
A. A. Arafa
Department of Agricultural Botany, Faculty of Agriculture,
Mansoura University, Mansoura, Egypt
M. A. Khafagy
Department of Agricultural Botany, Faculty of Agriculture,
Mansoura University, Mansoura, Egypt
M. F. El-Banna
Department of Agricultural Botany, Faculty of Agriculture,
Mansoura University, Mansoura, Egypt
International Journal of Botany
Year: 2007 | Volume: 3 | Issue: 3 | Page No.: 251-259







ABSTRACT
The effects of exogenously applied glycinebetaine or ascorbic acid (pre-soaking or pre-soaking plus spraying) on the salt-stress induced ultrastructural damages in sorghum seedlings were investigated. The seedlings grown in hydroponic culture containing nutrient solution for 3 weeks treated with NaCl at (0 and 6000 ppm). Salinity induced ultrastructural damages in leaf such as swelling of thylakoids, disintegration of grana stacking and intergrana lamellae, an increase in the number of plastoglobuli and starch grains (number and size), an increase in the number of mitochondria and destruction of mitochondria (few cristae, shortly the cristae long and some mitochondria appeared pale), shrinkage the plasma membrane, formation of the Myelin-Figures from plasma membrane. These damages were alleviated by the pretreatment (pre-soaking or pre-soaking plus spraying) with glycinebetaine at 4000 ppm or ascorbic acid at 100 ppm. It could be concluded that pre-soaking plus spraying with glycinebetaine at 4000 ppm or ascorbic acid at 100 ppm proved to be more effective since it reduced the harmful effect of salinity.
PDF Abstract XML References Citation
How to cite this article
A. A. Arafa, M. A. Khafagy and M. F. El-Banna, 2007. The Effect of Glycinebetaine or Ascorbic Acid on the Salt-Stress Induced Damages in Sorghum Plant Cells. International Journal of Botany, 3: 251-259.
DOI: 10.3923/ijb.2007.251.259
URL: https://scialert.net/abstract/?doi=ijb.2007.251.259
DOI: 10.3923/ijb.2007.251.259
URL: https://scialert.net/abstract/?doi=ijb.2007.251.259
INTRODUCTION
Salinity is known to affect many aspects of metabolism of plants and to induce changes in their anatomy. These changes are often considered to be adaptive, thus increasing the chances of survival during salinity stress; alternatively, they may be considered as signs of damage and disturbance of normal physiology (Poljakoff-Mayber, 1975). Salinity causes great losses to agriculture by lowering the yields of various crops. It caused various physiological and biological changes in plants where it destroyed cell membrane structure and permeability by causing lipid peroxidation and injuries to plant metabolism (Meneguzzo et al., 1998).
Salt stress caused swelling the thylakoids, swelling stroma, accumulation of a large starch grains and lipid droplets more than the control plants and increased number of starch grains in the chloroplasts as well as distortion of grana stacking (Yamane et al., 2003) and disintegration of grana stacking and intergrana lamellae in rice plant (Rahman et al., 2002). In addition, salinity caused disruption of mitochondrial structure and the whole mitochondria appeared to be swollen (Rahman et al., 2002). Furthermore, salinity caused detached plasma lemma from the cell wall at various positions, indicating plasmolysis and absent in control plants and increased the myelin-figure released from the plasma membrane (Rahman et al., 2001).
Glycinebetaine is one of the osmoregulators solutes naturally accumulate in the plants (Thomas et al., 1992). It is often localized in the cytoplasm of the leaf cells in response to salt stress (Rhodes and Hanson, 1993) and protects higher plants against salt not only by maintaining osmotic adjustment (Pollard and Wyn Jones, 1979) but also by stabilizing many functional units, like oxygen-evolving PSII complex and ATP synthesis (Harinasut et al., 1996) as well as cell membranes (Mansour, 1998). Apart from its major role, however, as an organic solute compatible with enzyme functioning (Rhodes and Hanson, 1993), glycinebetaine may have protective effects for membranes (Yang et al., 1996) during stress. More specifically, glycinebetaine can protect the O2 evolving machinery of chloroplasts when exposed to high NaCl concentrations (Murata et al., 1992).
