
Abhijit Sahasrabudhe
Department of Botany, Kelkar Education Trust�s V.G. Vaze College of Arts, Science and Commerce, Mithagar road, Mulund (E), Mumbai 400081, India
Manjushree Deodhar
Department of Botany, Kelkar Education Trust�s V.G. Vaze College of Arts, Science and Commerce, Mithagar road, Mulund (E), Mumbai 400081, India
International Journal of Botany
Year: 2010 | Volume: 6 | Issue: 3 | Page No.: 299-303







ABSTRACT
During ageing, elasticity of the skin decreases due to enzyme elastase leads to sagging and at the same time hyaluronic acid in the skin also diminishes and skin becomes dry and wrinkled. Hence, there is need to conserve matrix metallo proteins by inhibiting the activity of these matrix metallo proteinases. Certain plant extracts are reported to be good antioxidants. Garcinol and cambogiol present in the fruit rinds of Garcinia indica were reported to be good antioxidants due to presence of phenolic group. Along with this various preparations of G. indica have shown significant antioxidant ability. To check whether fractions separated from fruit rinds of G. indica play a role in prolonging ageing, we fractionated crude Methanolic Extract (ME) into ethyl acetate and Water Fraction (WF) and those fractions were screened for anti-hyaluronidase and anti-elastase activity. Among tested three fractions, Ethyl Acetate Fraction (EAF) at concentrations as low as 25 μg mL-1 showed significant hyaluronidase inhibition while water fraction proved to be good elastase and hyaluronidase inhibitor at 90 μg mL-1. Water fraction could be further exploited to be used in anti-ageing formulations.
PDF Abstract XML References Citation
Received: May 12, 2010;
Accepted: July 02, 2010;
Published: September 02, 2010
How to cite this article
Abhijit Sahasrabudhe and Manjushree Deodhar, 2010. Anti-hyaluronidase, Anti-elastase Activity of Garcinia indica. International Journal of Botany, 6: 299-303.
DOI: 10.3923/ijb.2010.299.303
URL: https://scialert.net/abstract/?doi=ijb.2010.299.303
DOI: 10.3923/ijb.2010.299.303
URL: https://scialert.net/abstract/?doi=ijb.2010.299.303
INTRODUCTION
Extracellular matrix (ECM) is composed of proteoglycans interwoven with matrix metallo proteins such as collagen, elastin and fibronectin. Collagen is the major structural protein provides supportive framework to the cell. Elastin maintains skin elasticity and resilience (Oikarinen, 2004). Hyaluronic Acid (HA) holds the water together and keeps body smooth, moist and lubricated.
During ageing, elasticity of the skin is decreased due to enzyme elastase which leads to sagging. At the same time, the amount of hyaluronic acid in the skin also diminishes and skin becomes dry and wrinkled. In the connective tissue fibroblasts are the cells which synthesize collagen and other important matrix metallo proteins. Proliferation and migration of these dermal fibroblasts suppresses ageing by producing more matrix metallo proteins. In young and healthy skin, the balance between synthesis and degradation of these proteins is maintained (Aslam et al., 2005). But this intricate balance gets disturbed as we age. For maintaining youthful skin, when production of metallo proteins is not enough, the best way is to conserve them and the best way to preserve them is by inhibition of matrix metallo proteinases (MMPase) (Aslam et al., 2005).
Enzyme hyaluronidase degrades hyaluronic acid by lowering its viscosity and increasing the permeability. It is found in human organs, body fluids and secretions of viruses, fungi, bacteria, leeches and also in venoms of snakes, bees, lizards, spiders etc. Hyaluronidase not only contributes in the destruction of ECM but also plays a vital role in many biological and physiological processes such as embryogenesis, angiogenesis, inflammation, disease progression, wound healing, bacterial pathogenesis, diffusion of systemic venoms and invasion of tumors (Girish and Kemparaju, 2007).
