
Mehrdad Lahouti
Department of Biology, Faculty of Science, Ferdowsi University, Mashad, Iran
Homa Mahmoodzadeh
Department of Biology, Faculty of Science, Islamic Azad University, Mashad, Iran
Samaneh Jamshidi
Department of Biology, Faculty of Science, Ferdowsi University, Mashad, Iran
International Journal of Botany
Year: 2007 | Volume: 3 | Issue: 4 | Page No.: 379-384







ABSTRACT
In the present research, effect of caffeine on structure and ultrastructure of shoot apical meristem of Phaseolus vulgaris L. was investigated. The present study showed that low concentrations of caffeine (<22.5 ppm) had no pronounced effect on the growth of bean seedlings and structure of shoot apical meristem, but higher concentrations of caffeine (40-60 ppm) had severe inhibitory effects. Electron microscopy (TEM and SEM) studies showed that apical meristem size and number of leaf primordia was decreased under caffeine treatment. Also in some meristem cells, formation of cell wall was incomplete. Present findings show that a lack of division in strategically located cells in primordia and during organogenesis could produce the organ abnormalities described. The observation that cytokinesis defects can be observed in developing primordial and meristems supports this.
PDF Abstract XML References Citation
How to cite this article
Mehrdad Lahouti, Homa Mahmoodzadeh and Samaneh Jamshidi, 2007. Effect of Caffeine on Structure and Ultrastructure of Shoot Apical Meristem of Phaseolus vulgaris L. International Journal of Botany, 3: 379-384.
DOI: 10.3923/ijb.2007.379.384
URL: https://scialert.net/abstract/?doi=ijb.2007.379.384
DOI: 10.3923/ijb.2007.379.384
URL: https://scialert.net/abstract/?doi=ijb.2007.379.384
INTRODUCTION
Plant morphogenesis is controlled by the rate and orientation of cell division and cell expansion. Cytokinesis is mediated by the assembly of the cell plate, an organelle whose position is governed by developmental cues. Morphological studies have elucidated the dynamic process of cell plate formation. The phragmoplast, a specialized cytoskeletal structure composed of microtubules and microfilaments, forms at the end of anaphase to guide cell plate assembly and expansion. During cytokinesis secretory vesicles carrying membrane and cell wall components are guided along the phragmoplast toward its center, where they fuse to form a membranous Tubulo-Vesicular Network (TVN) (Samuels and Steahlin, 1996). As additional vesicles are added, the TVN gradually develops into a smoother more plate-like structure that expands outward toward the margin of the cell, until it ultimately fuses with the parental plasma membrane yielding two individual cells separated by a new cell wall. The development of the phragmoplast is thought to be caused by the depolimerization of microtubules at the central region or the inner margin of the phragmoplast and the polymerization of microtubules at the outer margin (Gunning, 1982). In fact, the occurrence of rapid microtubule turnover in the phragmoplast has been demonstrated using florescence recovery after a photobleaching technique in cells microinjected with fluorescently labeled tubulin (Hush and Wadsworth, 1994). The arrested redistribution of phragmoplast microtubules similar to that observed in brefeldin A (BFA)-treated BY-2 cells, has been shown in Tradescantia stamen hair cells treated with caffeine, a well known inhibitor of plant cytokinesis (Valets and Hepler, 1997). Samuels and Staehelin (1996) reported that treatment of BY-2 cells with caffeine applied at metaphase inhibited the deposition of callose in the cell plate and the conversion of the fragile, thin, Fusion Tube-generated Membrant Network (FTN) into the more stable Tubulo-Vesicular Network (TVN) which normally occurs in forming cell plates. However, they did not mention the effects of caffeine on the redistribution of phragmoplast microtubules.
DNA and chromosomal damage effect of caffeine has also been reported by Paulovich (1997), Blasina and Brendan (1999), Sarkaria and Busby (1999) and Melo (2002) who observed chromosomal breakage as chromatin pieces in plant cells treated with caffeine.
