
K. Beena Anto
Division of Plant Physiology and Biochemistry, Department of Botany, University of Calicut Malappuram Dt., Kerala-673635, India
K. M. Jayaram
Division of Plant Physiology and Biochemistry, Department of Botany, University of Calicut Malappuram Dt., Kerala-673635, India
International Journal of Botany
Year: 2010 | Volume: 6 | Issue: 2 | Page No.: 122-126







ABSTRACT
The present investigation was undertaken to study correlation of seed water content and viability with its seed and seedling vigour in the temperature-treated seeds of pea and soybean during the early stages of germination. Seeds could withstand high temperature up to 70°C for 10 days continuously but high temperature treatment reduced the rate of germination percentage, moisture content as well as seed and seedling vigour. High temperature induced certain changes on pea seeds and resulted in the appearance of hard seeds. This effect was not observed in the soybean seeds due to oil-rich nature.
PDF Abstract XML References Citation
How to cite this article
K. Beena Anto and K. M. Jayaram, 2010. Effect of Temperature Treatment on Seed Water Content and Viability of Green Pea (Pisum sativum L.) and Soybean (Glycine max L. Merr.) Seeds. International Journal of Botany, 6: 122-126.
DOI: 10.3923/ijb.2010.122.126
URL: https://scialert.net/abstract/?doi=ijb.2010.122.126
DOI: 10.3923/ijb.2010.122.126
URL: https://scialert.net/abstract/?doi=ijb.2010.122.126
INTRODUCTION
It is well known that physiological activity in seeds is affected by water content (Rupley et al., 1983; Vertucci et al., 1985). In seeds water binds with varying strengths at different water concentrations and therefore has different thermodynamic properties (Rupley et al., 1983). The occurrence of 3 different types of water has been reported in dry seeds by various researchers (Vertucci and Leopold, 1984; Vertucci et al., 1985). The physical state of water in seeds determines the physiological manifestations connected with imbibition and germination. Ishida et al. (1987) reported that most of the biological activities cease or at minimum level in dry seeds including food consumption.
Moisture content reduction in seeds is initiated during maturation of seeds and orthodox seeds can be dried to low moisture content without any damage (Bewley and Black, 1994; Mayer and Poljakoff-Mayber, 1989; Bakker, 2001).
When the seeds are subjected to higher temperature, progressive removal of water occurs. Bound water, associated with macromolecules is lost first, resulting in structural and functional deterioration of seeds (Bewley and Black, 1994). Higher temperature induces tighter packing of molecules and increases their structural disorder leading to loss of seed vigour and/or viability. The final water content of dry seeds appears to be important in determining their survival over long periods of storage (Robert and Ellis, 1989).
Seed viability denotes the degree to which a seed is alive, metabolically active and possesses enzymes necessary for catalyzing metabolic reactions needed for germination and seedling growth (Basara et al., 2002). According to Gelmond et al. (1978) the seed vigour means a high rate of the overall biological activities of the seed, resulting in a high yield performance. They measured and predicted seed vigour according to the rate of root emergence of germination or field emergence. Vigour represents the potential ability of the seed to yield the maximum plant product at the earliest time under variable environmental field conditions.
In the present study, an attempt was made to correlate how temperature treatment and seed water content affected the viability of pea and soybean seeds.
MATERIALS AND METHODS
Seeds of pea (Pisum sativum L.) c.v. boneville and soybean (Glycine max L. Merr.) c.v. SL. 525 were used to study the correlation of germination percentage, moisture content and seed and seedling vigour. From the seed lots, seeds having uniform size, colour and shape with intact seed coat were selected and were subjected to temperature treatments such as 50, 60 and 70°C for 10 days each.
The temperature treated and fresh, healthy, untreated seeds, used as control were kept in Petri dishes for germination under laboratory conditions and germination count was taken daily up to 7 days. Parameters like moisture content, imbibition percentage, percentage of hard seeds, germination percentage and seed/Seedling Vigour Index (SVI) were calculated (ISTA, 1985; Copeland and McDonald, 1995).
