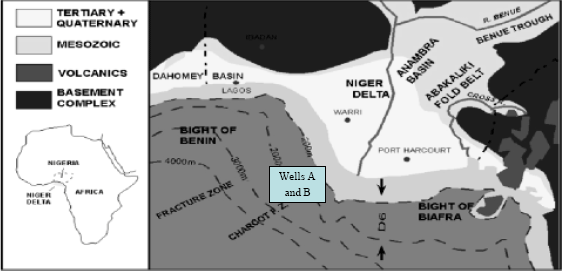 | |
| Fig. 1: | Map of the Niger Delta showing the location of the studied wells after Doust and Omatsola (1990) |
Sample preparation: Thirty grams of each sample was initially treated with hot Hydrochloric acid to remove carbonates, prior to complete digestion in (60%) Hydrofluoric Acid (HF) under a fume cupboard. Agitation of the acid/sample mixture was carried out to aid digestion. The sample was further treated with hot Hydrochloric Acid (HCL) and wet-sieved over a 5 micro mesh polypropylene sieve. The 5-micro sieve was used so that small sporomorphs such as Zonocostites ramonae were not washed off. The Branson Sonifier 250 was further employed during sieving to facilitate complete removal of silt and clay particles. Each residue was oxidized using concentrated nitric acid (HNO3). The residues were prepared for study as strewn mounts using Loctite on 22x32 mm cover slips. The slides were stained with Safranin O to enhance the study of dinoflagellate cysts.
Sample analysis: The prepared samples were analyzed between May to December 2005 in the Paleobotanical Laboratory of the University of Lagos. A Leitz Dialux 20 EB microscope was used for the analysis. All species (pollen, spores, dinoflagellate cysts, fungal remains, algae and foraminiferal linings) present were recorded. This enhanced the palyno-ecological groupings and paleovegetational/paleoenvironmental reconstructions processes.
Coordinates of index markers were aligned with the England Finder coordinates for easier location of the specimens on the slides.
The slides, residues, unprocessed samples, negatives and duplicate prints are in the palynological collections of the Department of Biological Sciences, Redeemer’s University, Mowe, Ogun State, Nigeria.
Species identification: The different palynomorphs (pollen, spores) were identified by comparison with local palynological catalogues of Shell (1980), Legoux (1978), Germeraad et al. (1968), Clarke and Frederiksen (1968), Evamy et al. (1978), Salard-Cheboldaeff (1979, 1990). For dinoflagellates and acritarchs, monographs of Fauconnier et al. (2004), Rochon et al. (1999), Wrenn et al. (1986), Lentin and Williams (1973, 1989), Head and Wrenn (1992), Powell (1992), Fensome et al., (1993, 1996) and Sluijs et al. (2003) were used. Generally, species nomenclature for dinoflagellate cysts followed by Fensome and Williams (2004).
Photomicrography: The specimens were examined and index species and dinoflagellate cysts photographed at X400 and X1000 using the Motic 2.0 camera at the Paleobotanical Laboratory of the University of Lagos.
Ecological grouping: The definition of the ecological groups were based on the study of Sowunmi (1981a, b, 1986), Salard-Cheboldaeff (1981), Rao (2001), Frederiksen (1985), Germeraad et al. (1968) and Rull (2003). Only Zonocostites ramonae and Avicennia sp. are taken as mangrove following (Rao, 2001; Richards, 2001; Germeraad et al., 1968; Frederiksen, 1985), while Monoporites annulatus is listed under savanna according to Germeraad et al. (1968) (Table 1 and 2; Fig. 2-5).
Chronostratigraphy: The chronostratigraphic ages were provided by diagnostic foraminifera using planktonic foraminiferal Zones of Blow (1969, 1979) and Hardenbol et al. (1998). The MFS’s were used as the zonal boundaries for convenience.
