
A. A. Adelusi
Department of Botany, Obafemi Awolowo University, Ile-lfe, Nigeria
O. A. Akamo
Department of Botany, Obafemi Awolowo University, Ile-lfe, Nigeria
A. M. Makinde
Department of Botany, Obafemi Awolowo University, Ile-lfe, Nigeria
International Journal of Botany
Year: 2006 | Volume: 2 | Issue: 1 | Page No.: 74-82







ABSTRACT
Macrotyloma geocarpa (Harms) Marechal and Baudet (Kersting`s groundbean), a grain legume was subjected to different nitrogen levels and spurge weed (Euphorbia heterophylla Linn) competition to study the combined effects of these factors on its growth. Weed competition was achieved by growing Kersting`s groundbean with spurge weed, while fertilizer stress was imposed by planting Kersting`s groundbean without fertilizer treatments. There was another treatment, which was a combination of the two stresses. Fertilizer application of the crops was achieved by broadcasting NPK fertilizers at 0, 15 and 30 kg N ha-1 in the Kersting`s groundbean seedlings plots. Plants grown under nitrogen fertilizer did better in terms of above ground biomass, root length, shoot height, Leaf Weight Ratio (LWR) and seed yield. These increased at successive intervals. Leaf Area (LA), Leaf Area Ratio (LAR), Specific Leaf Area (SLA), Leaf Area Duration (LAD), Relative Growth Rate (RGR), Leaf Area Index (LAI), Net Assimilation Rate (NAR) did not show a regular pattern at successive harvest times during the investigation.
PDF Abstract XML References Citation
How to cite this article
A. A. Adelusi, O. A. Akamo and A. M. Makinde, 2006. Nitrogen Fertilizer and Weed (Euphorbia heterophylla Linn.) Effects on the growth Parameters of Macrotyloma geocarpa (Harms) Marechal and Beudet. International Journal of Botany, 2: 74-82.
DOI: 10.3923/ijb.2006.74.82
URL: https://scialert.net/abstract/?doi=ijb.2006.74.82
DOI: 10.3923/ijb.2006.74.82
URL: https://scialert.net/abstract/?doi=ijb.2006.74.82
INTRODUCTION
There is a need to increase the performance of pulse crops, particularly in developing countries where most grain production is for human consumption and the demand is increasing due to population increase.
The agronomic problems associated with pulse differ between geographical areas. In Asia, Africa and oceanic regions (Johansen et al., 1992), North America (Muehlbauer and Kaiser, 1992) and Europe (Monti et al., 1992), drought and biotic stresses appear to be the major limiting factor while other stresses such as extreme temperature and nutrient deficiencies have less frequent impact. Crop management and plant breeding are among the various ways by which stresses could be alleviated.
Macrotyloma geocarpa is an indigenous grain legume cultivated in parts of Tropical Africa for food. It produces its seeds underground (Hepper, 1963). Its cultivation is not as widespread as cowpea and other legumes. The leaves are sometimes eaten in soup. Dried seeds of approximately 100 g can yield 348 calories and contains 9.7% moisture, 19.4 g proteins, 1.1 g fat, 66.6 g total carbohydrate, 5.5 g fibre, 3.2 g ash, 100 mg calcium etc.
The pods mature underground and are indehiscent usually divided by 1 or 2 constrictions into 2 or 3 joints seeds are oblong to oblong-ovoid, about 0.6 - 1.3 cm long, kidney-shaped with a white hilium, white, red, black, or mottled in colour. The seeds resemble the seeds of Phaseolus vulgaris but smaller and very hard when dried. Since the seeds are buried in the soil they are safe from attacks by flying insects that severely limit or destroy pulses like soybeans whose pods remain in the air. It takes between 4-5 months to mature. Seeds ripen, as leaves turn yellow. Plants are later dug up and left on the ground to dry and later beaten with sticks or in a mortar to remove seeds. These are later dusted with insecticide to prevent attack by weevils. The protein is rich in essential amino acids, such as lysine 6.2% and methionine 1.4%.