Germination percentage: Three replicates of 30 seeds each of treated and untreated or control seeds were placed in sterilized Petri dishes and covered with lid plates which also lined with moistened filter paper. Petri dishes were watered as required to replace evaporation losses. Radicle emergence of 1 mm was scored as germinated and germination percentage was calculated by using the formula:
 |
Moisture content: Thirty seeds from each of the treatment and control were taken and brought to room temperature by keeping in a desiccator and fresh weight of these seeds was determined by using electronic balance (Shimadzu AX 120). Then the weighed seeds were kept in a hot air oven at 100°C for 1 h and kept at 60°C until constant weight was obtained. Percentage of moisture content was calculated, as explained by ISTA (1985):
Seed Vigour Index (SVI): Thirty seeds each in triplicate from the control and temperature treated seed lot were sampled and sown in garden pots filled with garden soil, sand and dry powdered cow dung mixed in 2:1:1 ratio. Daily count of germinated seeds was taken and percentage of germination was calculated. The seed vigour index was calculated according to Copeland and McDonald (1995) using the formula given below:
 |
Seedling vigour: Vigour of seedlings of control and temperature treated pea and soybean was calculated using biomass method. Seedlings after 7 days of germination were uprooted carefully and washed in running tap water to remove the sand particles. The seedlings with cotyledon were blotted and weighed using electronic balance. After noting the initial weight, the seedlings were kept in hot air oven at 100°C for 1 h and transferred to and kept in 60°C until constant weight was obtained. The seedling vigour was expressed as biomass per seedling.
RESULTS
Seeds of P. sativum and G. max are orthodox and the seed lots exhibited 100% germination. When the seeds were subjected to temperature treatments at 50, 60 and 70°C for 10 days continuously, the germination percentage was decreased (Table 1). The high temperature up to 70°C given to the seeds reduced the germination percentage to 10% in pea and 17% in soybean.
Viability of the two legume seeds, P. sativum and G. max is inversely proportional to temperature and above 50°C, the germination percentage is below 50%. Soybean seeds exhibited higher resistance to temperature stress with a higher germination percentage and seed vigour index values than the pea seeds.
For the maintenance of 100% germination, 12.8 and 13.9% of moisture content are found to be essential in pea and soybean seeds, respectively. When the moisture content is reduced to 7.3% in pea and 8.7% in soybean at 60°C, the germination is reduced to 33 and 40%, respectively and this reduction in germination is directly proportional to the temperature (Table 1).
Viability of the two legume seeds, P. sativum and G. max is inversely proportional to temperature and above 50°C; the germination percentage is below 50%. The germination percentage declined to 10 from 33% when the temperature was raised from 60 to 70°C.
The germination is found delayed in both the seed samples subjected to temperature treatment and soybean exhibited more seed vigour in samples treated at 70°C than those of pea seeds.
| Table 1: | Effect of Temperature on water content and viability of Pisum sativum and Glycine max seeds |
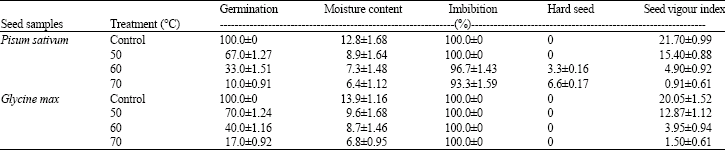 | |
In the present study, when the seeds were dried gradually, more time was taken for imbibition as well as germination and this was reflected in the reduced values of seed vigour index. An important observation in pea seeds was that, the percentage of hard seeds treated at 60°C was 3.3% and at 70°C, the percentage was increased to 6.6% (Table 1). But soybean seeds showed no hardness in any of the temperature treatments.
DISCUSSION
As a consequence of high temperature treatment, reduction in germination has been reported in Solanum nigrum (Del Monte and Tarquis, 1997) and in Raphanus sativum (Meng et al., 2003) seeds. Roberts (1973) opined that orthodox seeds can be dried to low moisture contents without damaging the embryo and their longevity increases with decrease in moisture content during storage over a wide range of conditions. Contradictory to this view, drying at high temperature leads to significant reduction of moisture content and concomitant loss of viability in pea and soybean seeds, plausibly due to the continuous treatment for 10 days at 50 to 70°C (Table 1).
It can be presumed that during this temperature treatment, the tissues lose the water molecules adsorbed to the molecules of cell membranes. This is in consonance with the views of Bewley and Black (1994), who suggested that the adsorbed water or bound water is loosely held by bonding to macromolecules.