Ascorbic acid is a major primary antioxidant (Nijs and Kelley, 1991), plays an important role in preserving the activity of enzymes (Padh, 1990). Moreover, Ascorbic acid is a natural antioxidant compound may be accumulated in all plants under normal and stress conditions.
Sorghum (Sorghum bicolor (L.) Moench. var. bicolor) is among the most important crop for the world human nutrition. Sorghum a moderately-tolerant species (Bernstein et al., 1995).
The present study is planned as an attempt to minimize the harmful effects of salinity on sorghum plants growing in NFT, through application of glycinebetaine or ascorbic acid either as pre-soaking or pre-soaking plus spraying application.
MATERIALS AND METHODS
Experiment was carried out in the glasshouse of the Agric. Bot. Dept., Fac. of Agric., Mansoura Univ. during the summer season 2005.
The following experiment was conducted to study the harmful effects of salinity on sorghum plants growing in NFT, through pre-soaking compared to pre-soaking plus spraying sorghum plants with osmoregulator (glycinebetaine) or antioxidant (ascorbic acid) under normal or saline conditions.
Plant materials: Sorghum (Sorghum bicolor (L.) Moench. var. bicolor) grains were secured from the Agricultural Research Centre, Giza, Egypt.
Chemicals:
| • | Glycinebetaine (GB) was supplied by Sigma Chemical Co., USA and used at the concentration of 2000 or 4000 ppm. |
| • | Ascorbic acid (AsA) was obtained from EL-Gomhoria Co., Egypt and was used at the concentration of 50 or 100 ppm. |
| • | Sodium Chloride (NaCl) from EL-Gomhoria Co., Egypt and was used at the concentration of 1500 (4.84 dS m-1), 3000 (7.19 dS m-1) and 6000 ppm (11.88 dS m-1). |
Methods of planting: A lot of homogenous sorghum grains were surface-sterilized by soaking in 0.001 HgCl2 for one min and washed with distilled water. Then divided into 5 groups and each group individually presoaked in distilled water (control), GB (2000 or 4000 ppm) or AsA (50 or 100 ppm). Grains were germinated between discs of filter paper (Whatman No. 1), placed in Petri dishes 11 cm moistened with 10 mL of Cooper nutrient solution (Cooper, 1979) salinized by NaCl at 1500 or 3000 or 6000 ppm in addition to non-salinized treatment. The grains were germinated in an incubator at 25±2°C in the dark.
The experiment layout consisted of 8 plastic channels (4 m long and 10 cm diameter). Every two channels were provided by an electric pump representing four groups (0, 1500, 3000 and 6000 ppm). Each channel had 40 pores (6 cm diameter). After 5 days 5 uniform sorghum seedlings with 3-5 seminal roots were transferred into 6 cm perforated pots (reticulated) containing peat moss and perlite (1:1) as a rooting medium. Every channel was divided into 5 sets i.e., water, GB1, GB2, AsA1, AsA2. Each set contained 8 seedlings (one seedling/pot) spaced 10 cm representing a Nutrient Film Technique (NFT).
Each group of channels was divided into 2 subgroups i.e., pre-soaking and pre-soaking plus spraying. The seedlings assigned for pre-soaking plus spraying were sprayed twice (7 and 14 days from transplanting) with the same levels previously applied in the first group (pre-soaking method).
To keep the concentrations of sodium chloride and mineral nutrients constant, the solution was changed every 5 to 7 days and the volume of the solution maintained by adding distilled water as required. A nutrient solution was pumped into the channels at a flow rate one L per minute from a reservoir containing 10 L.
For study the effect of salinity, glycinebetaine, ascorbic acid and their interactions on mesophyll ultrastructure, it was intended to illustrate only the treatment which proved to be more effective in this respect. Small pieces (5 mm2) from the right midrib region of the second leaf of sorghum were taken at the age 21 days after transferring plants into channels.
