
D. M. Musyimi
Department of Botany and Horticulture, Maseno University, P.O. Box 333, Maseno, Kenya
LiveDNA: 254.15012
G. W. Netondo
Department of Botany and Horticulture, Maseno University, P.O. Box 333, Maseno, Kenya
G. Ouma
Department of Botany and Horticulture, Maseno University, P.O. Box 333, Maseno, Kenya
International Journal of Botany
Year: 2007 | Volume: 3 | Issue: 1 | Page No.: 78-84







ABSTRACT
This study was conducted to investigate growth and gas exchange characteristics of avocado seedlings growing under different salinity levels under naturally illuminated greenhouse conditions, in order to relate this physiological information to the ecology of this avocado cultivar. Plants grown in 4.5 L plastic pots containing soil were subjected to 0 (control), 15, 30, 45 and 60 mM NaCl salinity treatments. The measured parameters started to show significant differences (p≤0.05) by day 7. Interactions between salt treatments and duration of salt exposure were highly significant at p≤0.05. Net photosynthetic rate (PN), stomatal conductance (gs), transpiration rate (E) and chlorophyll (chl) concentration decreased in response to increasing salt concentration in the growth medium. Water use efficiency (WUE) decreased in all the treatments during the time course of experiment. Substomatal CO2 concentration (Ci) and chloride ions content increased with increasing salt concentration of the growth medium. It is suggested that the greater inhibition of CO2 fixation may be due to impairment of photosynthetic apparatus. Treatment consisting of 60 mM NaCl caused maximum growth reductions. The findings in this study demonstrate that NaCl salinity hampers growth and gas exchange processes of avocado plants and improvement in salt resistance of avocado rootstock under study is more likely to come from increasing further screening of more avocado rootstocks for salt tolerance or resistance.
PDF Abstract XML References Citation
How to cite this article
D. M. Musyimi, G. W. Netondo and G. Ouma, 2007. Effects of Salinity on Growth and Photosynthesis of Avocado Seedlings. International Journal of Botany, 3: 78-84.
DOI: 10.3923/ijb.2007.78.84
URL: https://scialert.net/abstract/?doi=ijb.2007.78.84
DOI: 10.3923/ijb.2007.78.84
URL: https://scialert.net/abstract/?doi=ijb.2007.78.84
INTRODUCTION
A rapidly increasing world population and the fast approaching geographical limitations of the world agriculture system have led to a serious consideration for a possible expansion of agricultural activities mn to marginal lands which are unsuitable for plant growth and development. As with most types of environmental stresses, the assessment of the tolerance level of the different processes of developing salt stress is a complex task and fundamental to establishing criteria for action in such situations. The thrust of the study reported here, is to identify the physiological and cellular mechanisms utilized by the avocado plants to adapt to saline environments, so that a rational basis may be formed to develop salt tolerant avocados. For example, growth and reproduction can be altered in different ways in the same plant, as the shoot is usually more salt sensitive than the root (Munns and Termaat, 1986). Salt tolerance refers to the ability of plants to maintain growth in saline environments (Mickelbart and Arpaia, 2002). Salt stress is one of the major environmental stresses that cause decreases in growth and photosynthesis (Netondo et al., 2004a). Three major hazards associated with salinity are: - osmotic stress, ion toxicity and mineral deficiencies (Reinhardt and Rost, 1995; Hasegawa et al., 2000; Netondo et al., 2004a). Reduction in photosynthesis is directly related to stomatal conductance, though non-stomatal factors are also associated with lower photosynthetic capacity in salt treated plants (Ashraf et al., 2002; Netondo et al., 2004b). Growth and photosynthesis are particularly important under saline conditions since resistance to external salinity is influenced by plant vigour. The more vigorous the plant growth under non-saline conditions, the greater is its resistance to salt (Flowers et al., 1988). Photosynthetic performance in plants is usually enhanced by additional environmental factors such as high irradiance, water availability and soil fertility (Jimenez et al., 1997; Hofshi, 1998; Shalhevet, 1999).