| Table 1: | Well A ecological groups abundance/glacial record indicative dinoflagellate cysts data |
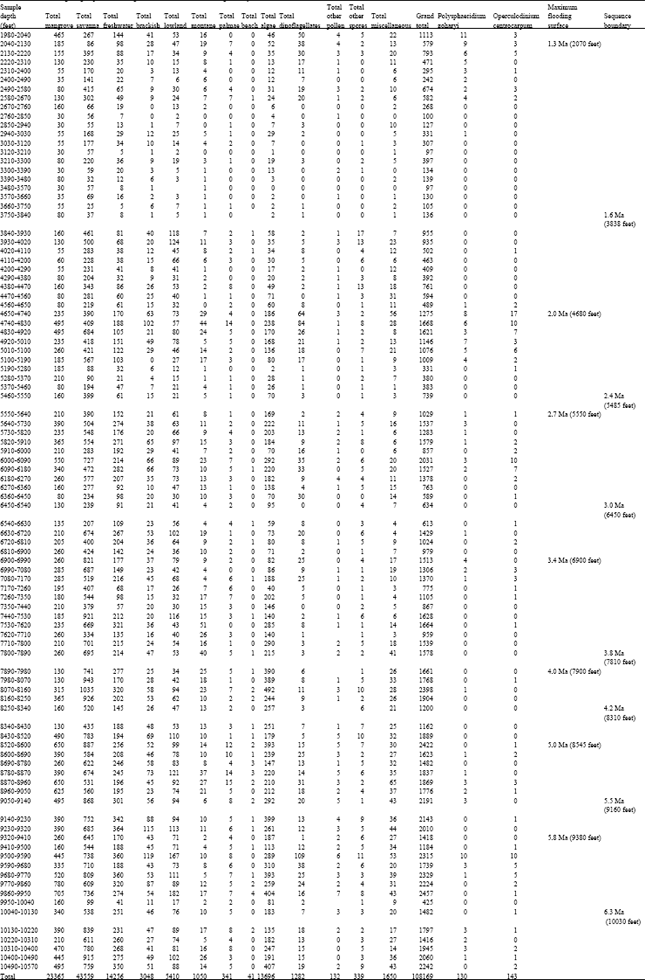 | |
| Table 2: | Well B Ecological groups abundance/glacial record indicative dinoflagellate cysts data |
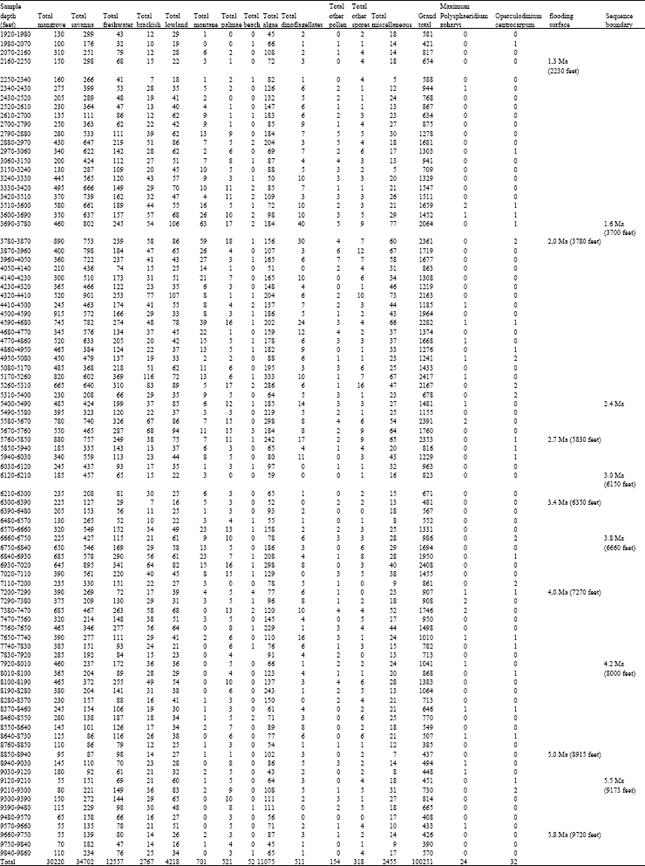 | |
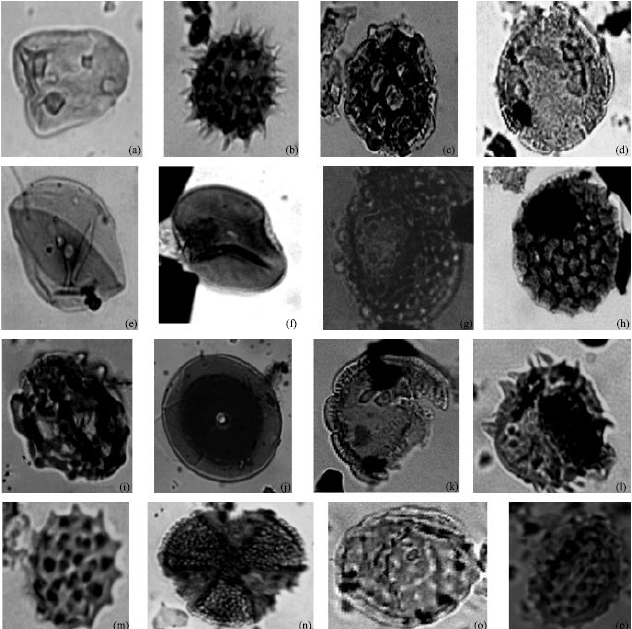 | |
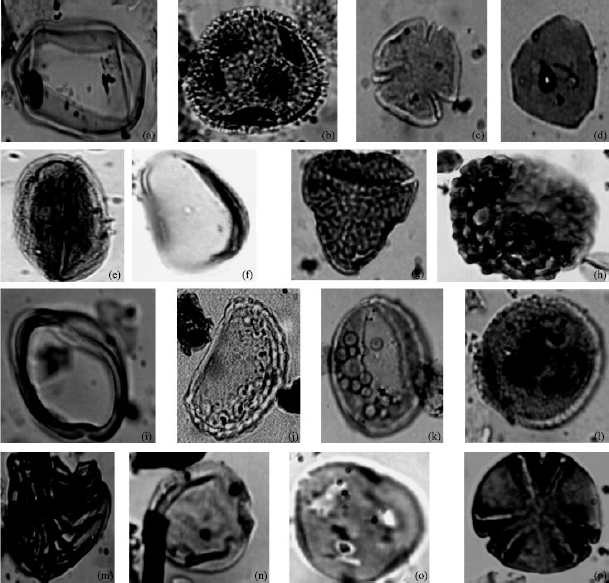
| Fig. 2: | Photomicrographs of some recovered Savanna pollen species from well A and B with corresponding well, sample and England finder location. (a) Cyperaceaepollis sp. Well B (8640-8730) G33/2, (b) Echitricolporites spinosus well A (7350-7440) G30/0, (c) Fenestrites spinosus well B (4590 - 4680) V39/ 1, (d) Retistephanocolpites gracilis well B (7740-7830) T19/2, (e) Monoporites annulatus well A (1980-2040) M24/3, (f) Multiareolites formosus well A (5010-5100) G24/1, (g) Gardenia imperialis well A (5010-5100) P35/2, (h) Polygonum sp. Well A (6720- 6810) N42/2, (i) Peregrinipollis nigericus well B (3690-3780) T52, (j) Numulipollis neogenicus well B (5170-5260) O34/1, (k) Retistephanocolpites gracilis well B (5170-5260) K24/3, (l) Echistephanoporites echinatus well B (2790-2880) T33/4, (m) Tubifloridites antipodica Cookson, 1947; well A (7530 -7620) M41 (n) Margocolporites vanwijhei well B (7650-7740) S39/1, (o) Chenopodipollis sp. (Chenopodiaceae) well B (7740-7830) T18/1 and (p) Tubifloridites antipodica well B (5400-5490) K24/3 |
Presentation of results: The analysis sheets (data sheets) were inputed on STARTABUGS, from which distribution charts showing the palynomorph species arranged by stratigraphic appearances and disappearances against increasing depth in the wells were generated, with the log and systems tracts by the side. The marker species were plotted separate from other species. This facilitated the picking of the palynological zonal boundaries easily.
Statistical analysis: Mean, standard deviation, student T-test, one way Analysis of Variance (ANOVA) and correlation analysis were performed on the different ecological groups using the Prism version 5.00 computer software programme (GraphPad Software, San Diego, CA).
 | |
| Fig. 3: | Photomicrographs of some Freshwater swamp species recovered from wells A and B with corresponding well, sample and England Finder location. (a) Nympheaepollis clarus well B (2790 -2880) M22/2, (b) Retitricolporites irregularis well A (2130 -2220) Q48/4, (c) Psilatricolporites operculatus well A(2310 -2400) V36/3, (d) Momipites sp. Well A (7260-7350) G35/1, (e) Striatricolpites catatumbus well B(5400-5490)W26/1, (f) Laevigatosporites sp. Well A (1980-2040) L28/2, (g) Lycopodium sp. Well A (1980-2040) K42/4, (h) Crassoretitriletes vanraadshooveni well B (3330-3420) D27/2, (i) Nympheaepollis clarus well B (4590-4680) R38/2, (j) Verrucatosporites sp. Well A (4590-4680) R38/2, (k) Gemmamonoporites sp. well B (8370-8460) R22/2, (l) Retibrevitricolporites obodoensis well B (5400-5490) U19/3, (m) Magnastriatites howardi well A (2490-2580) S42/1, (n) Anthocleista vogelii well B (3780-3870) S38/4, (o) Anacolosidites cf. luteoides well A (5400-5490)P28/2 and (p) Rauwolfia vomitora well A (5400-5490) S35/3 |
Significant limits were set at the 95% confidence interval.