Double fixation in Glutaraladehyde (2.5%) and osmium tetroxide (1%) was used. The fixative solutions were prepared in 0.01 M phosphate buffer of 6.5. Glutaraladehyde was used first for overnight in the refrigerator and replaced with cold buffer for 15 min and then the buffer was replaced with osmium tetroxide for 1 h. Most of the osmium tetroxide was removed and replaced with two changes of buffer for 15 min each. The materials were passed along the dehydration gradient by substituting the buffer with 50, 70, 80, 95 and 100% acetone, 10-15 min in each except 100% for which two changes of 30 min were made. The pure acetone was replaced gradually with a mixture of acetone/epoxy nesin (2:1, 1:1 and 1:2) for 15 min each. The dilute resin was then replaced with pure resin for overnight and was replaced again with fresh resin for 2 h and the materials were thus ready for embedding. Thick sections were made first to select the suitable area for ultrathin section (50-100 μ) using LKB ultratone III microtome. Sections were collected on copper grids and double stained with saturated uranyl acetate in 70% ethanol and Reynolds lead citrate for 15 min each. Sections were viewed, investigated and photographed, using transmission electron microscopy (JEOL 100s TEM).
RESULTS AND DISCUSSION
Chloroplast ultrastructure: The ultrastructure of sorghum mesophyll chloroplast of plants growing under normal conditions shows normal appearance where lamellae were differentiated into grana and intergrana regions (Fig. 1a). In addition, the thylakoids were generally oriented parallel to the long axis of the chloroplast. Moreover, the chloroplast contained a few small starch grains appeared through the stroma (Fig. 1a).
The chloroplast ultrastructure of sorghum leaf grown under NaCl salinity showed swelling, but the internal stroma was not affected and the inner lamellae were clearly distinguished into grana and intergrana (Fig. 1c). Moreover, some chloroplasts contained a large number of plastoglobuli and an increase of starch grains (number and size) (Fig. 1b) as compared to control (Fig. 1a).
Pre-soaking sorghum grains in glycinebetaine at 4000 ppm or ascorbic acid at 100 ppm and grown under NaCl at 6000 ppm showed that the chloroplast appeared normal and a well-defined inner lamellar structure but, the chloroplast contained a few small of plastoglobuli (Fig. 1d and e). It could be concluded that ascorbic acid at 100 ppm proved to be more effective since it reduced the harmful effect of salinity on chloroplast ultrastructure.
Concerning, glycinebetaine at 4000 ppm or ascorbic acid at 100 ppm (pre-soaking plus spraying) combined with NaCl at 6000 ppm NaCl (Fig. 1f and g) shows maintenance the chloroplast ultrastructure as compared to treatment by salinity.
In addition, chloroplast in plants treated with GB 4000 ppm contained a few large starch grains as compared to plants grown under NaCl. In could be concluded that pre-soaking in GB at 4000 ppm or ascorbic acid at 100 ppm proved to be more effective since it reduced the harmful effect of salinity on chloroplast ultrastructure.
The structural changes and swelling of thylakoid caused by the treatment with NaCl in the present study are probably due to change in the ionic composition of the stroma liquid (Yamane et al., 2003). Thus, swelling of the thylakoids membranes is probably due to a change in the ionic composition of the stroma liquid, rather than an effect of sodium ion on stacking of membranes. The other interesting point in chloroplast structure under salinity conditions was that the chloroplast stroma contained large starch grains (Fig. 2b). Rahman et al. (2000) suggested that the increased accumulation of starch in the chloroplast under salinity may be attributed to either the damage of the sucrose-phosphate synthesis in the cytosol, leading the triose phosphate pathway towards starch synthesis, or, to the damage of enzymes involved in starch degradation via changes in the ionic composition in the chloroplast. Under normal conditions, plants effectively scavenge active oxygen species, but the balance between the production of active oxygen Species and antioxidant systems is upset by senescence and abiotic stresses such as drought, salt, low-and high temperature, air pollutants and heavy metals, which often result in oxidative damage (Langebartels et al., 2002).