Avocado (Persea americana Mill.) is a salt sensitive tree crop (Hofshi, 1995; Hofshi, 1998; Crowley and Smith, 1999; Mickelbart and Arpaia, 2002) and is often grown in areas of relatively low rainfall and saline soils (Branson and Gustafson, 1972). Growth rates of avocado trees on avocado rootstocks have been shown to reduce during an active growth flush under salinity stress (Mickelbart and Arpaia, 2002). Earlier research by Mickelbart and Arpaia (2002) has indicated that sensitivity to salinity among avocado cultivars was reflected in different growth reductions and leaf necrosis. Schaffer and Whiley (2003) have indicated that stomatal conductance is a more reliable early indicator of stress in avocado than measurements of leaf water content, leaf water potential or growth variables. The importance to plant breeding of knowledge of the physiology is important because characters conferring adaptability in an ecological sense are not necessarily of agricultural benefit (Turner, 1986). Although physicochemical responses of crop plants to environmental stresses have been studied, relatively little is known about avocado rootstocks in Kenya. There is little information available on the possible interaction between salinity and PAR on gas exchange and water use efficiency of avocado rootstocks. The current interest in utilization of saline soils and saline water in arid areas necessitates knowledge of growth and gas exchange characteristics for this fruit tree under salinity stress, which is a suitable candidate for incorporation into agroforestry systems.
The main objective of this study was to evaluate the influence of salinity stress on growth, PN, gs, E, WUE and chl concentration in young avocado plants as part of study to determine the combined effects of salinity and low PAR irradiance. Data obtained may explain physiological mechanisms by which salinity affects growth and development in avocado and may help improve the management of avocado productivity in saline environments and agroforestry systems. The results should also help to form the basis for the breeding of salt tolerant avocado plants in Kenya.
MATERIALS AND METHODS
Plant material and growth conditions: Three-month-old avocado plants (Persea americana Mill.) were raised inside a naturally illuminated greenhouse whose conditions during the experiment were: temperature: min/max 20/41 °C, relative humidity: min/m`x. 50/95%. The CO2 concentration in the greenhouse was not controlled. The plants were selected on the basis of uniformity of size and transplanted in 4.5 L plastic pots filled with local soil classified as kandiudalfic Eutrodox (USDA, 1992). The exposed soil had been covered with aluminium foil to prevent growth of algae. The mineral fertilizer used was 20 g of Di-ammonium Phosphate (DAP) per pot at planting. Plants were separated into five lots of 4 plants each; with one lot as control, (0 mM NaCl) and subjecting the other four to different salinity treatments of 15, 30, 45 and 60 mM NaCl. The saline treatments were administered in a step-wise fashion, adding daily increments of 300 mL of 15 mM saline water until the desired concentration was reached. The application rate of saline solution was adequate to ensure more than 30% drainage of applied solution through perforations at the bottom of the pots. The pots were arranged in a completely randomized design on a bench. Weeds were controlled by hand pulling, while recommended pesticides were used to control pests.
Plant growth measurements: The data for growth parameters were recorded once every week before and after the commencement of the salinity treatments. Shoot height was measured using a meter rule, from the stem base up to the shoot apex. The number of fully expanded mature leaves per plant on the main stem and branches were counted and recorded. The change in growth of the stem diameter was determined in each plant by measuring the diameter at a height of 10 cm from the stem base using a vernier caliper. All the above measurements were done once every week. At the end of the experiment the plants were harvested and their roots and shoot regions were separated. Roots were rinsed in tap water after soaking, blotted dry on paper towels and weighed using an electronic weighing balance (Denver Instrument Model XL-3100D). Fresh weight reading for the shoot was taken immediately after harvesting. All the plant samples were then oven-dried at 60°C to constant dry weights, for at least 48 h after which time their dry weights were determined.
Gas exchange measurements: An open infrared gas analyzer system in which the CO2 concentration of in- and outcoming air was measured differentially (CIRAS-1, PP Systems, Stotfield, Hitchin, Herts, UK) was used to measure gas exchange parameters (PN, gs, E and Ci). Gas exchange was determined from an area of 2.5 cm2 of the fully expanded sun-exposed fifth leaf (from the shoot apex) of the plant in each treatment between 0900 and 1230 h. Photosynthetic rates were measured at 26 to 37°C. The Photosynthetically Active Radiation (PAR) measured at the leaf surface was 120-200 μmol m-2sec¯1.The vapour pressure deficit within the leaf cuvette was maintained throughout these experiments, at 0.5-0.7 kPa using this system. The air flow rate through the cuvette was 200 mL min-1. Ten consecutive measurements were taken at 3 sec intervals. Measurements were made in-doors and commenced on the seventh day after commencement of salt treatment and were done once per week.
Water use efficiency: Water use efficiency was calculated using the formula of Ashraf et al. (2002), according to the following equation.