Like hyaluronidase, elastase too contributes significantly in other activities. Human neutrophil elastase (HNE) has broad substrate specificity capable of not only enzymatic cleavage of elastin but also other ECM proteins such as collagen, hyaluronan, laminin etc. Under normal physiological conditions, endogenous inhibitors protect healthy tissue from damage but when impaired balance get disturbed, it leads to variety of diseases such as lung disorders, cardiovascular disorders, arthrosclerosis and cancer etc. (Kobayashi et al., 2004).
There are certain plant metabolites have been shown to inhibit hyaluronidase and elastase. Polyphenols from blackberry, quercetein, gallotanins, tannic acid, ascorbic acid, glycyrrhizin from Glycirrizia glabra glycoproteins from Withania somniferea and certain alkaloids are good hyaluronidase inhibitors (Girish and Kemparaju, 2005) while Curcumin from Curcuma longa (Chainani-Wu, 2003), procyanidins from Vitis vinifera (Maffei-Facino et al., 1994), phenols such as epi-catechin, resveratrol (Hrenn et al., 2006) and flavonoids such as galangin, kaempferol, quercetein and myricetin are potent elastase inhibitors (Kanashiro et al., 2006). The PGG, a condensed tannin and ellagic acid are reported to bound elastin and prevent its degradation (Isenburg et al., 2007).
The plant G. indica selected for present work is endemic to Western Ghats of India. Fruits are used to make attractive red colored beverage have bilious action. It is anthelminthic and cardiotonic and used in the treatment of piles, dysentery and tumors etc. Fruit rinds of G. indica contain about 2.4% anthocyanins. Out of them two were identified as cyanindin 3-glucoside and cyanindin 3-sambuboside (Krishnamurthy et al., 1981). Garcinol, yellow colored pigment and cambogiol present in the fruit rinds showed good antioxidant activity due to the presence of phenolic group (Krishnamurthy et al., 1982) and earlier methanolic extracts of fruit rinds of G. indica showed good hyaluronidase and elastase inhibition (Sambhus, 2006). But no extensive work has done yet on hyaluronidase and elastase inhibition activities of different fractions of G. indica. The objective of the present study was to separate different fractions from crude methanolic extract of fruit rinds of G. indica and study their anti aging ability using hyaluronidase and elastase inhibition assay.
MATERIALS AND METHODS
Reagents and chemicals: This work was carried out in Department of Botany, V.G. Vaze College, Mumbai, India. The project duration was March 2006 to Feb 2009. The ripe fruits of G. indica were collected from Dr. Balasaheb Sawant Konkan Krishi Vidyapith, Dapoli, Maharashtra. The organic solvents used were of the analytical grade (Merck and Qualigen). Elastase type IV from porcine pancreas (14.1 mg/solid). N- Succinyl-Ala-Ala-Ala-p nitroanilide (SANA), Hyaluronidase Type I S from Bovine testes (7900 units mg-1 solid) and Hyaluronic acid, sodium salt from Human Umbilical cord-purchased from Sigma-Aldrich, USA. For hyaluronidase and elastase inhibition activity. Colorimetric measurements were performed on UV-visible Spectrophotometer (Varian 50 Cary).
Preparation of plant extracts
Preparation of methanolic extract: The methanolic extract was prepared by immersing (20 g) of dried fruit rinds of G. indica in 200 mL of acidified Methanol (2% Concentrated HCl). After a week the extract was poured in the evaporating dish and methanol was removed from the extract by evaporation at room temperature and dried extract was further used fractionation.
Separation of ethyl acetate fraction: One gram of ME was dissolved in 50 mL of D/W. To this 50 mL of ethyl acetate was added and two fractions were allowed to separate in a separating funnel for at least 1 h. This was repeated twice. Both the fractions were completely dried at room temperature yielding 500 mg of WF and 300 mg of EAF.
Thin layer chromatography analysis: The ME, EAF and WF were analyzed by thin layer chromatography. Solvent system standardized was Butanol: Acetic acid: water (4:1:2).