RESULTS
Palynostratigraphy
Pollen, spores and algae: Ninety six and eighty nine samples of wells A and well B between (1980-10570 feet) and (1920-9860 feet) were analyzed. The palynomorphs recovered were abundant and diverse in most of the horizons studied. The assemblages were dominated by land derived sporomorphs particularly Monoporites annulatus, Zonocostites ramonae, Podocarpus milanjianus, Stereisporites sp., Cyperaceaepollis sp., Sapotaceoidaepollenites sp., Retistephanocolpites gracilis (aff. Borreria verticillata), Podocarpidites sp., Verrucatosporites sp., Laevigatosporites sp., Polypodiaceoisporites sp. and Acrostichum aureum.
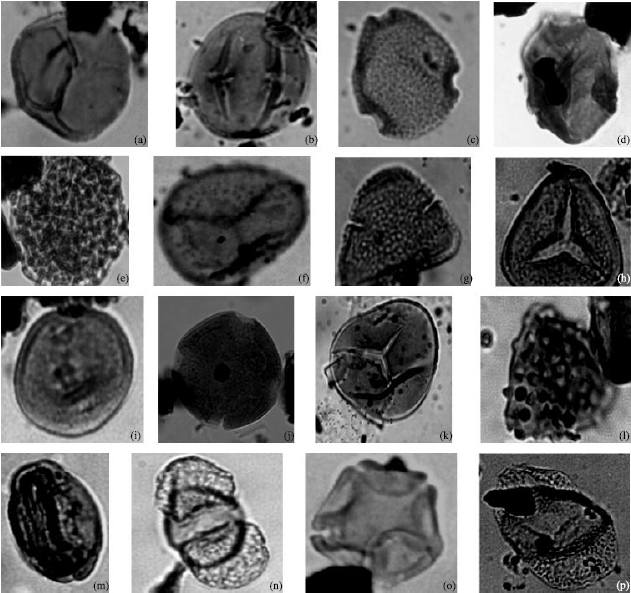 | |
| Fig. 4: | Photomicrographs of some Lowland Rainforest species recovered in well A and B, with the corresponding well, sample and England Finder liocation. (a) Stereisporites sp. (Smooth) well B (2880-2970)H33/4, (b) Sapotaceoidaepollenites sp. Well B (5400-5490) S32/1, (c) Canthium sp. well B (5760-5850) X17/3, (d) Corsinipollenites jussiaensis well A (10400-10490) J41/1, (e) Spirosyncolpites bruni well B (6750-6840) S45/2, (f) Sterisporites sp. (Rough) well B (2700 -2790) X48/2, (g) Bombacidites bellus well A (7800 -7890) N38/4, (h) Polypodiaceoisporites sp. well B (2700 -2790) M43/3, (i) Psilatricolporites crassus well A (5640 -5730) T34/3, (j) Pachydermites diederixi well A(4470 -4560) N36/3, (k) Acrostichum aureum well A (9590 -9680) N36/3, (l) Echitriporites trianguliformis well B (3420 -3510) H27/3, (m) Marginipollis concinnus well B (2700 -2790) X48/2, (n) Podocarpus milanjianus well A (6000-6090)T35/3, (o) Alnipollenites verus well A (6000-6090)T35/3 and (p) Podocarpidites sp. well A (2220-2310) J30/1 |
Two hundred and thirty palynomorph species were recorded in well A, while well B yielded one hundred and seventy-three species.
Dinoflagellates/marine elements: Abundant records of dinoflagellate cysts which peaked at some horizons characterized the well A. The dominant species were Polysphaeridium zoharyi, Operculodinium centrocarpum, Spiniferites ramosus, Achomosphaera ramulifera, Tuberculodinium vancampoae, Hystrichokolpoma rigaudiae, Spiniferites sp. (S. membranaceus, S. mirabilis, S. delicatus, S. bulloideus, S. elongatus, S. bentorii and S. hyperacanthus), Selenopemphix nephroides, Achomosphaera andalousiensis, Achomosphaera sp., Selenopemphix sp. and the acritarch Leiosphaeridia sp.
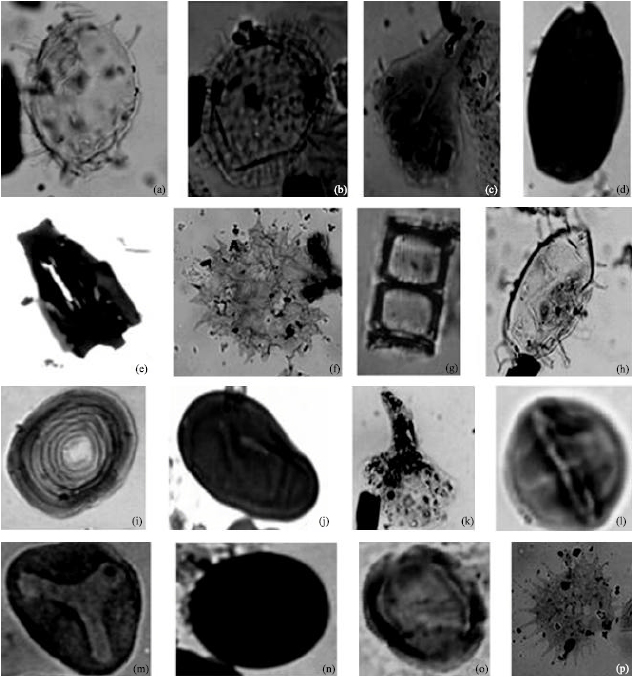 | |
| Fig. 5: | Photomicrographs of some dinoflagellate cycts and accessories recovered from wells A and B, with the corresponding well, sample and England Finder location. (a) Polysphaeridium zoharyi well A (2880-2970) H33/4, (b) Operculodinium centrocarpum well B (3690- 3780) O35/1, (c) Botryococcus braunii well B (4860-4950) K38/4, (d) Fungal spore. well B (5760-5850) O38/2, (e) Charred Graminae Cuticle well A (2130-2220) X36/1, (f) Pediastrum sp. Well B (8370-8460) W38/1, (g) Diatom Frustule well B (4680-4770) K20/2, (h) Polysphaeridium zoharyi well A (2880-2970) H33/4, (i) Concentricytes circulus well B (4680-4770) K20/2, ( j) Fungal spore well A (10400-10490) V18/2, (k) Botryococcus braunii well B (3690-3780)K24/2, (l) Zonocostites ramonae well B (3690-3780) L24/3, (m) Elaeis guineensis well A (7260- 7350) T12/1, (n) Fungal spore well A (3510-3600) M37/4, (o) Zonocostites ramonae well B (3780-3870) K24/1 and (p) Pediastrum sp. well B (5760-5850) K38/4 |
The recoveries were moderate in well B, with Polysphaeridium zoharyi, Spiniferites ramosus, Operculodinium centrocarpum, Protoperidinium sp. (Pteridinium sp. of Wall et al., 1977), Selenopemphix nephroides, Hystrichokolpoma rigaudiae, Spiniferites sp., Achomosphaera sp., Selenopemphix sp., Brigantedinium sp., Tectatodinium sp. and the acritarch Leiosphaeridia sp. as the commonest species. Deepening at some intervals was indicated by the records of Impagidinium sp., Nematosphaeropsis labyrinthus and N. lemniscata. The spot records of Dinogymnium euclaense, Muderongia sp. and Odontochitina sp., which are Maastrichtian and older dinocysts at 6000 and 8160 feet, in the well A, together with such older palynomorphs as Monocolpopollenites sphaeroidites, Ephedripites sp., Gnetaceaepollenites sp., Foveotriletes margaritae, Proteacidites longipinosus, Psilamonocolpites marginatus, Cinctiperiporites mulleri, Corsinipollenites jussiaeensis, Retimonocolpites obaensis all suggest the reworking of older sediments onto younger horizons.