The roots grow well in fine sand or silt where phosphorus and nitrogen are available but roots do not move into regions of moist gravel or coarse sand easily even when fertilizer is applied. The extent of the root system is related to texture and structure of soils as well as available nutrients. Some grain legumes such as varieties of cowpea will do well with about 20 kg N ha-1 and 8 kg ha-1 (Ezedinma, 1961; Fennel, 1962). Optimum response to fertilizer as well as overall yield levels depends on timely sowing (Bandyopadhyay and De, 1986). Nitrogen deficiency is known to cause a reduction in the photosynthetic capacity of plants (Simpson et al., 1983). The quantum yield for CO2 uptake decreased heavily with leaf nitrogen content. The critical nitrogen concentration of a plant can be defined as the minimum nitrogen concentration required for maximum growth rate at any time (Sheehy et al., 1996). Leaching and erosion have been the cause of low levels of nutrients particularly nitrogen in cultivated soils.
Weeds are known to constitute a major limiting factor to grain legumes production and probably the most important yield depressing factor to grain legumes in Nigeria (Fadayomi, 1979). Weeds compete for nutrients, moisture, light and space. Weeds with good competitive ability show a faster rate of root elongation and development than the crop (Ayeni et al., 1984). Such weeds cause yield losses ranging from 50 to 86% (Moudy, 1973; Remison, 1978; Akobundu, 1979). Weed competition is most serious when crop is young and the critical time for weeding has been reported to be between 4 and 6 weeks after planting (Fadayomi, 1979; Ayeni et al., 1984). Weeds may also serve as a host for insects, pests and pathogens (Akinyemiju, 1987; Akinyemiju and Echendu, 1987).
In the present study nitrogen as a fertilizer and Euphorbia heterophylla that has been identified as a common weed in grain legume fields were applied to Macrotyloma geocarpa and the growth and yield responses followed over a period of time.
MATERIALS AND METHODS
The site of the experiment is situated in the Biological Gardens unit of Obafemi Awolowo University Ile-Ife, Nigeria. Soil samples were tested for pH and found to be 6.8, which is still ideal for the growth of legumes, which ranges between 6.5-7.0. Seeds of Macrotyloma geocarpa were collected from Plant Science Department, Faculty of Agriculture, Obafemi Awolowo University, Ile-Ife. The site was cleared and ridges were made. This was divided into three replicates R1, R2 and R3 and space between each replicates being 1 m. Each replicate was divided into six representing,
Weed +Nitrogen control (WN0)
Weed +Nitrogen dose 1 (WNI)
Weed +Nitrogen dose 2 (WN2)
Nitrogen Control (N0)
Nitrogen dose 1 (N1)
Nitrogen dose 2 (N2)
With a space of 60 cm between each one which was 3x2.4 m.
Analysis started on the day of fertilizer application that is, zero day and at 7 day intervals thereafter.
Seeds of Macrotyloma geocarpa were planted in a space of 60x60 cm. NPK fertilizer was applied 18 days after germination. Application was at 15 kg and 30 kg N ha-1.
Shoot height was determined by picking 6 plants randomly from the subunits measured by transparent rule-measure graduated in centimeter. Shoot height was measured from the soil level to the apex of the main stem.
Root length was determined from just below the soil level after uprooting the plant gently the attached soil removed by soaking in water. Root length was determined from below the soil to the apex of the taproot. This was done using a transparent rule measure.
Total leaf area was determined by measuring the length and width of the terminal leaflet and leaf area expressed as follows: L x W x 0.75.
Total values from all leaflets were later multiplied by 3 according to Nangju and Wanki (1980).
LA /plant = Total leaf area of terminal leaflets x 3
The weights of the root and shoot were also determined separately and these were dried to constant weight at 80oC. The weight of the whole plant was also determined.
The Leaf Area Index (LAI) was determined using the formula
LW/AGB x LA/LW = S/AGB
LW- Leaf weight
AGB- Above ground biomass
S- total leaf area (Olowe, 1989).
Leaf Area Ratio (LAR) accounts for total surface area used for assimilation per unit of plant biomass.
LAR = LA/ WS (West et al., 1920)
The unit is cm2 g-1 where WS is plant dry weight. Or LAR = S/W where, Sis total leaf area and W is plant dry weight.
Leaf Weight Ratio (LWR) was determined using the formula:
LWR=W1/WS where, W1 is leaf dry weight and WS is the plant dry weight. LWR defines assimilation in terms of leaf density and the unit is g g-1 (Evans, 1972).
Specific Leaf Area (SLA) was determined using the formula: S/LW (Beadle, 1985), where, S is the total leaf area and LW is leaf weight.