Water Use Efficiency (WUE) = PN / E, where, PN = net photosynthetic rate and E = transpiration rate
Chlorophyll concentration: The fourth fully expanded leaves were harvested at the end of the experiment. The chlorophyll concentration was determined in 80% acetone extract on a spectrophotometer (Model Novaspec II, Pharmacia Biotech, Cambridge, England). Absorbency was measured against an 80% acetone blank at 645 nm and 663 nm. The tchl (mg g-1) was obtained by summation of the calculated values of chl a and chl b following the formulae of Arnon (1949).
Leaf chloride ion: 0. 1 g of finely ground oven dried tissue was digested over night with 25 cm3 of 0.1M HNO3 at room temperature according to Sibole et al. (2003). Chloride content was determined from the aqueous extract by titration with silver nitrate. Ion concentrations were calculated on a tissue basis from the dry masses of the same leaf. 25 cm3 of the aqueous extract was used to titrate with 0.1M Ag NO3.
Statistical analysis: All data were subjected to analysis of variance (ANOVA) using SAS statistical computer package to test the significance of treatments.
RESULTS
Plant growth: There were no significant (p≥0.05) differences in shoot height growth between salinity treatments (Table 1a). Salinity stimulated stem diameter growth in the first few days after initiation of salt treatments. Salt treated seedlings had significantly (p≤0.05)) fewer number of leaves than control plants. Growth at high salinity resulted in large reductions in fresh and dry weight production of both shoot and root (Table 1b). The reduction in shoot dry weight was attributed to lower leaf number and development of smaller leaves with increased salinity of the growth medium.
| Table 1a: | Analysis of growth parameters after 39 days of saline water irrigation |
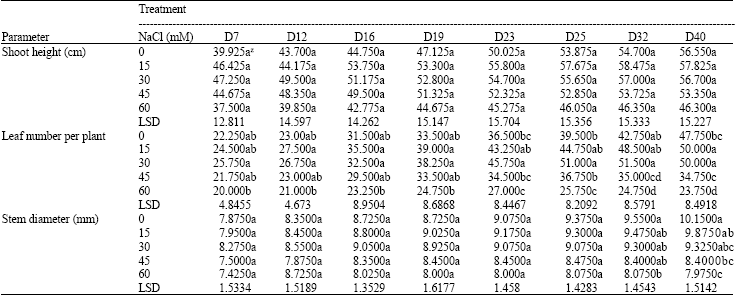 | |
| zLetter(s) show significant differences at p<0.05 with t-test. Data were log-transformed for analysis and back-transformed for presentation | |
| Table 1b: | Analysis of growth parameters after 39 days of saline water irrigation |
 | |
| zLetter(s) show significant differences at p<0.05 with t-test. Data were log-transformed for analysis and back-transformed for presentation | |
| Table 2: | Analysis of means of chlorophyll concentration and chloride content after 39 days of saline water treatment |
 | |
| zLetters show significant differences at p<0.05 with t-test. Data were log-transformed for analysis and back-transformed for presentation | |
| Table 3: | Analysis of means of net photosynthetic rate and related parameters |
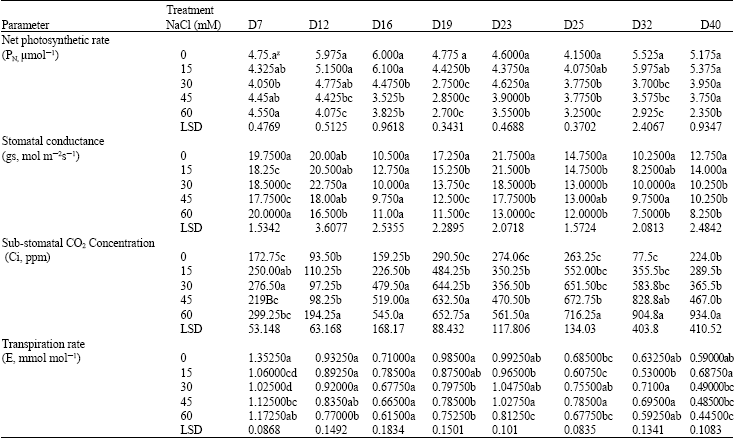 | |
| zLetter(s) show significant differences at p<0.05 with t-test, Data were log-transformed for analysis and back-transformed for presentation, Days (D) after salt application a | |
Chlorophyll and leaf chloride ion content: Total chlorophyll content was higher at control than at salt treated plants (Table 2). Chlorophyll content decreased at higher salinities. The differences between control plants and more saline conditions were significant at (p≤0.05). However, chlorophyll a content was higher in the leaf than Chl b. An increase in Chl a:b ratio occurred in plants receiving salt concentration from 15-45 mM NaCl (Fig. 1) while those receiving the higher salt concentration of 60 mM NaCl experienced a marked decrease. Total chloride content of leaves increased significantly (p≤0.05) with external salinity in the growth medium (Table 2). Tip burn symptoms due to chloride (Mickelbart and Arpaia 2002) were visible on mature leaves of avocado plants exposed to high levels of NaCl salinity (30, 45 and 60 mM).