Determination of hyaluronidase inhibition: Hyaluronidase inhibition was determined by measuring the amount of N-acetylglucosamine splited from sodium hyaluronate by the method described by Lee and Choi (1999a) with slight modifications. Fifty microliter of bovine hyaluronidase (7900 units mL-1) dissolved in 0.1 M acetate buffer (pH 3.6) was mixed with 50 μL of designated concentrations of ME, EAF and WF dissolved in 5% DMSO. The control group was treated with 50 μL of 5% DMSO instead of extracts and was incubated for 20 min at 37°C. After 20 min 50 μL of calcium chloride (12.5 mM) was added to the reaction mixture and again incubated for 20 min at 37°C. This Ca2+ activated hyaluronidase was treated with 250 μL sodium hyaluronate (1.2 mg mL-1) and incubated at 37°C for 40 min. After incubation 50 μL of 0.4 M sodium hydroxide and 100 μL of 0.2 M sodium borate were added to the reaction mixture and then incubated in the boiling water bath for 3 min. After cooling to room temperature 1.5 mL of PDMAB solution (4 g PDMAB dissolved in 50 mL of 10N HCL and 350 mL of glacial acetic acid) was added to the reaction mixture that was then incubated in water bath at 37°C for 20 min when color developed. The absorbance was measured at 585 nm on Varian Cary 50 UV spectrophotometer. All the analytical data in the tables are the means of 3 determinations. Inhibitory effect was expressed as follows:
Determination of elastase inhibition: Porcine pancreatic elastase (PPE, Sigma, Type IV) was assayed spectrophotometrically using N-Succ-(Ala) 3-nitroanilide (SANA) as the substrate, monitoring the release of p-nitroaniline by the method described by Lee and Choi (1999b) with few modifications. The reaction mixture contained 800 μL of 0.2 M Tris buffer (pH 8.0), 100 μL of enzyme elastase and 100 μL of 0.8 mM succ-(Ala) 3-nitroanilide as substrate and different concentrations of plant extracts in Tris- HCl buffer. Extract was preincubated with enzyme for 20 min at 25°C and the reaction was started with the addition of substrate. In control, buffer was used in place of the extracts. The absorbance was monitored at 410 nm for 20 min using UV spectrophotometer. As a positive control we used A.catechu extract. Each treatment was replicated thrice. Inhibitory effect of the extracts on the Elastase activity was calculated as:
Statistical analysis: All assays were performed at least three times. Statistical analysis was performed using one way ANOVA followed by * which is significant at p< 0.05 (n = 3). Values given are Mean±SD.
RESULTS AND DISCUSSION
Thin layer chromatography: Methanolic extract comprised of many compounds that appeared to be violet, red, yellow and brown spots on TLC plates (Fig. 1). When methanolic extract was fractioned into EAF and WF, ethyl acetate fraction comprised of mainly yellow and brown pigments and water fraction contained majority of all pink and red colored pigments.
Hyaluronidase inhibition: Methanolic extract at low concentrations, i.e., 250 μg mL-1 failed to produce noticeable hyaluronidase inhibition. Concentrations in the range of 500 to 750 μg mL-1 produced significant inhibition (Table 1). At 750 μg mL-1 it was 94% and then there was a decline at higher concentrations.
 | |
| Fig. 1: | Thin layer chromatogram of fractions separated from fruit rinds of G. indica |
| Table 1: | Effect of various concentrations of ME separated from fruit rinds of G. indica on hyaluronidase inhibition |
 | |
| Each value is expressed as Mean±SD of three replicate determinations. Numbers followed by * are statistically significant at p<0.05 according to ANOVA (n = 3) | |
| Table 2: | Effect of various concentrations of EAF and WF separated from fruit rinds of G. indica on hyaluronidase inhibition |
 | |
| Each value is expressed as Mean±SD of three replicate determinations. Numbers followed by * are statistically significant at p<0.05 according to ANOVA (n = 3) | |
When ME was fractioned into EAF and WF, EAF at very low concentrations, i.e., 25 μg mL-1 produced significant inhibition (88.33%) while water fraction failed to produce any inhibition at such low concentrations. Ethyl acetate fraction at a very broad range of concentrations 25 to 500 μg mL-1 produced significant hyaluronidase inhibition. Water fraction from concentrations 125 μg mL-1 showed effective inhibition (Table 2).