A comparative study of the effect of 2, 6-dichlorobenzonitril and Caffeine on cytokinesis, was made by Buron and Hevdugo (1983). Dichlobenil had temporary inhibitory effect on cytokinesis metabolic pathway while caffeine had permanent effect. In another investigation caffeine caused rapid enhancement of cytosolic Ca2+ and alchohol dehydrogenase activity in cultured cells of Zea mays (Baxter and Ghang, 2003).
Several genes involved in plant cyotkinesis have been revealed recently through the characterization of mutant plants that either display defects in the determination of the division plane or that show aberrant cell plate formation. Mutants that fall into the first class such as ton/fass show defects in the proper alignment of the division machinery and thus have misoriented cell walls (Smith, 2001). In contrast, mutants that are defective in the construction of the cell plate have multinucleated cells with incomplete or missing cell walls (Nacry et al., 2000; Sollner and Glasser, 2002). The identification of these genes has yielded insight into molecular mechanisms that govern plant cytokinesis. However, given the complexity of this process, it is clear that genetic screens for other componenets of the cytokinetic machinery are not yet saturated.
The aim of the present research was to investigate effect of different concentrations of caffeine on the growth and biomass of Phaseolus vulgaris L. as an important crop plant and also to study ultrastructural features of apical meristem cells treated with caffeine using plant tissue culture and electron microscopy (TEM and SEM) techniques.
MATERIALS AND METHODS
The experiment was conducted in Science Faculty, Ferdowsi University, Mashad, Iran (2005). Seeds of Phaseolus vulgaris were germinated in seed germinator at 20°C. Nine day old seedlings were transferred to Hoagland nutrient solution containing 5, 10, 15, 17.5, 20, 27.5, 30, 40, 50 and 60 ppm caffeine. Seedlings were grown in growth chamber in hydroponic condition at 25°C (during 16 h light period) and 17°C (during 8 h darkness). Hydroponic solutions were frequently aerated and changed every week. Plants were harvested after 5 weeks and biomass and length of roots and shoots were measured and compared with the controls. Shoot apical meristem tissue explants were transferred onto solid MS culture medium containing Caffeine. Callus formation occurred after 15 days. Variations of callus formation in relation to different levels of added caffeine was studied.
Electron microscopy studies was carried out using TEM and SEM (LEO 914 AB, LEO1450 vp, respectively).
Apical meristem tissues from the original bean plants as well as callusmasses were passed through TEM and SEM preparation procedures. SEM tissue preparation procedure including tissue dehydration using different concentrations of acetone (25, 50, 70, 95 and 100%), coating by thin layer of Au-Pd was carried out. Tissue samples were placed on stubs and observed by SEM.
For TEM, apical meristem pieces were fixed in 3% glutaraldehyde in 75 mM potassium/sodium phosphate buffer, post-fixed in 2% osmium tetroxide, dehydrated in an acetone series and infiltrated with Spurr's epoxy resin. Sections were stained with uranyl acetate and lead acetate and examined with a LEO 914 AB TEM.
RESULTS AND DISCUSSION
Results of our experiments showed that increasing concentrations of caffeine (more than 20 ppm) significantly reduced vegetative growth of bean seedlings. Other growth factors including length of roots and shoots, lateral roots extension, number of leaves and leaf area index also significantly decreased in comparison with the control (Fig. 1). Toxic caffeine symptoms (marginal chlorosis and nectrotic spots of leaves) were more obvious at higher concentrations (30, 40, 50 and 60 ppm).
Greatest reduction in root and shoot length of the seedlings occurred at 60 ppm caffeine treatment. The same was true for both dry and fresh biomass of roots and shoots (Table 1).