Gas exchange: Salinity stimulated PN in the first few days of salt application for the plants receiving low salt concentrations (Table 3). Net photosynthetic rate of salinised plants was 63.6 to 93.3% of the control plants after 39 days. Salinity treatment had significant effect on PN (p≤0.05) after 39 days of salt application. At 60 mM NaCl, the plants were slightly more affected than at all other salinity levels. Generally, gs of salinised plants significantly (p≤0.05) decreased throughout the experimental period in contrast to control plants (Table 3). The gs values after 39 days of salt application ranged from about 65.6 to 78.5% of values from control plants. Transpiration rate (E) decreased in response to increasing salt concentration of the growth medium (Table 3). From 15 to 60 mM NaCl, the decreases in E were 94.1, 93.9, 95.1 and 87.9% of control plants respectively after 39 days. Significant (p≤0.05) differences in E between control and salinity treatments were evident in all the days of measurement except on day 16. Stomatal limitation of water loss may account for these observations. Salinity stress of the growth medium (Table 3) caused significant (p≤0.05) increase in Ci. The increases in Ci were 141.7, 187.8, 222.0 and 295.8% of control plants values, from 15 to 60 mM NaCl respectively after 39 days of salt application.
Water Use Efficiency (WUE): Generally water use efficiency of salt treated plants was lower than that of control plants, but the differences among treatments were not statistically significant (p≥0.05) between day 7 and day 12 (Fig. 2). Probably this is a consequence of minimizing water loss at the expense of carbon acquisition, which may be an adaptive mechanism to water stress due to salinity stress.
 | |
| Fig. 1: | The Effect of saline water irrigation on Chal a: Chl b ratio of avocado seedling after 39 days. Each value is the mean of four replications ±SE |
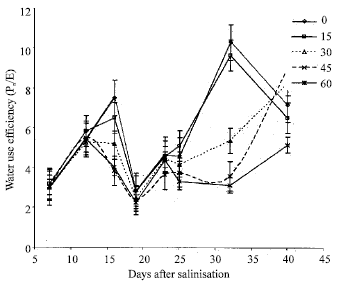 | |
| Fig. 2: | Effects of saline water irrigation on water use efficiency of avocado seedling. Each point represents the mean of four replications ±SE |
DISCUSSION
Salinity had a significant influence on the growth pattern of the avocado seedlings. The pattern of growth performance in height and dry weight indicate that growth parameters were decreased by saline irrigation. Salinity reduced shoot and root growth (Table 1a and b) of especially plants receiving water of highest salinity. A reduction in growth caused by increasing salinity is a well known phenomenon, but the growth of some plants may be stimulated by sodium chloride (Robinson et al., 1983; Soussi et al., 1998). An increase in shoot height and stem diameter growth observed from the study, but not detectable at 60 mM NaCl, may suggest increased cell growth and increased cell number due to osmotic adjustment. In most cases, salinity stress reduces root growth (Munns and Termaat, 1986; Reinhardt and Rost, 1995), although mild stress can increase extension as a result of osmotic adjustment process which maintains root growth during periods of salt stress. There was a short term stimulation of PN and growth during the first days of saline water irrigation (Table 1a and 3), except for 60 mM treatment. This stimulation was also evident in the results for stem diameter growth. According to Soussi et al. (1998), this observation may be attributed to increased activity of phosphoenolpyruvate carboxylase (PEPC). The stimulation may also be linked to improved water use efficiency through reduced water loss (Fig. 2). Increased leaf death and defoliation evidenced during the study may account for the few number of leaves (Table 1a) and hence reduced PN. Qalinity toxicity showed u` as interveinal leaf burn, scorch and dead tissues along the outside edges of leaves. The decline in net photosynthesis with increasing salifity was associated with similar reductions in gs in salt treated plants; so that there were only small changes in Ci of control plants than of salt treated plants (Table 3). Closure of the stomata could reduce Ci and CO2 assimilation rate (Boyer, 1971; Seemann and Sharkey, 1986; Ashraf et al., 2002; Netondo et al., 2004b). It is possible that the fluctuations in gs observed from time to time were due to increased vapor pressure deficit, which is known to increase with increased temperature. In the current study closure of the stoeata had only minimal contribution to reduction in internal CO2 concentration of salt treated plants; suggesting a presence of non stomatal factor being involved in reduction in PN (Hand et al., 1982; Bradford, 1983a; Sharp and Boyer, 1986; Rao et al., 1987; Belkhodja et al., 1999). There were fluctuations in PN almost everi week, which may be associated with the growth behaviour of avocado plants, since there are periods when the plants tend to have high percentage of young leaves.