Mitochondria ultrastructure: A typical mitochondria in sorghum mesophyll cells of the control leaf are shown in (Fig. 2a). The mitochondria processed typical well defined double membranes and normal distribution of cristae. While, NaCl at 6000 ppm induced clear changes in the mitochondria ultrastructure. The most notable changes were that the mitochondria had a double membrane with a few cristae, shortly the cristae long and some mitochondria appeared pale or clear, as well as increasing the number of mitochondria (Fig. 2b). Pre-soaking sorghum grains in glycinebetaine at 4000 ppm or ascorbic acid at 100 ppm combined with NaCl at 6000 ppm, had no effect on reducing the harmful effect of salinity on mitochondria ultrastructure (Fig. 2c and d). Moreover, the number of mitochondria was reduced compared to plants grown under salinity alone (Fig. 2b). While, glycinebetaine at 4000 ppm or ascorbic acid at 100 ppm (pre-soaking plus spraying) reduced the harmful effect of salinity on mitochondria ultrastructure and decreased its number. Ascorbic acid treatment increased the size of mitochondria to long axis compared to control plants or salinity treatment. The damaged elicited by NaCl treatment in mitochondria may result from at least one or more of the following responses (a) a large accumulation of sodium and/or chloride ions in leaf cells (Rahman et al., 2002); (b) a difference in ion compartmentation, the deleterious ion (s) having greater access to the mitochondria in plants; and/or (c) a greater sensitively of the mitochondria to the presence of the ions in the cells or the culture medium (Smith et al., 1982).
Furthermore, Koryo et al. (1993) reported that the salt induced activation of F-ATPase in sorghum corresponding to the increase in the number of mitochondria in the root tip cells and therefore, may supply additional energy for osmotic adaptation and for selective uptake and transport processes. The salt stress led to an increase in the number of mitochondria and swollen cristae and also F-ATPase activity.
Cell wall and plasma membrane ultrastructure: Salinity caused shrinkage plasma membrane and partly detachment (Fig. 3b) of the cell wall at various positions indicating membrane vesicles (Fig. 3d). In addition, cytoplasm contained small and large membrane vesicles from the plasmalemma and fragmentation or degradation of tonoplast (Fig. 3d), but the vacuolar volume or size was increased (Fig. 3b and c).