The preponderance of the paleoenvironmental indicators dinoflagellate cysts, microforaminiferal wall linings and freshwater algae Botryococcus braunii and Pediastrum sp., in the studied sections of the wells suggests sediment deposition in a shallow marine environment with frequent freshwater incursions.
Age: The wells A and B sequences fell within the broad pan-tropical Echitricolporites spinosus zone of Germeraad et al. (1968) and the P800-P900 zones of Evamy et al. (1978). The sequence also correlated with the J2-J3 Zones of Legoux (1978). The ages of the studied sections of the wells ranged from Late Miocene (P860) at the base to Early Pleistocene (P900) subzone at the top. This is attributed to the Quantitative Base and Base Occurrences of the Niger Delta palynological Miocene/Pliocene boundary marker species Retistephanocolpites gracilis (aff. Borreria verticillata) at 8520 and 9030 feet respectively in wells A and B (Evamy et al., 1978). Again the 5.0 Ma MFS’s were delineated at 8545 feet and 8915 feet in wells A and B, defined by the FDO’s of Cyclammina minima, Eggerella scabra and LDO of Globorotalia tumida as well as the marked influx of Arenaceous foraminifera. These events further indicated the penetration of Late Miocene below these depths. The uppermost part of the wells are dated Lower Pleistocene based on the first peak abundance of Podocarpus milanjianus at 2040 feet and the presence of Spiniferites elongatus, Spiniferites bulloideus and Spiniferites hyperacanthus (Powell, 1992; Rochon et al., 1999), coupled with the FDO: Globorotalia miocenica dated 2.0 Ma at 4680 and 4650 feet, respectively for wells A and B.
Ecological group assessment: The ecological group trends in the two wells appeared similar with the savanna dominating over mangrove and the other ecological groups especially in well A (Fig. 6 and 7; Table 1 and 2). The least were beach, other pollen, other spores and Palmae. The abundance of freshwater algae especially Botryococcus braunii is in agreement with Morley (1995), who recognized its abundance in the Pliocene-Pleistocene of the Niger Delta. There were clear crests and troughs in the microfloral abundance, suggesting alternations of dry and wet climates.
The statistical tests revealed significant relationships between all the ecological groups (at 95%) confidence interval except the marine elements in the well A. A comparison of the wells A and B showed that the means of the ecological groups were significantly different and the Bartlett's test for equal variances depicted significant differences. An entirely different scenario was observed in the well B, there were significant relationships between the mangrove, savanna, montane and marine elements while none existed for the freshwater, brackishwater, lowland rainforest, palmae, beach, algae and other spores.
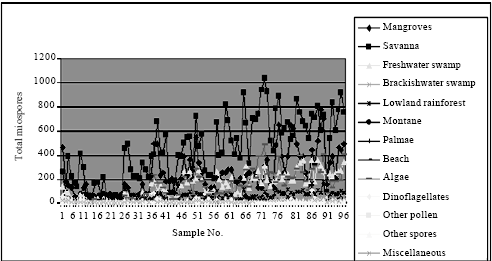 | |
| Fig. 6: | Well A ecological groups abundance plot |
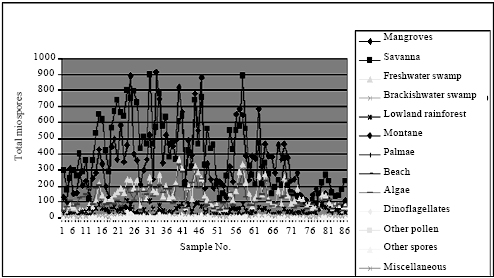 | |
| Fig. 7: | Well B ecological groups abundance plot |
This shows that though the two wells were from the same field, their miospore associations are slightly different.
Dinoflagellate stratigraphy: The dominant dinoflagellate cysts in well A were Operculodinium centrocarpum and Polysphaeridium zoharyi accounting for 11.15 and 10.14% respectively (Fig. 5). The dominance of these two species which were concentrated at some points could have resulted from lowered sea levels following glacial maxima (Mozardec-Kerfourn, 1988, 1992a, b). A critical evaluation of well B, revealed the dominant dinoflagellate cysts to be Operculodinium centrocarpum, Polysphaeridium zoharyi, Protoperidinium sp. (Peridinium sp. of Wall et al., 1977), Spiniferites ramosus, Invertocysta sp. and the acritarch species Leiosphaeridia sp.
Evidence of climatic variations, sea level changes and contributions to the global glacial history: As highlighted earlier the chronostratigraphic ages for the sediments were provided by diagnostic foraminifera which enabled correlation to worldwide chronostratigraphy. The MFS’s were used as the zonal boundaries for convenience. The recognized SB’s were dated by comparison with the Global Sequence Cycle Chart of Haq et al. (1988) as revised by Berggren et al. (1995) and updated by Grandstein et al. (2004).