Leaf Area Duration (LAD) was determined using the formula: LA2/lnLA2-LA1/lnLA1 * dt (Evans, 1972). The above accounts for changes over a particular period in terms of persistent assimilatory surface over a time interval. Where,
LA1-First leaf area
LA2-Second leaf area.
Unit Leaf Rate (ULR)/ Net Assimilation Rate (NAR) was determined using the following formular:
1/S x dw/dt where, S is total leaf area (Beadle, 1985).
Relative growth rate was determined using the formula:
1/WS x dwS/dt
=1/La x dwS/dt x LA/WS
This measures the dry matter accumulated at a particular time regardless of the assimilatory surface present and the unit is g g-1d-1 (Evans, 1972).
RESULTS
The effect of nitrogen fertilizer and weed competition on dry matter production in Macrotyloma geocarpa showed that total plant dry weight were higher in N1 plants, followed by N2, WN2, N0, WN0 while WN1 plants recorded the lowest total plant dry weight (Table 1).
| Table 1: | The total plant dry weight at five havest period of Macrotyloma geocarpa treated with different levels of nitrogen fertilizer and weed (values are means with standard errors of means) |
 | |
| Table 2: | Coefficient of Variation (CV) showing means variation |
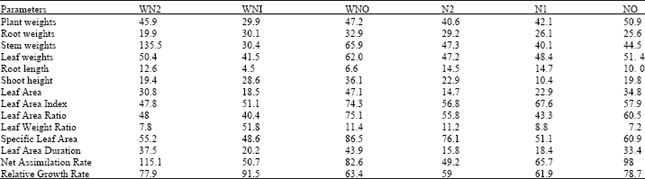 | |
| Table 3: | The root dry weight at five different harvest periods of Macrotyloma geocarpa treated with different levels of fertilizer and weed (values are means with standard error of means) |
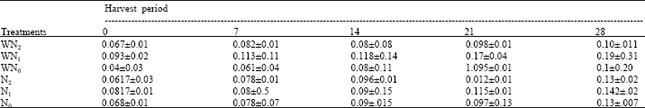 | |
| Table 4: | The stem dry weight at five different harvest periods of Macrotyloma geocarpa treated with different levels of nitrogen fertilizer and weed (values are means with standard error of means) |
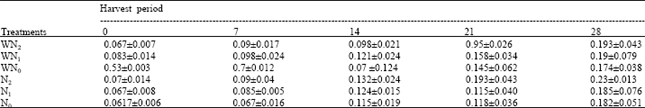 | |
| Table 5: | The leaf dry weight at five different harvest periods of Macrotyloma geocarpa treated with different levels of nitrogen fertilizer and weed (values are means with standard error of means) |
 | |
The coefficient of variation showed that the means of the treatments varied depending on both weed interference and the different nitrogen levels (Table 2).
The dry weight production in roots showed that WN1 plants had the highest dry weight measurements and this was lowest in the roots of the WN0 plants throughout the duration of the experiment. In the other treatments root weights were not stable as they varied from day zero till the 28th day (Table 3).
| Table 6: | The root length at five different harvest periods of Macrotyloma geocarpa treated with different levels of nitrogen fertilizer and weed (values are means with standard error of means) |
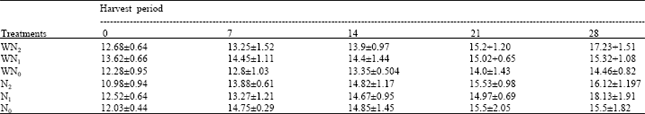 | |
| Table 7: | The shoot height at five different harvest periods of Macrotyloma geocarpa treated with different levels of nitrogen fertilizer and weed (values are means with standard error of means) |
 | |
| Table 8: | The leaf area at different harvest periods of Macrotyloma geocarpa treated with different levels of nitrogen fertilizer and weed (values are means with standard error of means) |
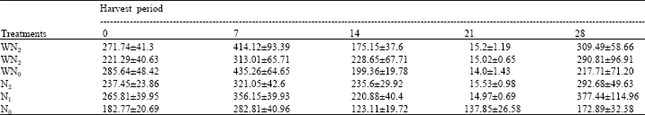 | |
| Table 9: | The leaf area index at five different harvest periods of Macrotyloma geocarpa treated with different levels of nitrogen fertilizer and weed (values are means) |
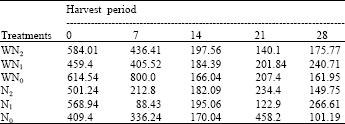 | |
| Table 10: | The leaf area ratio at five different harvest periods of Macrotyloma geocarpa treated with different levels of nitrogen fertilizer and weed (values are means) |
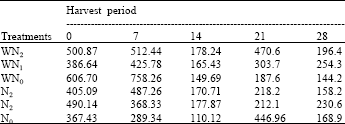 | |
At the end of the experiment, the root dry weight of WN1 plants were highest followed by N1, N2, N0, WN2 while WN0 were lowest. N0 and N1 plants had means that were close irrespective of the different nitrogen levels, WN1 and N2 we re also close while WN2 and WN0 plants had means that vary irrespective of weed competition and nitrogen levels.