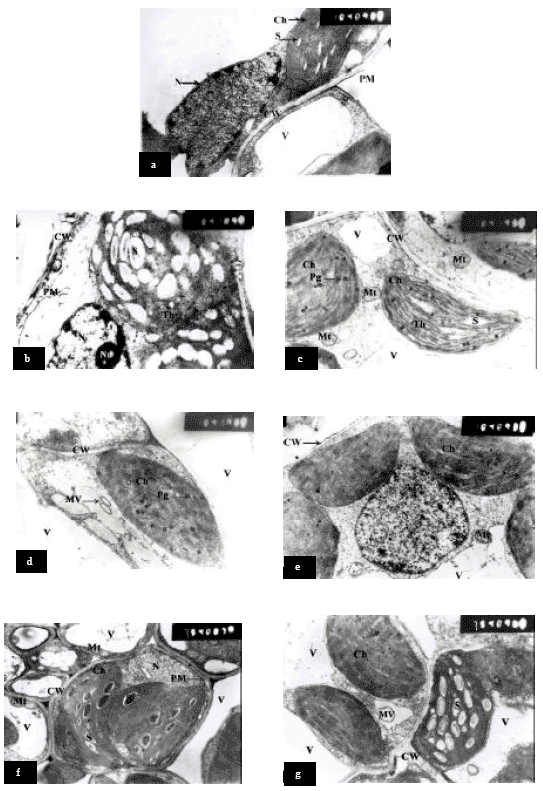 | |
| Fig. 1: | Transmission electron micrographs of mesophyll cells of Sorghum showing chloroplast ultrastructural changes a = Control, b; c = 6000 ppm NaCl, d = Pre-soaking in GB 4000 ppm + NaCl 6000 ppm, e = Pre-soaking in AsA 100 ppm + NaCl 6000 ppm, f = Pre-soaking plus spraying with GB 4000 ppm + NaCl 6000 ppm, g = Pre-soaking plus spraying with AsA 100 ppm + NaCl 6000 ppm, Ch = Chloroplast, CW = Cell Wall, MV = Membrane vesicles, Mt = Mitochondria, N = Nucleus, Nu = Nucleolus, Pd = Plasmodesmata, Pg = Plastoglobuli, PM = Plasma Membrane, S = Starch grain, V = Vacuole and Th = Thylakoid |
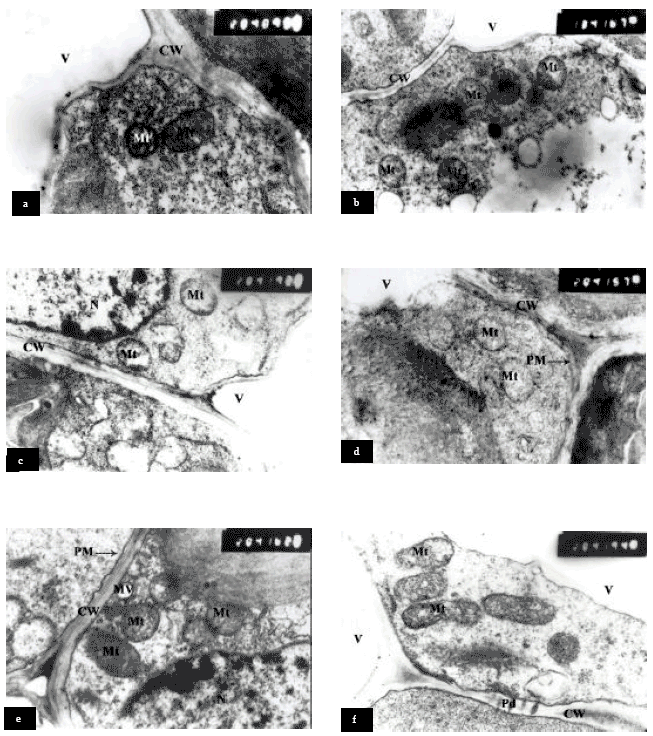 | |
| Fig. 2: | Transmission electron micrographs of mesophyll cells of Sorghum showing mitochondria ultrastructural changes. a = Control, b; c = 6000 ppm NaCl, d = Pre-soaking in GB 4000 ppm + NaCl 6000 ppm, e = Pre-soaking in AsA 100 ppm + NaCl 6000 ppm, f = Pre-soaking plus spraying with GB 4000 ppm + NaCl 6000 ppm, g = Pre-soaking plus spraying with AsA 100 ppm + NaCl 6000 ppm, CW = Cell Wall, MV = Membrane vesicles, Mt = Mitochondria, N = Nucleus, Nu = Nucleolus, Pd = Plasmodesmata, PM = Plasma Membrane and V = Vacuole |
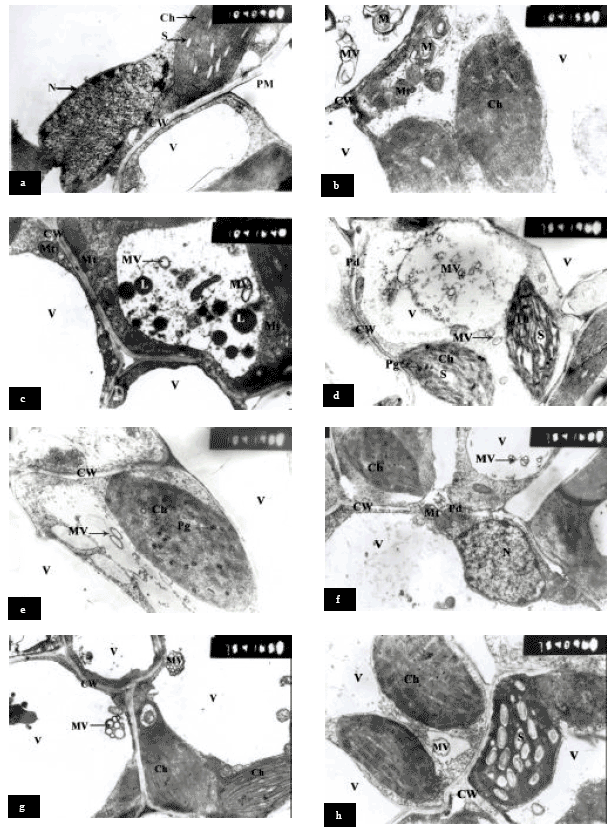 | |
| Fig. 3: | Transmission electron micrographs of mesophyll cells of Sorghum showing cell wall and plasma membrane ultrastructural changes. a = Control, b; c = 6000 ppm NaCl, d = Pre-soaking in GB 4000 ppm + NaCl 6000 ppm, e = Pre-soaking in AsA 100 ppm + NaCl 6000 ppm, f = Pre-soaking plus spraying with GB 4000 ppm + NaCl 6000 ppm , g = Pre-soaking plus spraying with AsA 100 ppm + NaCl 6000 ppm, Ch = Chloroplast, CW = Cell Wall, L = lipid droplets, M = Myelin-Figures, MV = Membrane vesicles, Mt = Mitochondria, N = Nucleus, Nu = Nucleolus , Pd = Plasmodesmata, Pg = Plastoglobuli, PM = Plasma Membrane, S = Starch grain, V = Vacuole and Th = Thylakoid |
Furthermore, the Myelin-Figures were also found from plasma membrane (Fig. 3b), which were absent in control plants (Fig. 3a). Moreover, a large amount of lipid droplets in cytoplasm was accumulated in sorghum leaf cells (Fig. 3c).