Early pleistocene (c.a 1.3-2.0 Ma): Top defined by the FDO of Globorotalia truncatulinoides and Hyalinea balthica. This horizon corresponds to the 1.3 Ma MFS. The zonal base is defined by the 2.0 Ma MFS marked by the FDO of Globorotalia miocenica and the LDO’s of Pulleniatina obliquiloculata and Pulleniatina finalis. Well A: (2070-4680 feet) and Well B: (2040-3750 feet) Early Pleistocene.
In the well A, except at few points where the counts of Zonocostites ramonae exceeded Monoporites annulatus, (Table 1) the microfloral assemblage is dominated by Monoporites annulatus and other savanna species. This trend occurred with common records of the dinocysts Polysphaeridium zoharyi and Operculodinium centrocarpum between 1980-2670 feet suggesting a marked dry climate possibly associated with lowered sea level (glacial maximum) around this period in the Niger Delta. This event is demonstrated again around the 2.0Ma MFS at 4680, where Polysphaeridium zoharyi, occurred regularly from 4560-5190 feet in association with abundant savanna species. In the well B (Table 2), around the 2.0Ma MFS at 3750 feet are clusters of the dinocysts Polysphaeridium zoharyi and Operculodinium centrocarpum between 3510-3870 feet. The few mangrove peaks at (2070 and 3780 feet) show possible wet periods sandwiched in-between an extremely dry climate.
Early-late pliocene (c.a 5.0-2.0 Ma): The top of this Zone is defined by the FDO of Globorotalia miocenica and the LDO’s of Pulleniatina obliquiloculata and Pulleniatina finalis which coincided with the 2.0 Ma MFS. The base is defined by the FDO’s of Cyclammina minima, Eggerella scabra and LDO of Globorotalia tumida. The horizon coincides with the 5.0 Ma MFS. It also lies close to the Base Occurrence/Quantitative Occurrence of the savanna pollen specie Retistephanocolpites gracilis (aff. Borreria sp.), which is the index Pliocene /Miocene boundary marker in the Niger Delta (Evamy et al., 1978).
0-2.7 Ma: The top of this Zone is defined by the FDO of Globorotalia miocenica and the LDO’s of Pulleniatina obliquiloculata and Pulleniatina finalis which coincide with the 2.0 Ma MFS. The base is defined by the FDO’s of Bulimina marginata and Cyclammina cancellata as well as the LDO of Sphaeroidenella dehiscens, which is dated 2.7 Ma.
2.0-2.7 Ma Well A: (4680-5580 feet); Well B: (3750-5490 feet): Savanna species dominated the well A microfloral assemblage within this time (2.0-2.7 Ma) except at 5190-5370 feet where mangrove peaked, showing a possible wet period in-between an extremely dry climate. The wet period is supported by common freshwater swamp species. The common records of Polysphaeridium zoharyi from 5550-6180 feet, revealed a well defined dry climate around the 2.7 Ma.
Within this time lapse in well B, there were peaks of mangrove at (4500 and 4860 feet) and a longer wet period between 5080-5850 feet. A marked drier period supported by common records of Polysphaeridium zoharyi occurred between 4860-5080 feet.
2.7-3.4 Ma: The top is defined by the FDO’s of Bulimina marginata and Cyclammina cancellata as well as the LDO of Sphaeroidenella dehiscens, which are dated 2.7 Ma. The base of the Zone is defined by the FDO’s of Rectuvigerina multicostata, Amphistegina lessonii, Globigerinoides bollii, Globorotalia margaritae and Globorotalia pseudomiocenica, which is correlated with the 3.4 Ma MFS.
Well A: (5580-6900 feet) and well B: (5490-6390 feet): In well A, savanna species dominated the microfloral assemblages, the common freshwater swamp species suggests possible climatic fluctuations. Polysphaeridium zoharyi became common from 6900-7170 feet, Operculodinium centrocarpum was common between 5820-6180 and 6990-7260 feet. This indicates a possible glacial maxima and lowered sea level around 2.7 Ma. On the other hand in well B, there were peaks of mangrove between 5490-5850 feet and 6210-6390), in-between savanna dominated microflora. The dinocysts Polysphaeridium zoharyi and Operculodinium centrocarpum occurred in spots around 5310-6030 feet.
3.4-4.0 Ma: The top of this Zone is defined by the FDO’s of Rectuvigerina multicostata, Amphistegina lessonii, Globigerinoides bollii, Globorotalia margaritae and Globorotalia pseudomiocenica, which is correlated with the 3.4 Ma MFS. Its base coinciding with the 4.0 Ma MFS is defined by the FDO’s of Marginulina costata, Haplophragmoides compressa and Globigerina nepenthes.
3.4-4.0 Ma-well A: (6900-7980 feet) and well B: (6390-7170 feet): Abundant savanna species dominated the microfloral assemblage. Polysphaeridium zoharyi and Operculodinium centrocarpum were common between 6900-7170 and 6990-7350 feet, respectively. The assemblagein well A indicates an extremely dry climate. While in well B, the microfloral assemblage was dominated by savanna species. The dinocyst Polysphaeridium zoharyi was absent, while Operculodinium centrocarpum occurred in spots around 6660-6930 feet. This suggests an extremely dry climate and agrees with the results from well A.
4.0-5.0 Ma: The top of this Zone is defined by the FDO’s of Marginulina costata and Globigerina nepenthes. It coincides with the 4.0 Ma MFS. The base is defined by the FDO’s of Cyclammina minima, Eggerella scabra and LDO of Globorotalia tumida.
Well A: (7980-8570 feet) and well B: (7170-8880 feet): This interval is characterized by extremely high records of savanna pollen and common Retistephanocolpites gracilis (aff. Borreria sp.). The records of the dinocysts Polysphaeridium zoharyi and Operculodinium centrocarpum were sparse in well A, while mangrove pollen, in association with common freshwater swamp, brackish water swamp species dominated the microfloral assemblage in well B. The dinocyst Polysphaeridium zoharyi was common between 7200-7470 feet and later occurred in spots within this section. Operculodinium centrocarpum occurred in spots all through. This suggests a period of climatic fluctuations with brief dry periods and fllowered sea levels.
Late miocene: 5.0-5.8 Ma: The top is defined by the FDO’s of Cyclammina minima, Eggerella scabra and LDO of Globorotalia tumida. The base of this Zone dated 5.8 Ma MFS is defined by the FDO’s of Haplophragmoides narivaensis and Globoquadrina dehiscens.