The results indicate that chloride may play an important role in inhibition of chloroplasts reactions by inhibiting the synthesis of rubisco and chlorophyll or accelerate chlorophyll degradation (Soussi et al., 1998; Ashraf et al., 2002). The results indicated that chl a was higher than chl b showing that salinity induced a marked decrease in chl b. Reduction of chl b may suggest structural damage of the photosystem II reaction centres; and would explain the high Ci in salt treated avocado plants (Table 3). Earlier findings by Lutts et al. (1996) have indicated that chl b is associated with PS antenna. Losses in chloroplast activity include decreases in electron transport and photo-phosphorylation and are associated with changes in conformation of the thylakoids and of coupling factor (ATP synthetase, a sub unit of the thylakoids) and decreased substrate binding by coupling factor (Bradford, 1983b; Rao et al., 1987). Non-stomatal factors may be the limiting factors in decreases of photosynthetic activity under salinity stress (Richardson and McCree, 1985; Robinson, 1985; Bar et al., 1996; Soussi et al., 1998; Ashraf et al., 2002) and mai include inhibition of electron transport (Robertson et al., 1985; Soussi et al., 1998; Sibole et al., 2003; Netondo et al., 2004b). High external salt concentrations could affect thylakoid membranes by disrupting lipid bilayer or lipid protein associations and impair electron transport activity (Netondo et al., 2004b). According to Farquhar and Sharkey (1982), greater inhibition of net photosynthesis at high Ci than low, would suggest this stress affeats ribulose bisphosphate (RUBP) regeneration. Reduction in chlorophyll may partly account for the reduction in photosynthetic rate of avocado seedlings.
Water use efficiency is an important aspect in tolerance of salinity stress (Flowers et al., 1988; Gorrham et al., 1985). Initially; plants exhibited reductions in WUE with increasing NaCl levels and later own plants at higher salinity levels increased their WUE (Fig. 2). Generally, the exposure of plants to salinity stress resulted in a significant (p≤0.05) decrease in WUE. The observed results may be due to reduced water loss in salt treated plants than in control; caused by rapid decrease in water potential in the growth medium (Hand et al., 1982; Richardson and McCree, 1985; Munns and Termaat 1986, Munns, 2002). Higher salinity resulted in lower transpiration rates (Table 3), indicating that salinity caused a reduction in water loss per unit leaf area. This effect of salinity on transpiration has been reported in other plant species (Gorrham et al., 1985; Marler and Zozer, 1996; Ashraf et al., 2002). Other researchers have observed limited carbon supply due to increased incidence of necrotic margins on the leaves of salinity stressed plants (Oster and Arpaia, 1992; Cramer et al., 1994; Mickelbart and Arpaia, 2002), which would reduce the transpiration rate because of the reduced leaf area. An increase in WUE means that there was reduced transpiration rate than net photosynthesis per single leaf.
CONCLUSIONS
This study presents evidence showing that high growth inhibition of avocado seedlings at high salinity may be related to high leaf chloride content. The results show not only that salinity reduces growth and photosynthetic capacity of avocado plants, but also the combined effects of salinity and low PAR irradiance may contribute to reduced photosynthetic rate. Further studies are needed to determine the parameters related to chlorophyll fluorescence and gaseous exchange of the individual leaves to bring complementary information on the nature of constraints acting on photosynthetic processes. The study has shown clearly that this Kenyan avocado rootstock (var. Puebla) is sensitive to substrate salinity and hence cannot be depended upon in reclaiming saline problematic soils.
ACKNOWLEDGMENTS
We thank the institute of research and postgraduate studies of Maseno University- Kenya for providing financial support to conduct this research.
REFERENCES
- Arnon, D.I., 1949. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol., 24: 1-15.