Results of tissue culture experiment showed that fresh and dry weight of callus tissues obtained from shoot apical meristem cultured on MS culture medium decreased with increasing concentrations of caffeine. The greatest reduction occurred at 60 ppm treatment, 65% for fresh weight and 78% for dry weight compared to the controls (Table 1).
 | |
| Fig. 1: | Root and shoot extension of bean seedlings (caffeine-treated plant and control) |
| Table 1: | Effect of different concentration of caffeine on seedling growth and callus weight of Phaseolus vulgaris (values are means±SE) |
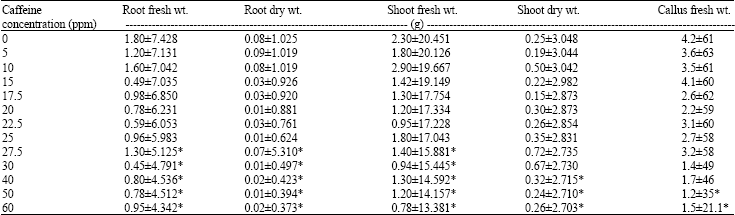 | |
| *: Based on Duncan's test, values are differ significantly p<0.05 | |
 | |
| Fig. 2: | Scanning electron microscopy images of shoot apical meristem of seedlings. (a) Control plant and (b) Treated with 60 ppm caffeine showing reduction of meristem size and number of leaf primordia. SAM: Shoot Apical Meristem; LF: Leaf Primordium |
SEM studies on shoot apical meristem showed that apical meristem size and number of leaf primordia decreased in caffeine-treated plants treatments, especially at 60 ppm caffeine (Fig. 2a and b).
TEM studies of shoot apical meristem of caffeine-treated plants revealed that cell wall formation of some meristematic cells was incomplete and only some parts of the wall was formed. Also vacuolar system of the treated meristematic cells was more extended compared to the controls (Fig. 3-5).
This study, showed that low concentrations of caffeine (<22.5 ppm) had no pronounced effect on the growth of bean seedlings and structure of shoot apical meristem, but higher concentrations of caffeine (40-60 ppm) had severe inhibitory effects. This is consistent with the previous reports that caffeine results in reducing of cell division and cell number of shoot apical meristem (Liu et al., 1995; Hepler and Bonsignore, 1990; Galatis and Apostolakos, 1991; Terryn et al., 1993). Caffeine, which also caused wall stubs, may guide vesicle fusion to the cell plate or block plate stabilization (Hepler and Bonsignore, 1990; Liu et al., 1995; Valets and Hepler, 1997).
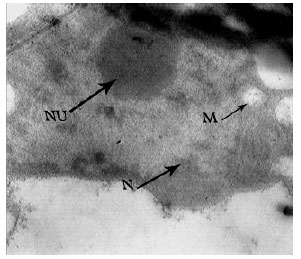 | |
| Fig. 3: | Transmission electron microscopy images of bean shoot apical meristem (untreated plant) showing nucleus, bulky nucleolus and mitochondria. Cytoplasm background is densed and osmiophyll due to abundance of ribosomes (x 3150): N: Nucleus; NU: Nucleolus; M: Mitochondrium |
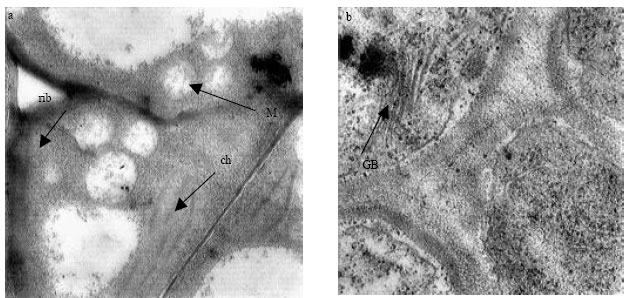 | |
| Fig. 4: | (a and b) Transmission electron microscopy images of bean shoot apical meristem chloroplasts (x3150). (untreated plant) showing a large number of mitochondria in cytoplasm, abundance of ribosomes, golgi body and differentiation some of proplastids to chloroplasts (x3150). M: Mitochondrium; ch: Chloroplast; rib: Ribosome; GB: Golgi body |
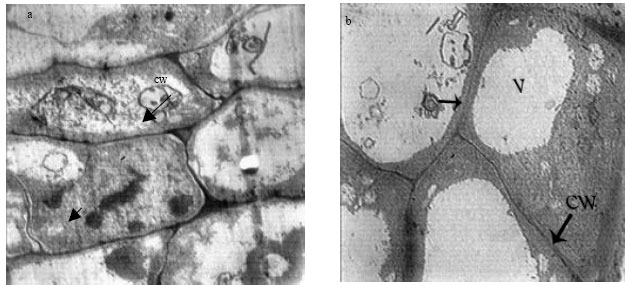 | |
| Fig. 5: | (a and b) Transmission electron microscopy images of bean shoot apical meristem (treated plant) showing incomplete cell wall (small arrow) (x 1600) V: Vacuole; CW: Cell Wall |