The reduction of moisture content below 6.4 and 6.8%, respectively in pea and soybean seeds, when treated at 70°C, resulting in the loss of viability presumably reveals the loss of all the three types of water as suggested by Bewley and Black (1994). However, temperature treatment at 50°C showed above 60% germination, which may be due to the loss of free water alone. The metabolic activities of the seeds at this temperature treatment could be maintained almost normal even in the absence of free water. Since, the rate of viability loss in P. sativum and G. max seeds as a response to higher temperature treatment and to the rate of water loss is gradual and more or less continuous, the concepts of bound water cannot be directly correlated to the reduction of moisture content in these seeds at any particular temperature. It seems that the entire bound water or type III of bound water is not completely removed at 70°C because viability is still retained at least 10% in pea and 17% in soybean.
Bewley and Black (1994) suggested that water content of seeds exists in a glassy (vitrified) state even at physiological temperatures and it stops or slows down chemical reactions, assures stability and quiescence and prevents interactions of cell components. In vitro studies have shown that vitrification of water retards or prevent denaturation of proteins including enzymes (Bruni and Leopold, 1991). So, in the present study, it can be presumed that even the vitrified water in P. sativum and G. max seeds is being slowly removed by temperatures higher than 50°C, resulting in gradual loss of viability.
The reason for the viability of seeds being maintained at high temperatures like 70°C may be at least partial maintenance of vitrification or glass formation of water within the cytoplasm, which is the potential mechanism to avoid crystallization of proteins and solutes present in the cytoplasm as suggested by Bruni and Leopold (1991). Williams and Leopold (1989) explained the glassy nature of water in corn embryos as a liquid with the viscosity of a solid and its formation from a liquid involves no chemical and physical change in the solution. The major function of the glassy state in the dry seeds is its contribution to the stability of the seed components during storage and thus to the survival during desiccation (Leopold et al., 1994).
According to Nichols and Heydecker (1968), the rate or speed of germination is considered as the criteria for seed vigour index determination. The time taken for imbibition/germination is also very important. The decline of seed vigour index in P. sativum and G. max seeds is closely correlated with the loss of moisture content which is the determinant factor of seed quality. This observation is in conformity with the view of Fu et al. (1994), who suggested that seed vigour index is an accurate measure for testing quality of seeds.
A close correlation between moisture content, germination percentage and seed vigour index was observed in both the legume seeds studied. The temperature treatments made the seeds more desiccated, resulting in reduction of seed vigour index, germination percentage and moisture content percentage throughout the period of study. According to Harrington (1972), Villiers (1973) and Douglas (1975), seed vigour is affected by various environmental factors, such as temperature, moisture content and concentrations of oxygen and carbon dioxide.
In the present study, soybean seeds showed higher resistance to temperature stress with a higher germination percentage and seed vigour than the pea seeds. The oil-rich nature of soybean seeds can be correlated to these qualities (Table 1). This observation is in agreement with the view of Pixton (1967) and according to the researcher, oil content influences seed water relations and is a major determinant of equilibrium and relative humidity, which in turn are related to water potential. Roberts and Ellis (1989) and Ellis et al. (1989) reported the evidence for a common response of seed longevity to moisture content in oily and non-oily seeds.
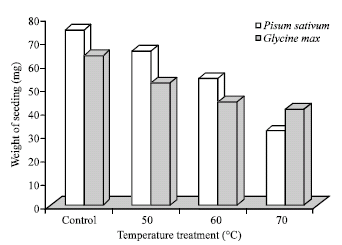 | |
| Fig. 1: | Effect of temperature on seedling vigour in Pisum sativum and Glycine max temperature treatment |
The effect of temperature treatment on pea and soybean seeds reflected on the rate of germination and growth of seedlings up to 7 days showed that the seedling vigour was gradually decreased proportional to the increase in temperature (Fig. 1).
Even when conditions are apparently favorable for germination, high temperature treatment induced dormancy. Ellis and Roberts (1982) suggested that hard seeded condition is an important consequence of desiccation. Hard seeded nature has been attributed to both genetic factors and environmental conditions such as soil fertility, photoperiod, relative humidity and temperature (Bewley and Black, 1983; Mayer and Poljakoff-Mayber, 1989). So the difference in the distribution of hard seeded condition between pea and soybean seeds treated to similar higher temperature regimes is presumably due to the genetic factors.
Hard seeds achieve and maintain a very low percentage of moisture, despite wide fluctuations in the moisture contents of the surrounding air. Mai-Hong et al. (2003) observed that in seeds of Peltophorum pterocarpum, a tree legume, the hard-seededness was induced when seeds were dried to about 15% moisture content. Storage of seeds for extended period of time at high temperature and humidity resulted in the occurrence of increased hardness in most legumes.