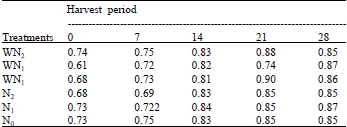
| Table 11: | The leaf weight ratio at five different harvest periods (day after planting) of Macrotyloma geocarpa treated with different nitrogen fertilizer and weed competition (values are means) |
 | |
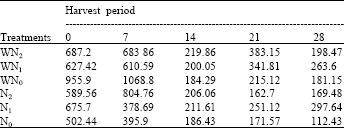
| Table 12: | The specific leaf area at five different harvest periods of Macrotyloma geocarpa treated with different levels of nitrogen fertilizer and weed (values are means) |
 | |
The stem dry weight of all the treatments increased appreciably throughout the experiment. There was no variation in the treatment means of the plants under different nitrogen levels, while there was a wide variation in the plants under both weed competition and different nitrogen levels (Table 4).
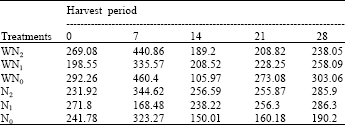
| Table 13: | The leaf duration at five different harvest periods of Macrotyloma geocarpa treated with different levels of nitrogen fertilizer and weed (values are means) |
 | |
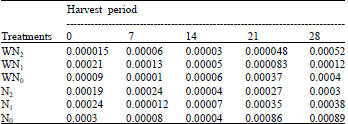
| Table 14: | The net assimilation rate at five different harvest periods of Macrotyloma geocarpa treated with different levels of nitrogen fertilizer and weed (values are means) |
 | |
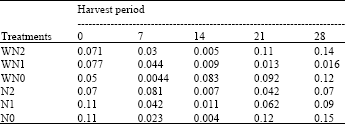
| Table 15: | The relative growth rate at five different harvest periods of Macrotyloma geocarpa treated with different levels of nitrogen fertilizer and weed (values are means) |
 | |
The leaf dry weight of all treatments increased appreciably throughout the experiment. N1 plants had the highest leaf dry weight followed by N2, WN2, N0, WN0 while the WN1 plants had the lowest leaf dry weight at the end of the experiment. There was no variation in the treatment means (Table 5).
The plants that were treated with nitrogen levels only had means that were closer than those treated with weeds.
The roots of the plants were observed to increase with plant age. The lengths of roots were approximately the same in the six treatments. Root lengths were highest in the N1 plants followed by WN2, N2, WN0 and WN1 and lowest in N0 plants. The coefficient of variation showed that root length means of plants treated with different nitrogen levels do not vary, while this varies in plants treated with different nitrogen levels and weed interference (Table 6).
Shoot heights increased gradually with plant age in all the treatments from the zero day till the end of the experiment. WN0 plants recorded the highest shoot height followed by WN1, N2 N0 and WN2 while N1 plants recorded the lowest. The heights of WN2 and N0 plants were approximately the same at the end of the experiment (Table 7). The coefficient of variation showed that the means of the treatments varied depending on the weed interference but independent of the nitrogen levels.
There was an initial increase followed by a decrease and later an increase in leaf area of M. geocarpa due to fertilizer application and weed competition throughout the period the experiment lasted. There was variation in all the treatments (Table 8).
There was decrease in LAI in all treatments up till the 14th day followed by a gradual increase till the end of the experiment except in N0 plants that recorded a sharp decrease. There was a high variation in the leaf area index of the plants that were treated with both nitrogen levels and weed competition (Table 9).