Pre-soaking sorghum grains in glycinebetaine at 4000 ppm or ascorbic acid at 100 ppm and grown under NaCl at 6000 ppm, maintained the plasma membrane and cell wall structure and no shrinkage or detachment of plasma membrane was detected. In addition, the membrane vesicles were less compared to treatment with NaCl. While, some chloroplast contained a few of plastoglobuli (Fig. 3e).
The application of glycinebetaine or ascorbic acid (pre-soaking plus spraying) on plants growing under NaCl at 6000 ppm reduced the harmful effects of salinity on plasmamembrane and cell wall ultrastructure, similar to the effect of pre-soaking (Fig. 3g and h). In addition, plants treated with pre-soaking plus spraying showed the membrane vesicles aggregated in side of the plasmamembrane (Fig. 3g). In this study, the most frequently observed ultrastructural alteration due to NaCl treatment was the formation of many small to large vacuoles and membrane vesicles in the leaf mesophyll cells. Therefore, Munns and Termaat (1986) pointed out that salt injury may be due to salt accumulation to the levels exceeding the ability of the cells to compartmentalize the salt to the vacuole.
The appearance of vesiculation in the cells after exposure to NaCl stress is an adaptive response to compartmentalize sodium ions away from the cytosol and these vesicles might act to store the toxic ions thus avoiding their injurious action in the cytoplasm (Cachorro et al., 1993).
Furthermore, Koryo (1997) pointed out that a multiplicity of small vacuoles possesses larger surface than one big vacuole, hence a higher exchange capacity (Na versus K).
This system enables a plant cell to avoid ion toxicity, imbalance and interactions between substances in the cytoplasm (Rahman et al., 2001).
Myelin-figures as observed in the present study are considered as artifactas due to double fixation with gluteraldehyde and osmium tetroxide (Bowers and Maser, 1988). These artifactas were absent in the control plants but they were observed in the NaCl-Treated plants and thus were considered as a reflection of the membrane changes due to the NaCl Treatment (Rahman et al., 2001). The lipid droplets which were not found in the control plants leaves observed in the cytoplasm of salt-treated plants may be due to loss of lipid in the cell membranes (Olmos and Hellin, 1996).