In legume seeds like pea, storage brings about hardening of the seed coat by oxidation of the phenolic compounds present in the testa (Van Staden et al., 1989). According to those researchers, the oxidation of phenolic compounds blocks the small pores in the seed coat and thereby makes the seeds impermeable to water. Therefore, a large percentage of hard seeds were seen in stored samples for a long period. However, in the present study, instead of prolonged storage, high temperature treatment results in enhanced oxidation of phenolics of the testa of pea seeds. In Glycine max seeds, there is no sign of decolorization or browning of the seed coat and therefore no hard seeds were observed and all the seeds imbibed readily and uniformly when soaked in water.
ACKNOWLEDGMENTS
The first author (KBA) is thankful to the Head of the Department of Botany, University of Calicut for providing necessary facilities and the U.G.C. for granting financial assistance to carry out the study.
REFERENCES
- Basara, S.M.A., N. Ahmad, K. Rehman and N. Iqbal, 2002. Cotton seed invigoration by pre-sowing seed humidification. Int. J. Agric. Biol., 4: 127-130.
Direct Link - Bakker, J., 2001. Seeds, ecology, biogeography and evolution of dormancy and germination. C.C. Baskin and J.M. Baskin J. Plant Ecol., 152: 204-205.
CrossRefDirect Link - Bruni, F. and A.C. Leopold, 1991. Glass transition in soybean seed: Relevance to anhydrous biology. Plant Physiol., 96: 660-663.
Direct Link - Del Monte, J.P. and A.M. Tarquis, 1997. The role of temperature in the seed germination of two species of the Solanum nigrum complex. J. Exp. Bot., 48: 2087-2093.
Direct Link - Ellis, R.H., T.D. Hong and E.H. Roberts, 1989. A comparison of the low moisture content limit to the logarithmic relation between seed moisture and longevity in twelve species. Ann. Bot., 63: 601-611.
Direct Link - Fu, J.R., J.P. Jin, Y.F. Peng and Q.H. Xia, 1994. Desiccation tolerance in two species with recalcitrant seeds: Clausena lansium (Lour.) and Litchi chinensis (Sonn.). Seed Sci. Res., 4: 257-261.
CrossRefDirect Link - Gelmond, H., I. Luria, L.W. Woodstock and M. Perl, 1978. The effect of accelerated ageing of Sorghum seeds on seedling vigour. J. Exp. Bot., 29: 489-495.
Direct Link - Ishida, N., H. Kano, T. Kobyashi, H. Hamaguchi and T. Yoshida, 1987. Estimation of biological activities by NMR in soybean seeds during maturation. Agric. Biol. Chem., 51: 301-307.
Direct Link - Leopold, A.C., W.Q. Sun and I. Bernal-Lugo, 1994. The glassy state in seeds: Analysis and function. Seed Sci. Res., 4: 267-274.
CrossRefDirect Link - Mai-Hong, R., T.D. Hong, N.T. Hien and R.H. Ellis, 2003. Onset of germinability, desiccation tolerance and hardseededness in the seeds of Peltophorum pterocarpum (DC) K. Heyne. (Caesalpinioideae). Seed Sci. Res., 13: 323-327.
CrossRefDirect Link - Nichols, M.S. and W. Heydecker, 1968. Two approaches to the study of germination data. Proc. Int. Test Assoc., 33: 531-540.
Direct Link - Pixton, S.W., 1967. Moisture Content-Its significance and measurement in stored products. J. Stored Prod. Res., 3: 35-47.
CrossRef - Roberts, E.H., 1973. Predicting the storage life of seeds. Seed Sci. Technol., 1: 499-514.
Direct Link - Vertucci, C.W. and A.C. Leopold, 1984. Bound water in soybean seed and its relation to respiration and imbibitional damage. Plant Physiol., 75: 114-117.
Direct Link - Vertucci, C.W., J.L. Ellenson and A.C. Leopold, 1985. Chlorophyll fluorescence characteristics associated with hydration level in pea cotyledons. Plant Physiol., 79: 248-252.
Direct Link - Williams, R.J. and A.C. Leopold, 1989. The glassy state in corn embryos. Plant Physiol., 89: 977-981.
PubMedDirect Link