Caffeine acts at an early stage of ytokinesis, just after the Golgi-derived vesicles have arrived at the cell equator and begun to use with each other via thin (20 nm) membrane tubules. This initial round of fusions produces a delicate membrane network which in control cells is rapidly converted in a more substantial tubulo-vesicular network covered by a thick, fuzzy coat on its cytoplasmic surface. Caffeine disrupts the conversion of the fragile, thin, fusion tube-generated membrane network into the more stable tubulo-vesicular network, the assembly of its fuzzy coat and the budding of clathrin-coated vesicles from its surface (Samuels and Staehelin, 1996). Current theories of the mechanism of action of caffeine are about its ability to disrupt local calcium gradients.
Bonsignore and Hepler (1985) showed that caffeine easily reaches the target site and disrupts normal plate formation in Tradescantia stamen hair cells. In their experiments, treatment with caffeine that started 10 min before the onset of cell plate formation inhibited cytokinesis in all the cells examined. Caffeine inhibits callose deposition in the cell plate and the depolymerization of microtubules in the central region of the phragmoplast (Yasuhara, 2005). The following three possible mechanisms can be postulated to account for the concomitant inhibition of callose deposition and the phragmoplast microtubule depolymerization. First, caffeine inhibits callose deposition that might trigger the depolymerization of phragmoplast microtubules. Secondly, caffeine inhibits the depolymerization of phragmoplast that might trigger the callose deposition. Thirdly, caffeine might inhibit some signaling steps that trigger both the callose deposition and the microtubule depolymerization. The second possibility seems to be untenable because callose has already been deposited at the equatorial region of the phragmoplast at early developmental stages where the depolymerization of microtubules has not yet occurred (Samuels and Staehelin, 1996; Yasuhara and Shibaoka, 2000). Yang et al. (1999) showed the cyd1 mutant of Arabidopsis identifies a new locus affecting cytokinesis. The presence of wall protrusions, rather than central island, in both cytokinesis mutants and in caffeine-treated plants could imply that cell plates are more stable where they join the parent wall than in the cell center. Evidence suggests that wall maturation and insertion factors originate from the parent wall and spread into the new wall to stabilize it (Mineyuki and Gunning, 1990).
Present findings show that a lack of division in strategically located cells in primordial and during organogenesis could produce the organ abnormalities described. The observation that cytokinesis defects can be observed in developing primordial and meristems supports this hypothesis. Defects during emberyonic development may be of more consequence and more deleterious than those that occur after germination.
REFERENCES
- Baxter, A. and R. Ghang, 2003. Gene and enhance trap transposable elements reveal oxygen deprivation-regulated genes and their complex patterns of expression in Arabidopsis. Ann. Bot., 91: 129-141.
Direct Link - Blasina, A. and D.P. Brendan, 1999. Caffeine inhibits the checkpoint Kinase ATM. Curr. Biol., 9: 1135-1138.