In this study, the plasma membrane was detached from the cell wall at several positions indicating plasmolysis which might be caused by excess ions or osmotic stress (Pareek et al., 1997). In addition, salt stress is known to result in extensive lipid peroxidation, which has often been used as indicator of salt induced oxidative damage in membranes (Hernández and Almansa, 2002). Ascorbic acid (AsA) is one of the most studied and powerful antioxidants that exist mostly in the reduced form (90%) of the ascorbate pool in leaves and chloroplasts (Smirnoff, 2000).
Ascorbic acid regenerates tocopherol from tocopheroxyl radical providing membrane protection and carries out a number of non-antioxidant functions in the cell. It has been implicated in the regulation of the cell division, cell cycle progression and cell elongation (De Tullio et al., 1999).
Glycinebetaine is a one such osmoprotectant and it occurs in many drought-and salt tolerant angiosperms. It is likely that betaine is involved in the protection of macro component of plant cell, such as protein complex and membranes under stress conditions (Sakamoto and Murata, 2000).
The action of betaine in vivo is not, however confined to Osmoregulation. Numerous studies in vitro have indicated that betaine acts as an Osmoprotectant by stabilizing both the quaternary structure of proteins (Gorham, 1995). Moreover, exogenous application of betaine to leaves has been shown to increase the tolerance to various stresses of several species of plants, including both natural accumulators and non-accumulators (Mäkelä et al., 1996).
In the present study exogenous application of glycinebetaine on sorghum plants reduced the harmful effect of salinity on ultrastructure of mesophyll cells compared to plant treated by NaCl salinity. Moreover, Rahman et al. (2002) showed that salinity induced ultrastructural damages in leaf and such damages were largely prevented by pretreatment with glycinebetaine may be due to protection of membranes from lipid peroxidation Chen et al. (2000) as well as protect the integrity and stability of membrane under salt stress (Fang et al., 2004).
From the abovementioned results it could be concluded that pre-soaking plus spraying application was more effective in alleviating the damages caused by NaCl salinity than the pre-soaking one. Moreover, pre-soaking plus spraying with glycinebetaine at 4000 ppm or ascorbic acid at 100 ppm proved to be more effective in this respect.
REFERENCES
- Bernstein, N., W.K. Silk and A. Lauchli, 1995. Growth and development of sorghum leaves under condition of NaCl stresses: Possible role of some mineral elements in growth inhibition. Planta, 196: 699-705.
Direct Link - Cachorro, P., A. Ortiz and A. Cerda, 1993. Effects of saline stress and calcium on lipid composition in bean roots. Phytochemistry, 32: 1131-1136.
Direct Link - Chen, W.P., P.H. Li and T.H. Chen, 2000. Glycinebetaine increases chilling tolerance and reduce chilling induced lipid peroxidation in Zea mays L. Plant Cell Environ., 23: 609-618.
Direct Link - De-Tullio, M.C., C. Paciolla, F. Dalla Vecchia, N. Rascio and S. D'Emerico et al., 1999. Changes in onion root development induced by the inhibition of peptidyl-prolyl hydroxylase and influence of the ascorbate system on cell division and elongation. Planta, 209: 424-434.
Direct Link - Fang, G.Q., M.Q. Quan, S. Can, L.S. Zhong, Z.Q. Meng and W. Wei, 2004. Effects of exogenous glycinebetaine on the salt-tolerance of wheat. Acta Botanica Boreali Occidentalia Sin., 24: 1680-1686.
Direct Link - Harinasut, P., K. Tsutsui, T. Takabe, M. Nomura, T. Takabe and S. Kishitani, 1996. Exogenous glycinebetaine accumulation and increased salt-tolerance in rice seedlings. Biosci. Biotech. Biochem., 60: 366-368.
Direct Link - Hernandez, J.A. and M.S. Almansa, 2002. Short-term effects of salt stress on antioxidant systems and leaf water relations of pea leaves. Physiol. Plant., 115: 251-257.
CrossRefDirect Link - Koryo, H.W., R. Stelzer and B. Huchzermeyeyer, 1993. ATPase activities and membranes fine structure of rhizodermal cells from Sorghum and Spartina roots grown under mild salt stress. Bot. Acta, 106: 110-119.