Well A: 5.0-5.8 Ma(8570-9380 feet) and well B: (8880-9720 feet): This section is of Late Miocene age, savanna species still dominated the assemblage in well A. This occurred with common freshwater swamp species, mangrove peaked between 9770-10040 feet, indicating a possible wet inter-phase between a dominantly dry climate. This section is characterized by abundant records of savanna species, mangrove just peaked between 8940-9120 feet in well B. The dinocysts Polysphaeridium zoharyi and Operculodinium centrocarpum were sparse in occurrence.
>5.8 Ma well A: (9380-10570 feet) and well B: (9720-9860 feet): Savanna species still dominated the microfloral assemblage within this thin section older than 5.8 Ma. The dinocysts Polysphaeridium zoharyi and Operculodinium centrocarpum were sparse to absent.
Paleoclimatic deductions: The dominance of Polysphaeridium zoharyi and Operculodinium centrocarpum which were concentrated at some points in the two wells could have resulted from lowered sea levels following glacial maxima (Mozardec-Kerfourn, 1988, 1992a, b). The 1.3, 2.0, 2.7, 3.4, 4.0, 5.0 and 5.8 Ma Maximum Flooding Surfaces were respectively dated using diagnostic foraminifera. Close to these MFS’s were clusters of Polysphaeridium zoharyi and Operculodinium centrocarpum, especially around the 2.0-2.7 Ma MFS’s in well A (4650-5640 feet) and (3780-5850 feet) in well B (Table 1 and 2). The signature being more pronounced in well A. Furthermore, this trend was slightly repeated between the 2.7-3.4 Ma MFS’s (5640-6900 feet) and (5850-6390 feet) in wells A and B, respectively. Further downhole, between the 3.4-4.0 Ma MFS’s well A (6900-7900 feet) and well B (6350-7270 feet), were spot records of the two species. The recoveries between the 4.0-5.0 Ma MFS’s were similar to the former, while below the 5.0 Ma MFS (Late Miocene) in well B were spot records of these species contrary to well A, in which a remarkable increase in the abundance of the two species especially below the 5.8 Ma MFS down to the last sample analyzed 10570 feet were observed.
DISCUSSION
A predominantly dry climate is inferred for these sediments due to the abundance of Monoporites annulatus (Graminae), Cyperaceaepollis sp., Chenopodipollis sp. and other savanna species. This agrees with the reports of Morley (1995) who inferred from late Miocene data from the Niger Delta which spanned a period of about 1.7 Ma, that the abundance of Gramineae pollen and common mangrove pollen within lowstand fan deposits was consistent with a dry climate and widespread representation of mangroves on the lower delta plain. Possibly this trend continued into the Pliocene and younger ages. Again, Hooghiemstra et al. (1986) had also associated the abundance of Gramineae pollen and Chenopodiaceae-Amaranthaceae with the dry conditions of desert and savanna. Moreover, Pocknall et al. (2001) in carrying out an integrated paleontological study of Pliocene to Pleistocene sediments of the Orinoco Delta in Eastern Venezuela and Trinidad, had attributed the increase in the abundance of Cyperaceae in the Pleistocene to the same climatic (cooling) event that led to the introduction of Alnus into the Andes which they observed coincided with the appearance of Avicennia sp. In these studied Niger Delta wells apart from the abundant records of Cyperaceae pollen, the moderate records of Podocarpus milanjianus could have arisen from the same cooling event reported by Pocknall and co-workers.
An assessment of the dinoflagellate cysts records further gives credence to a dominantly dry climate coupled with warm temperatures in the Pliocene-Pleistocene of the Niger Delta. Mozardec-Kerfourn (1988, 1992a, b) had associated Polysphaeridium zoharyi with dry climate and lowered sea levels, while Udeze and Oboh-Ikuenobe (2005) linked Operculodinium centrocarpum with warm climate. These trends were clearly revealed in the well A, around the 2.0 Ma MFS at 4680 feet, where Polysphaeridium zoharyi, occurred regularly from 4560-5190 feet in association with abundant savanna species. In the well B, around the 2.0 Ma MFS at 3750 feet were clusters of the dinoflagellate cysts Polysphaeridium zoharyi and Operculodinium centrocarpum between 3510-3870 feet. The most pronounced period of glacial maxima and lowered sea level as revealed by the results from the wells A and B are the periods close to and around the 2.0-2.7 Ma MFS. This particular event has been reported by Molnar and Cane (2004) who opined that early to Middle Pliocene global climate was clearly different from those since 2.7 Ma. They further reported that, around 2.7 Ma, the warm equable climates of early and middle Pliocene time (5-2.7 Ma) were replaced by recurring ice ages.
The inferred warm climate and lowered sea levels around the 3.4-2.7 Ma from the studied samples concurs with the earlier reports from Colombia by Hooghiemstra (1993) who suggested warm climatic conditions around the 3.2-2.7 Ma.
Leroy and Dupont (1994) had recognized this event from their study of a 200 m marine pollen record from northwestern Africa. Their study revealed cyclic vegetational and continental climatic fluctuations in this area from 3.7-1.7 Ma. The cycles paralleled oxygen isotope stages. Tropical forests and mangrove swamps got to Cape Blanc about 5°N of the present distribution before 3.5 Ma. Between 3.5 and 2.6 Ma, forests occurred and later disappeared after which a Saharan paleo-river flowed until isotope stage 134 (3.35 Ma). After this period wind transport of pollen grains prevailed over fluvial, as pollen indicators of trade winds increased between 3.3 and 2.5 Ma. They suggested a strong aridification of the Northwestern African climate during isotope stage 130 (3.26 Ma) after which humid conditions got re-established followed by another aridification around 2.7 Ma.
Pocknall et al. (2001) had related the dominance and subsequent decline of Grimsdalea magnaclavata to a possible climatic warming and cooling which coincided with the major regional lowstand displayed on Haq et al. (1987) Global sea level curve at around 2.1 to 2.0 Ma. This same event is clearly revealed in the well A, around the 2.0 Ma MFS at 4680 feet, where Polysphaeridium zoharyi, occurred regularly from 4560-5190 feet in association with abundant savanna species. In the well B, around the 2.0 Ma MFS at 3750 feet were clusters of the dinoflagellate cysts Polysphaeridium zoharyi and Operculodinium centrocarpum between 3510-3870 feet. Furthermore, the cooling events must have encouraged the preponderance of Podocarpus milanjianus between 4650-4920 feet in well A and 3510-4230 feet in well B respectively. The few mangrove peaks at (2070 and 3780 feet) in well B indicate possible wet period in-between an extremely dry climate.