Fluctuations that involve increases and decreases were recorded in all the treatments throughout the experiment. LAR was highest in WN1 followed by NI, N0, N2, WN0, while WN2 was lowest. The degree of variability in the LAR was dependent on both the nitrogen levels and weed competition (Table 10).
There was fluctuation in the levels of LWR in M. geocarpa plants that were exposed to different levels of nitrogen fertilizer band weed competition. It was observed that the N1 plants recorded the highest LWR for a greater part of the experiment followed by WN1, WN2 and N1, while the lowest LWR was recorded in N0 and N2 plants towards the end of the experiment. There was no variation in the LWR in all treatments except the WN1 plants that varied from the other treatments. Decrease in Specific Leaf Area (SLA) was recorded up to the 14th day followed by a gradual increase up to the end of the experiment in all the treatments. At the end of the experiment the SLA was highest in N1 plants followed by WN1, WN2, WN0 and N2 while N0 had the lowest specific leaf area. There was variation in the specific leaf area of all treatments (Table 11, 12).
Apart from N1 plants there was a decrease in leaf area duration (LAD) virtually all the treatments up till the 14th day followed by an increase till the end of the experiment. At the end of the experiment WN0 plants recorded the highest LAD followed by N2, N1, WN1and WN2, while the lowest LAD was observed in the control (i.e. N0 plants). There was a lot of variation in the LAD of all the treatments (Table 13).
Apart from WN0 plants that recorded a decrease in Net Assimilation Rate (NAR) only in the first 7 days, there was a decrease in all treatments up to the 14th day of the experiment followed by a gradual increase till the end of the experiment. At the end of the experiment, N0 plants recorded the highest NAR followed by WN2, N1, WN0 and N2 plants, while WN1 plants had the lowest NAR values. There was a lot of variation in the means of NAR of all the treatments (Table 14).
There was an initial decrease in the Relative Growth Rate (RGR) of all treatments followed by a gradual increase till the end of the experiment. Initially, RGR was highest in N0 and N1 followed by WN1, WN2 and N2, while WN0 plants had the lowest RGR. There was a variation in the RGR of all treatments. Variation was greater in WN1 plants than in other treatments (Table 15).
DISCUSSION
This study sets out to highlight, the effects of interspecific competition, nitrogen fertilizer application and effects of the two (weed + fertilizer) treatments combined on the overall growth, performance and reproductive yield of Macrotyloma geocapa. All the investigations were carried out under the same experimental and environmental conditions, except for the variation in the doses of nitrogen fertilizers and weed interference. Any observed differences therefore in the growth parameters of the crop, in the control experiment (N0 – no weed and no fertilizer) and other treatments were due to the effects of the applied treatments. The high significant differences obtained in the overall growth parameters of the crop between nitrogen fertilizer treatments at different levels imply that crop grain yield performance is predominantly determined by a constant ratio of growth rate to relative nitrogen uptake as the productivity in the control was lower than that in which there was application at the end of the experiment. This agrees with Ezedinma (1964) that there is a linear relationship between growth rate of a crop and the relative nitrogen uptake. Nitrogen fertilizer will affect the rate of this process only when the nitrogen fertilizer increases the external (soil) concentration of nitrogen. This declines with crop age. Growth response to nitrogen application was observed throughout. Ezedinma (1964), cited by Osiname (1978) noted that nitrogen application to cowpea at planting eliminated any retardation in growth and development that might follow the loss of cotyledons shortly after emergence and before the nodules become functional.
A good index for measuring the growth and performance of active tissues is the dry weight. The dry weight of the roots of the fertilized Macrotyloma geocarpa plants was higher than that of untreated ones that is without nitrogen fertilizer. This observation is in agreement with that of Osiname (1978) on the application of nitrogen fertilizer, which significantly increased the grain yield of cowpea variety Ife- Brown. The nitrogen fertilizer applied at planting acted primarily as a growth booster, with significant effect on yield. Simpson et al. (1983) also stated that the pathway of recently acquired nitrogen is not unidirectional from roots to tops but involves cycling of a variable proportion of total amino acid through the phloem to the roots.