Direct Link - Koryo, H.W., 1997. Ultrastructural and physiological changes in root cells of sorghum plants (Sorghum bicolor x S. Sudanensis cv. sweet sioux) induced by NaCl. J. Exp. Bot., 48: 693-706.
Direct Link - Langebartels, C., H. Wohlgemuth, S. Kschieschan, S. Grun and H. Sandermann, 2002. Oxidative burst and cell death in ozone-exposed plants. Plant Physiol. Biochem., 40: 567-575.
Direct Link - Makela P., P. Peltonen-Sainio, K. Jokinen, E. Pehu and H. Setala et al., 1996. Uptake and translocation of foliar-applied glycinebetaine in crop plants. Plant Sci., 121: 221-230.
Direct Link - Mansour, M.M.F., 1998. Protection of plasma membrane of anion epidermal cells by glycinbetaine and praline against NaCl stress. Plant Phsiol. Biochem., 36: 767-772.
CrossRefDirect Link - Murata, N., P.S. Mohanty, H. Hayashi and G.C. Papageoriou, 1992. Glycinebetaine stabilizes the association of extrinsic proteins with the photosynthetic oxygen-evolving complex. FEBS Lett., 296: 187-189.
Direct Link - Nijs, D. and P.M. Kelley, 1991. Vitamin C and E. donate single hydrogen atoms in vivo. FEBS Lett., 284: 147-151.
Direct Link - Pareek, A., S.L. Singla and A. Grover, 1997. Short-term salinity and high temperature stress-associated ultrastructural alterations in young leaf cells of Oryza sativa L. Annl. Bot., 80: 629-639.
Direct Link - Pollard, A. and R.G.W. Jones, 1979. Enzyme activities in concentrated solutions of glycinebetaine and other solutes. Plants, 144: 291-298.
CrossRef - Rahman, M.S., T. Matsumuro, H. Miyake and Y. Takeoka, 2000. Salinity-induced ultrastructural alternations in leaf cells of rice (Oryza sativa L.). Plant Prod. Sci., 3: 422-429.
Direct Link - Rahman, M.S., T. Matsumuro, H. Miyake and Y. Takeoka, 2001. Effects of salinity stress on the seminal root tip ultrastructure of rice seedlings (Oryza sativa L.). Plant Prod. Sci., 4: 103-111.
Direct Link - Rahman, M.S., H. Miyake and Y. Takeoka, 2002. Effects of exogenous glycinebetaine on growth and ultrastructure of salt-stressed rice seedlings (Oryza sativa L.). Plant Prod. Sci., 5: 33-44.
Direct Link - Rhodes, D. and A.D. Hanson, 1993. Quaternary ammonium and tertiary sulfonium compounds in higher plants. Ann. Rev. Plant Physiol. Plant Mol. Biol., 44: 357-384.
CrossRefDirect Link - Smirnoff, N., 2000. Ascorbic acid: Metabolism and functions of a multi-facetted molecule. Curr. Opin. Plant Biol., 3: 229-235.
PubMedDirect Link - Thomas, C.E., L.R. McLean, R.A. Parker and D.F. Ohlweiler, 1992. Ascorbate and phenolic antioxidant interactions in prevention of liposomal oxidation. Lipids, 27: 543-550.
CrossRefDirect Link - Yamane, K., M. Kawasaki, M. Taniguchi and H. Miyake, 2003. Differential effect of NaCl and polyethylene glycol on the ultrastructure of chloroplasts in rice seedlings. J. Plant Physiol., 160: 573-575.
Direct Link - Yang, G., D. Rhodes and R.J. Joly, 1996. Effect of high temperature on membrane stability and chlorophyll fluorescence in glycinebetaine-deficient and glycinebetaine-containing maize lines. J. Plant Physiol., 23: 437-443.
Direct Link - Munns, R. and A. Termaat, 1986. Whole-plant responses to salinity Aust. J. Plant Physiol., 13: 143-160.
CrossRefDirect Link