CrossRefPubMedDirect Link - Ashraf, M., F. Karim and E. Rasul, 2002. Interactive effects of gibberellic acid (GA3) and salt stress on growth, ion accumulation and photosynthetic capacity of two-spring wheat (Triticum aestivum L.) cultivars differing in Salt tolerance. Plant Growth Reg., 36: 49-59.
CrossRef - Bar, Y., A. Apelbaum, U. Kafkafi and R. Goren, 1996. Polyamines in chloride-stressed Citrus plants: Alleviation of stress by nitrate supplementation via irrigation water. J. Am. Soc. Hortic. Sci., 121: 507-513.
Direct Link - Belkhodja, R., F. Morales, A. Abadia, H. Medrano and J. Abadia, 1999. Effects of salinity on chlorophyll fluorescence and photosynthesis of barley (Hordeum vulgare L.) grown under a triple-line-source sprinkler system in the field. Photosynthetica, 34: 375-387.
CrossRef - Cramer, G.R., G.J. Alberico and C. Schmidt, 1994. Leaf expansion limits dry matter accumulation of salt-stressed maize. Aust. J. Plant Physiol., 21: 663-674.
Direct Link - Farquhar, G.D. and T.D. Sharkey, 1982. Stomatal conductance and photosynthesis. Ann. Rev. Plant Physiol., 33: 317-345.
CrossRefDirect Link - Hasegawa, P.M., R.A. Bressan, J.K. Zhu and H.J. Bohnert, 2000. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol., 51: 463-499.
CrossRefDirect Link - Jimenez, M.S., A.M. Gonzalez-Rodriguez, D. Morales, M.C. Cid, A.R. Socorro and M. Caballero, 1997. Evaluation of chlorophyll fluorescence as a tool for salt stress detection in roses. Photosynthetica, 33: 291-301.
CrossRef - Lutts, S., J.M. Kinet and J. Bouharmont, 1996. NaCl-induced senescence in leaves of rice (Oryza sativa L.) cultivars differing in salinity resistance. Ann. Bot. 78: 389-398.
CrossRefDirect Link - Marler, T.E. and Y. Zozer, 1996. Salinity influences photosynthetic characteristics, water relations and foliar mineral composition of Annona squamosa L. J. Am. Soc. Hort. Sci., 121: 243-248.
Direct Link - Mickelbart, M.V. and M.L. Arpaia, 2002. Rootstock influences changes in ion concentration, growth and photosynthesis of Hass avocado trees in response to salinity. J. Am. Soc. Hortic. Sci., 127: 649-655.
Direct Link - Munns, R. and A. Termaat, 1986. Whole-plant responses to salinity Aust. J. Plant Physiol., 13: 143-160.
CrossRefDirect Link - Munns, R., 2002. Comparative physiology of salt and water stress. Plant Cell Environ., 25: 239-250.
CrossRefDirect Link - Netondo, G.W., J.C. Onyango and E. Beck, 2004. Sorghum and salinity: I. Response of growth, water relations and ion accumulation to NACl salinity. Crop Sci., 44: 797-805.
Direct Link - Netondo, G.W., J.C. Onyango and E. Beck, 2004. Sorghum and salinity: II. Gas exchange and chlorophyll fluorescence of sorghum under salt stress. Crop Sci., 44: 806-811.
Direct Link - Robertson, J.M., R.P. Pharis, Y.Y. Huang, D.M. Reid and E.C. Yeung, 1985. Drought induced increases in abscissic acid levels in the root apex of sunflower. Plant Physiol., 79: 1086-1089.
Direct Link - Seeman, J.R. and T.D. Sharkey, 1986. Salinity and nitrogen effects on photosynthesis, ribulose-1,5-bisphosphate carboxylase and metabolite pool size in Phaseolus vulgaris L. Plant Physiol., 82: 555-560.
Direct Link - Shalhevet, J., 1999. Salinity and water management in avocado. Proceedings of the Avocado Brainstorming, October 27-28, 1999, Riverside, CA. Hofshi Foundation, pp: 84-91.
Direct Link - Sibole, I.V., C. Cabot, C. Poschenrieder and J. Barcelo, 2003. Efficient leaf ion partitioning: An overriding condition for abscisic acid-controlled stomatal and leaf growth responses to NACl salinization in two legumes. J. Exp. Bot., 54: 2111-2119.
Direct Link - Soussi, M., A. Ocana and C. Lluch, 1998. Effects of salt stress on growth, photosynthesis and nitrogen fixation in chick-pea (Cicer arietinum L.). J. Exp. Bot., 49: 1329-1337.
Direct Link