Elastase inhibition: In case of methanolic extract, the optimum range of elastase inhibition was 150 to 200 μg mL-1. At 200 μg mL-1 inhibition was 64.02%. After fractionation, EAF up to 150 μg mL-1 didn’t produce any significant inhibition. At 250 μg mL-1 it was 73.02%. On the other hand, water fraction even at very low concentrations, i.e., 100 μg mL-1 onwards inhibited elastase (Fig. 2). Significant inhibition was seen at 200 μg mL-1 (81.02%) as compared to A. catechu which had 59% inhibition at same concentration.
Hyaluronidase mediated degradation of HA increases the permeability of the connective tissue and decreases the viscosity of the body fluids and is involved in the bacterial pathogenesis, spread of toxins and venoms. It facilitates the entry of sperm during invitro fertilization (IVF) and also involved in cancer progression.
Phenolic compounds such as quercetin, tannic acid acts as the inhibitors of venom hyaluronidase (Girish and Kemparaju, 2005) or inhibit sperm hyaluronidase (Garg et al., 2005). Compounds like Apigenin, is known to inhibit tumor induced angiogenesis (Lokeshwar et al., 2006). Similarly elastase released by neutrophils is physiologically controlled by specific endogenous inhibitors such as α-antitrypsin, elafin etc.
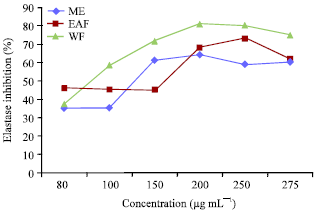 | |
| Fig. 2: | Elastase inhibition assay of ME, EAF and WF separated from the fruit rinds of G. indica |
The imbalance between elastase and its inhibitors is the cause of many inflammatory diseases such as rheumatoid arthritis, pulmonary emphysema and acute lung injury. As mentioned earlier, Pentagalloyl glucose (PGG), condensed tannin is reported to bind with enzyme elastase and render resistance to elastase inhibitors in arotic aneurysm and thus can be used as effective therapy in cardiovascular disorders. The above description suggests the necessity of inhibition of these two enzymes not only to prolong ageing but also to prevent the risk of such fetal disorders.
As mentioned earlier, there are certain phenols and flavonoids which inhibit hyaluronidase and elastase in dose dependant manner. Lee et al. (2001) showed that phenolic substances isolated from A.catechu inhibited elastase significantly (IC50 ~ 26.92 μg mL-1). Flavonoids and triterpenoids separated from Cornus kousa showed more than 50% elastase inhibition at 100 μg mL-1 while (IC50 ~ 21.21 μg mL-1) (Lee and Sultana, 2007). Similarly flavonoids of Achillea millefolium reported to be good elastase inhibitors. There are various reports where polyphenols such as tannic acid, apigenin and quercetin inhibited hyaluronidase activity significantly (Girish and Kemparaju, 2005). The above mentioned examples proved that polyphenols and flavonoids could be used as potent antioxidants in anti ageing formulations.Here in the present work we found out that crude methanolic extract at low concentrations failed to inhibit hyaluronidase but was effective in bringing out elastase inhibition. After fractionation into EAF and WF, EAF even at very low concentrations inhibited hyaluronidase significantly (at 25 μg mL-1 it was 88.03% inhibition) while WF didn’t had any effect on hyaluronidase activity at such low concentrations. On the other hand, for elastase inhibition, WF was more effective than that of EAF. The WF at 200 μg mL-1 produced significant elastase inhibition (81.02%) with that of A. catechu (used as the positive control) had only 59% at same concentration.
Kumar et al. (2007) fractionated methanolic extract of G. indica in to hexane, chloroform and ethyl acetate fraction and showed that ethyl acetate fraction is the most toxic fraction against various cell lines. Further they isolated two benzophenones namely xanthochymus and isoxanthochymus from ethyl acetate fraction and studied their cytotoxicity.
CONCLUSION
This study reveals that among tested fractions, water fraction could be used as potential antioxidant and efforts are already underway to isolate and identify the compounds of water fraction which are responsible for bringing out such significant hyaluronidase and elastase inhibition and at the same time it will be less toxic to dermal fibroblasts.