CONCLUSION
The 2.0 and 2.7Ma glacial maxima events which had been reported in different parts of the world, has been indicated in Western Niger Delta sediments. The evidence is clearly demonstrated by both marine and terrestrial palynomorphs.
ACKNOWLEDGMENTS
The author is grateful to the Management of Chevron Nigeria limited for donating the ditch cutting samples and providing other data for this research work. I also appreciate Prof. O.K Ulu of the Earth Sciences and Reservoir Services Division for his guidance and assessment of the manuscript.
To Prof. Niels Poulson of the Geological Survey of Denmark and Greenland (GEUS) for the identification and confirmation of the dinoflagellate cysts.
To Prof. O. S. Adegoke and the Management of Mosunmolu limited for allowing me use some of their facilities.
To Dr. Francisca Oboh-Ikuenobe of the Geology Dept. University of Missouri, Rolla for her wise counsel and for providing relevant journals.
REFERENCES
- Adam, D.P., A.M. Sarna-Wojcicki, H.J. Rieck, J.P. Bradbury, W.E. Dean and R.M. Forester, 1989. Tulelake, California: The last 3 million years. Palaeogeogr. Palaeoclimatol. Palaeoecol., 72: 89-103.
Direct Link - Allen, J.R., 1964. The nigerian continental margin: Bottom sediments, submarine morphology and geological evolution. Mar. Geol., 1: 289-332.
CrossRef - Allen, J.R. and J.W. Wells, 1962. Holocene coral banks and subsidence in the Niger Delta. J. Geol., 70: 381-397.
Direct Link - Adekanmbi, O.H. and O.T. Ogundipe, 2009. Pollen grains of lagos lagoon swamp and hinterland vegetation-I. Int. J. Bot., 5: 270-278.
CrossRefDirect Link - Biffi, U. and D. Grignani, 1983. Peridinioid dinoflagellate cysts from the Oligocene of the Niger Delta, Nigeria. Microplaeontology, 29: 126-145.
Direct Link - Clarke, R.T. and N.O. Frederiksen, 1968. Some new sporomorphs from the Upper Tertiary of Nigeria. Grana Palynol., 8: 210-224.
CrossRefDirect Link - DeMenocal, P., J.T. Oritz, J. Guilderson, M. Adkins and M. Sarnthein et al., 2000. Abrupt onset and termination of the African humid period: Rapid climate responses to gradual insulation forcing. Quaternary Sci. Rev., 19: 347-361.
CrossRefDirect Link - Dupont, L.M. and M. Welnelt, 1996. Vegetation history of the Savanna corridor between the guinean and the congolian rain forest during the last 150,000 years. Veg. Hist. Archeobot., 5: 273-292.
CrossRefDirect Link - Dupont, L.M., F. Marret and K. Winn, 1998. Land sea correlations by means of terrestrial and marine palynomorphs from the equatorial East Atlantic: Phasing SE trade winds and the ocean productivity. Palaeogeogr. Palaeoclimatol. Palaeoecol., 142: 51-84.
CrossRef - Dupont, L.M., S. Jahns., F. Marret and N. Shi, 2000. Vegetation change in equatorial West Africa: Time-slices for the last 150 ka. Palaeogeography Palaeoclimatol. Palaeoecol., 155: 95-122.
CrossRef - Evamy, B.P., J. Haremboure, P. Kamerling, W.A. Knaap, F.A. Molly and P.H. Rowlands, 1978. Hydrocarbon habitat of tertiary Niger Delta. Am. Assoc. Pet. Geol. Bull., 62: 1-39.
Direct Link - Finch, J.M. and T.R. Hill, 2008. A late quaternary pollen sequence from Mfabeni Peatland South Africa: Reconstructing forest history in maputaland. Quaternary Res., 70: 442-450.
CrossRef - Germeraad, J.H., C.A. Hopping and J. Muller, 1968. Palynology of tertiary sediments from tropical areas. Rev. Paleobot. Palynol., 6: 189-348.
CrossRef - Haack, R.C., P. Sundaraman, J.O. Diedjomahor, H. Xiao, N.J. Gant, E.D. May and K. Kelsch, 2000. The Niger Delta Petroleum Systems, Nigeria. In: Petroleum Systems of South Atlantic Margins, Mello, M.R. and Katz, B.J. (Eds.). Vol. 73. American Association of Petroleum Geologists, Tulsa, OK, USA., pp: 213-231.
- Haq, B.U., J. Hardenbol and P.R. Vail, 1987. Chronology of fluctuating sea levels since the triassic. Science, 235: 1156-1167.
CrossRefDirect Link - Haq, B.U., J. Hardenbol and P.R. Vail, 1988. Mesozoic and Cenozoic Chronostratigraphy and Cycles of Sea- Level Change. In: Sea-Level Changes: An Integrated Approach, Wilgus, C.K., B.S. Hastings, C.G. St. C. Kendall, H.W. Posamentier, C.A. Ross and J.C. Van Wagoner (Eds.). Vol. 42. SEPM Publication, USA., pp: 72-108.
- Hardenbol, J., J. Thierry, M.B. Farley, T. Jacquin, P.C. De Graciansky and P.R. Vail, 1998. Meozoic-Cenozoic Sequence Chronostratigraphic Framework of European Basins. In: Mesozoic and Cenozoic Sequence Stratigraphy of European Basins, De Graciansky, P.C., J. Hardenbol, T. Jacquin and P.R.J. Vail (Eds.). Vol. 60. SEPM, Tulsa, Okla, pp: 3-13.
- Hooghiemstra, H., A.M. Lezine, S.A.G. Leroy, L. Dupont and F. Marret, 2006. Late quaternary palynology in marine sediments: A synthesis of the understanding of pollen distribution patterns in the NW African setting. Quaternary Int., 148: 29-44.
CrossRef - Ivanov, D.A., A.R. Ashraf and V. Mosbrugger, 2007. Late Oligocene and miocene climate and vegetation in the eastern paratethys area (Northeast Bulgaria), based on pollen data. Palaeogeog. Palaeoclimatol. Palaeoecol., 255: 342-360.
CrossRefDirect Link - Jahns, S., M. Huls and M. Sarnthein, 1998. Vegetation and climate history of West Equatorial Africa based on a marine pollen record off liberia (site Gik 16776) covering the last 400,000 years. Rev. Palaeobot. Palynol., 102: 277-288.