Direct Link - Bonsignore, C.L. and P.K. Hepler, 1985. Caffeine inhibition of cytokinesis: Dynamics of cell plate formation-deformation in vivo. Protoplasma, 129: 28-35.
CrossRefDirect Link - Galatis, B. and P. Apostolakos, 1991. Microtubule organization and morphogenesis of stomata in caffeine-affected seedlings of Zea mays. Protoplasma, 165: 11-26.
Direct Link - Hepler, P.K. and C.L. Bonsignore, 1990. Caffeine inhibition of cytokinesis: Ultrastructure of cell plant formation/degradation. Protoplasma, 157: 182-192.
Direct Link - Hush, J.M. and P. Wadsworth, 1994. Quantification of microtubule dynamics in living plant cells using fluorescence redistribution after photobleaching. J. Cell Sci., 107: 775-784.
Direct Link - Liu, C.M., S. Johnson and T.L. Wang, 1995. Cyd, a mutant of pea that alters embryo morphology is defective in Cytokinesis. Dev. Genet., 16: 321-331.
Direct Link - Melo, J. and D. Toczyski, 2002. Unified view of the DNA-damage checkpoint. Curr. Opin. Cell Biol., 14: 237-245.
Direct Link - Mineyuki, Y. and B.E.S. Gunning, 1990. A role for preprophase bands of microtubules in maturation of new cell walls and a general proposal on the function of preprophase band sites in cell division in higher plants. J. Cell Sci., 97: 527-537.
Direct Link - Nacry, P., U. Mayer and G. Jurgens, 2000. Genetic dissection of cytokinesis. Plant Mol. Biol., 43: 719-733.
Direct Link - Paulovich, A.G., J.F. Lopez-Saez, P. Carrera, M.H. Navarrete and de la Torre, 2003. Effect of caffeine on in vivo processing of alkylated bases in proliferating plant cells. Cell Biol. Int., 27: 837-843.
CrossRef - Samuels, L. and L.A. Staehelin, 1996. Caffeine inhibits cell plate formation by disrupting membrane reorganization just after the vesicle fusion step. Protoplasma, 195: 144-155.
Direct Link - Sarkaria, J.N. and E.C. Busby, 1999. Inhibition of ATM and ATR kinase activities by the radiosensitizing agent, caffeine. Cancer Res., 59: 4375-4382.
Direct Link - Smith, L.G., 2001. Plant cell division: Building walls in the right places. Nat. Rev. Mol. Cell Biol., 2: 33-39.
Direct Link - Sollner, R. and G. Glasser, 2002. Cytokinesis-defective mutants of Arabidopsis. Plant Physiol., 129: 678-690.
Direct Link - Terryn, N., M.B. Arias and G. Engler, 1993. rha 1, a gene encoding a small GTP binding protein from Arabidopsis, is expressed primarily in developing guard cells. Plant Cell, 5: 1761-1769.
Direct Link - Valets, A.H.D. and P.K. Hepler, 1997. Caffeine inhibition of cytokinesis: Effect on the phragmoplast cytoskeleton in living tradescantia stamen hair cell. Protoplasma, 196: 155-166.
Direct Link - Yang, M., J. Nadeu, A. Zhao and F.D. Sack, 1999. Characterization of a cytokinesis defective (cyd1) mutant of Arabidopsis. J. Exp. Bot., 388:: 1437-1446.
Direct Link - Yasuhara, H. and H. Shibaoka, 2000. Inhibition of cell-plate formation by brefeldin the depolymerization of microtubules in the central region of the phragmoplast. Plant Cell Physiol., 41: 300-310.
Direct Link - Yasuhara, H., 2005. Caffeine inhibits callose deposition in the cell plate and the depolymerization of microtubules in the central region of the phragmoplast. Plant Cell Physiol., 46: 1083-1092.
Direct Link