REFERENCES
- Aslam, M.N., E.P. Lansky and J. Varani, 2006. Pomegranate as a cosmeceutical source: Pomegranate fractions promote proliferation and procollagen synthesis and inhibit matrix metallo proteinases-1 production in human skin cells. J. Ethnopharmacol., 103: 311-318.
CrossRef - Chainani-Wu, N., 2003. Safety and anti-inflammatory activity of curcumin: A component of tumeric (Curcuma longa). J. Altern. Complementary Med., 9: 161-168.
CrossRefDirect Link - Maffei-Facino R., M. Carini, G. Aldini, E. Bombardelli, P. Morazzoni and R. Morelli, 1994. Free radicals scavenging action and anti enzyme activities of procyanidins from Vitis vinifera: A mechanism for their capillary protective action. Arzneimittelforschung, 44: 592-601.
PubMedDirect Link - Garg, A., R. Anderson, L. Zaneveld and S. Garg, 2005. Biological activity assessment of a novel contraceptive anti microbial agent. J. Androl., 26: 414-421.
CrossRefDirect Link - Girish, K.S. and K. Kemparaju, 2005. Inhibition of Naja naja venom Hyaluronidase by plant derived bioactive components and polysaccharides. Biochemistry, 70: 948-952.
PubMedDirect Link - Girish, K.S. and K. Kemparaju, 2007. The magic glue hyaluronan its eraser hyaluronidase: A biological overview. Life Sci., 80: 1921-1943.
CrossRefDirect Link - Hrenn, A., T. Steinbrecher, A. Labahn, J. Schwager, C. Schempp and I. Merfort, 2006. Plant phenolics inhibit neutrophil elastase. Planta Med., 72: 1127-1131.
PubMedDirect Link - Isenburg, J.C., D.T. Simionescu, B.C. Starcher and N.R. Vyachare, 2007. Elastin stabilization for treatment of abdominal arotic aneurysms. Circulation, 115: 1729-1737.
Direct Link - Kanashiro, A., J.G. Souza, L.M. Kabeya, A. Elisa, C.S. Azzolini and M. Lucisano-Valim, 2006. Elastase release by stimulated neutrophils inhibited by flavonoids: Importance of the catechol group. Z. Naturforsch., 62: 357-361.
Direct Link - Kim, Y.J, H. Uyama and S. Kobayashi, 2004. Inhibition effects of (+)-catechin-aldehyde polycondensates on proteinases causing proteolytic degradation of extracellular matrix. Biochem. Biophys. Res. Commun., 320: 256-261.
CrossRefPubMedDirect Link - Kumar, S., S.K. Chattopadhayay, M.P. Darokar, A. Garg and S.P. Khanuja, 2007. Cytotoxic activities of xanthochymol and isoxanthochymol substantiated by LC-MS/MS. Planta Med., 73: 1452-1456.
PubMedDirect Link - Lee, K.K. and J.D. Choi, 1999. The effects of Areca catechu L extracts on anti inflammation and anti melanogenesis. Int. J. Cosmet. Sci., 21: 275-284.
CrossRefDirect Link - Lee, K.K. and J. Choi, 1999. The effects of Areca catechu L extracts on anti ageing. Int. J. Cosmet. Sci., 21: 285-295.
CrossRefPubMedDirect Link - Lee, K.K., J.J. Cho, E.J. Park and J.D. Choi, 2001. Anti-elastase and anti-hyaluronidase of phenolic substance from Areca catechu as a new anti-ageing agent. Int. J. Cosmet. Sci., 23: 341-346.
CrossRefPubMedDirect Link - Sultana, N. and N.H. Lee, 2007. Antielastase and free radical scavenging activities of compounds from the stems of Cornus kousa. Phytother. Res., 21: 1171-1176.
PubMed - Isoyama, T., D. Thwaites, M.G. Selzer, R.I. Carey, R. Barbucci and V.B. Lokeshwar, 2006. Differential selectivity of hyaluronidase inhibitors toward acidic and basic hyaluronidases. Glycobiology, 16: 11-21.
CrossRefDirect Link - Oikarinen, A., 2004. Connective tissue and ageing. Int. J. Cosmet. Sci., 26: 107-108.
CrossRefDirect Link