CrossRefDirect Link - Leroy, S.A.G. and L.M. Dupont, 1997. Marine palynology of the ODP site 658 (N-W Africa) and its contribution to the stratigraphy of late pliocene. Geobios, 30: 351-359.
CrossRef - Leroy, S.A.G. and L. Dupont, 1994. Development of vegetation and continental aridity in northwestern Africa during the Late Pliocene: The pollen record of ODP site 658. Paleogeography Paleoclimatol. Paleoecol., 109: 295-316.
CrossRef - Lezine, A. and J. Cazet, 2005. High-resolution pollen record from core KW31, Gulf of Guinea, documents the history of the lowland forests of West Equatorial Africa since 40,000 yr ago. Quaternary Res., 64: 432-443.
CrossRef - Mozardec-Kerfourn, M.T., 1992. Estuarine Dinoflagellate Cysts Among Oceanic Assemblages of Pleistocene Deep-Sea Sediments from the West African Margin and their Paleonenivronental Significance. In: Neogene and Quartenary Dinoflagellate Cysts and Acritarchs, Head, M.J. and J.H. Wrenn (Eds.). American Association of Stratigraphic Palynologists Foundation, Dallas, pp: 133-146.
- Mozardec-Kerfourn, M.T., 1992b. Upper Pleistocene and Holocene Dinoflagellate Cyst Assemblages In Marine Environments of the Mediterranean Sea and the Northwest Atlantic Coast of France. In: Neogene and Quartenary Dinoflagellate Cysts and Acritarchs. Head, M.J. and J.H. Wrenn (Eds.). American Association of Stratigraphic Palynologists Foundation, Dallas, pp: 121-132.
- Oboh, F.E., M.B. Salami and J.L. Chapman, 1992. Palynological interpretation of the palaeoenvironments of Miocene strata of the well Igbomotoru-1, Niger delta. J. Micropalaeontol., 11: 1-6.
CrossRefDirect Link - Pocknall, D.T., L.J. Wood, A.F. Geen, B.E. Harry and R.W. Hedlund, 2001. Integrated Paleontological studies of Pliocene to Pleistocene deposits of the Orinoco delta, Eastern Venezuela and Trinidad. Proceedings of the 1X International Palynological Congress, (IPC'01), American Association of Stratigraphic Palynologist Foundation, Houston, Texas, USA., pp: 319-326.
- Richards, K., 2001. Palynology and sequence stratigraphy of early Miocene-late Oligocene deltaics in the Sihapas group, central Sumatra, Indonesia. Proceedings of the 11th International Palynological Congress, (IPC'01), American Association of Stratigraphic Palynologist Foundation, Houston, Texas, USA., pp: 253-262.
- Rochon, A., A. De Vernal, J. Turon, J. Matthiessen and M.J. Head, 1999. Distribution of Recent Dinoflagellates Cysts in Surface Sediments from the North Atlantic Ocean and Adjacent Seas in Relation to Sea-Surface Parameters. AASP Contribution Series, 35. American Association of Stratigraphic Palynologists Foundation, North Atlantic, pp: 152.
CrossRefDirect Link - Salard-Cheboldaeff, M., 1979. Palynologie maestrichtienne et tertiaire du cameroun. Etude qualitative et repartition verticals des principales especes. Rev. Paleobotany Palynol., 28: 365-372.
CrossRef - Salard-Cheboldaeff, M., 1981. Palynologie maestrichtienne et tertiaire du cameroun. Resultats Botaniques Rev. Paleobotany Palynol., 32: 401-439.
CrossRef - Salard-Cheboldaeff, M., 1990. Intertropical african palynostratigraphy from cretaceous to late quaternary times. J. Afr. Earth Sci. (Middle East), 11: 1-24.
CrossRefDirect Link - Short, K.C. and J.A. Staeuble, 1967. Outline of Geology of Niger Delta. AAPG Bull., 6: 761-799.
Direct Link - Sluijs, A., H. Brinkhuis, C.E. Stickley, J. Warnaar, G.L. Williams and M. Fuller, 2003. Dinoflagellate cysts from the Eocene-oligocene transition in the Southern Ocean: Results from ODP Leg 189. Proc. Ocean Drilling Program Scient. Results, 189: 1-42.
CrossRefDirect Link - Sowunmi, M.A., 1981. Late quaternary environmental changes in Nigeria. Pollen Spores, 23: 125-148.
Direct Link - Sowunmi, M.A., 1981. Aspects of late quaternary vegetational changes in West Africa. J. Biogeogr., 8: 457-474.
Direct Link - Sowunmi, M.A., 1999. The significance of the oil palm (Elaeis guineensis Jacq.) in the late Holocene environments of west and west central Africa: A further consideration. Veg. Hist. Archaobot., 8: 199-210.
CrossRefDirect Link - Sowunmi, M.A., 2002. Environmental and Human Responses to Climatic Events in West and West Central Africa During the Late Holocene. In: Food and Culture: Ecological Change and Food Security in Africa's Later Prehistory, Hassan, F.A. (Ed.). Kluwer Academic/Plenum Publishers, Droughts, Dordrecht, pp: 95-104.
Direct Link - Sowunmi, M.A., 2004. Aspects of Nigerian Coastal Vegetation in the Holocene: Some Recent Insights. In: Past Climate Variability through Europe and Africa, Battarbee, R.W., F. Gasse and C.E. Stickley (Eds.). Springer, Dordrecht, The Netherlands, pp: 258.
Direct Link - Udeze, C.U. and F.E. Oboh-Ikuenobe, 2005. Neogene paleoceanographic and paleoclimatic events inferred from palynological data: Cape Basin off South Africa, ODP Leg 175. Palaeogeography Palaeoclimatol. Palaeoecol., 219: 199-223.
CrossRef - Wall, D., B. Dale, G.P. Lohmann and W.K. Smith, 1977. The environmental and climatic distribution of dinoflagellate cysts in modern marine sediments from regions in the North and South Atlantic Oceans and adjacent seas. Mar. Micropaleontol., 2: 121-200.
CrossRef - Weber, K.J., 1971. Sedimentological aspects of oil fields in the Niger Delta. Geol. Mynbouw., 50: 559-576.
Direct Link - Ige, O.E., 2009. A late tertiary pollen record from Niger Delta, Nigeria. Int. J. Bot., 5: 203-215.
CrossRefDirect Link - Oboh, F.E., 1993. Depositional history of the middle miocene E2.0 reservoir in the Kolo Creek Field, Niger Delta. J. Petroleum Geol., 16: 197-212.
CrossRef