The results obtained for the dry weight of the shoots (i.e, stem and leaves) also agreed with the observation that roots biomass increase with plant age. The higher weights of the shoots can be interpreted to mean greater metabolic activities in the shoots of the plants regardless of the different levels of fertilizer application. The dry weight of an organ also reflects the rate of movement of solutes in to that organ. Under dry conditions, nitrogen efficiency at harvest generally ranges from 20 to 80%, depending on factors such as fertilizer type as explained by Monem Abdel (1986).
Optimum response to fertilizer as well as overall yield levels depends on time of sowing. Mactotyloma geocarpa was sown in the month of June 1998 at the onset of rains. This agreed with an observation in a two-year experiment on pearl millet (Pennisetum typhoides) in the month of June 1985 and this was repeated in 1986. For example, Bandyopadhyay and De (1986) showed that with timely sowing (i.e. at the onset of rains in early June), nitrogen recovery was 23% compared with only 17% when sowing was at the end of July.
However the Monsoon rains start late, delay in sowing, which coincides with moist soil conditions, is inevitably the overall best prize.
The time of application of nitrogen fertilizer has considerable influence on the yield of rain-fed crops. Broadcasting the nitrogen was practiced on the 18th day after planting or sowing for Macrotylome geocarpa plants. This is in line with an experiment conducted at several locations on cultivar fields in India. Spratt and Chowdhury (1978) reported that broadcasting 50% of fertilizers at the point of sowing and 50% at 25 to 50 days after sowing (depending on soil moisture availability) was advantageous for upland rice (Oryza sativa), pearl millet (Pennisetum typhides), sorghum (Sorghum bicolor), maize (Zea mays) and cowpea (Vigna unguiculata) and this is generally recommended.
The photosynthetic rate depends on the descriptive growth and partitioning functions (Beadle, 1985). In this study LA, NAR, LAR, LAI, LAD and SLA in the unfertilized plants that is N0 and WN0 plants especially on the 14th and 28th days of the experiment recorded reduction in values.
Adequate nitrogen supply increased the dry matter yield under light, which can be attributed to optimal rate of photosynthesis, increase in leaf surface area and higher chlorophyll content. This showed that leaves of these plants retained a relatively high photosynthetic capacity irrespective of the nitrogen levels. This justifies the high photosynthetic capacity irrespective of the nitrogen levels. This justifies the high photosynthetic and high growth rates recorded at h igh nitrogen fertilizer levels aided by irradiance. This supports the opinion of Holt (1991) that whole plant productivity depends on the LAI and allocation of resources. It equally agrees with Nobel (1983) that in a low photo synthetically active radiation (PAR) environment, plants would be tall and consequently possess a high LAR. The low LWR in and LAR in fertilized plants with high irradiance showed that there was diversion of a greater fraction photosynthate to other organs like root and reproductive structures. The current study confirms and extends previous observations of Vos and Biennond (1992) and Vos (1995) showing marginal effects of nitrogen supply in the rate of leaf appearance and specific leaf weight. The greater proportion of the photosynthates would be locked up in the leaves where they are made. Full frown leaf area is affected by the duration of expansion in response to nitrogen supply (Vos, 1995). A combination of interspecific competition and fertilizer stress was observed to cause a reduction in biomass probably due to the prevailing lower PAR and temperature by two stresses. Weed which is the non-leguminous competitor could not fix N biologically. Thus less soil N was available for growth in the mixed culture contrary to the monoculture where all the individuals did fix nitrogen. The effect of fertilizer application and weed interference was shown in WN0 plants.
Gallagher (1978) reported that legumes would suppress weed growth once a complete canopy cover is obtained. The above result showed that the weight of the weed in the mixture was lower compared to that of the crop. The result of the present study agrees with the above assertion, especially at the critical period of 14th and 21st days (that is 39-46 days after planting). The crop treated with nitrogen fertilizer required a high biomass to significantly suppress the weed, Euphorbia heterophylla. This was undoubtedly due to the increases in the crops height, LA and deeper and more branched root system (Fitter and Hay, 1987; Bozsa and Oliver, 1990; Compbell et al., 1992).
Initially seedling size (as determined by seed size) and relative growth rate together result in a given seedling biomass at any point in time, seed size being the more important factor soon after germination and RGR the more important later, in good growth conditions (Savarimuttu, and Westoby, 1996). It may equally be due to the crop’s larger seed size and weed smaller size (Patterson, 1990) or uniform germination (Frazee and Stoller, 1974). The critical period of weed interference in a crop therefore depends on the growing conditions and stage of growth of the crops. Weed competition is most serious and when cowpea crop is young (Fadayomi and Abayomi, 1988).
The significant continuous reduction of root length by intraspecific competition reported by Donald (1958), Wilson and Newman (1987), which may be due in part to the fact that root competition may be more intense than shoot competition, negated the observation recorded in the present experiment. Instead of a reduction there was a slight increase in both root length and shoot height. Improved soil physical conditions may promote root growth and increased use of soil nitrogen fertilizer, which lead to high yield.
The residual effects of below ground legume biomass i.e, roots and nodules reflected on root length and shoot height. It has generally been observed that in plant, the nitrogen required for spring growth can be obtained from two sources, re-mobilization of stone material and root uptake, in which nitrogen was re-mobilized from roots and swollen basal internodes to supply shoot growth primarily before the onset of root uptake (Thornton and Milland, 1993). The above ground legume biomass (i.e, shoot) generally accounts for most of the legume leaves, stem and flowers contain considerable quantities of nitrogen. George et al. (1992; 1995) found out that the roots of mungbean (Vigna rodiata), Soybean (Glycine max L.) and sesbania (Sesbania sesban) species that contain an average of 40 kg N ha-1 for fertilizer rate had an increase in the grain yield. The observable reports of many researches on percentage of fertilizers to the hectare of the land are as follows: The fertilizer rates were 0, 20 and 40 kg N ha-1, Singh (1973) used 15, 30 and 45 kg N ha-1 + rhizobial inoculation. From the above reports, the weight of nitrogen fertilizer (15 and 30 kg N ha-1) applied in this experiment is in line with others on legumes.
The natural means of controlling Euphorbia heterophylla and related weed species in Macrotyloma geocarpa fields may be inter-specific suppression of the weed by Macrotyloma geocarpa due to its spreading ability. The improvement of these crops will further increase the protein base of the poor people who cannot afford animal protein. This is needed with other ground bean varieties such as Macrotyloma uniflorum, Vigna subterranes, Vigna poisseniii (L.) Verde (Bambara groundnuts) and weed species with other types of fertilizers at different levels to widen scope of this investigation of the effects of weed competition and fertilizer application on ground beans.
REFERENCES
- Ayeni, A.O., W.B. Dduke and R.O. Akobundu, 1984. Weed interference in maize, cowpea and maize/cowpea intercrop. Weed Res., 24: 287-290.
Direct Link - Bozsa, R.C. and L.R. Oliver, 1990. Competitive Mechanisms of common cocklebur. (Xanthium strumarium) and Soybean (Glycine max) during seedling growth. Weed Sci., 38: 344-350.
Direct Link - Compbell, B.D., J.P. Crime, J.M.C. Mackey, 1992. Shoot thrust and its role in plant competition. J. Ecol., 80: 333-341.
Direct Link - Donald, C.M., 1958. The interaction of competition for light and for nutrients. Aust. J. Agric. Res., 9: 421-435.
CrossRef - Frazee, R.W. and E.W. Stoller, 1974. Differential growth of Soybean and seven dicotyledonous weed seedlings. Weed Sci., 22: 336-339.
Direct Link - George, T., J.K. Ladha, R.J. Buresh and D.P. Garrity, 1992. Managing native and legume-fixed nitrogen in lowland rice-based cropping systems. Plant Soil., 141: 69-91.
CrossRef - Hepper, F.N., 1963. Plants of the 1957-58 W.A. Expedition. The bambarra groundnut (Voadzeia subterraneal) and kerstings groundnut (Kerstigiella geocarpa) Wild. Kew Bull., 16: 395-407.
Direct Link - Holt, J.S., 1991. Application of physiological ecology to weed science. Weed Sci., 39: 521-528.
Direct Link - Johansen, C., B. Balder, J.B. Brouwer, W. Erskine, W.A. Jermyn and L. Li-Juan, 1992. Biotic and Abiotic Stresses Constraining Productivity of Cool Season Food Legume in Asia. In: Africa and Oceanic Production and Use of Cool Season Food Legumes, Muehlbauer, F.J. and W.J. Kaiser (Eds.). Kluwer Academic Publishers, Dordrecht, pp: 175-194.
- Sheehy, J.F., F. Gastal, P.L. Mitochell, J.L. Durand, G. Lemaire and L. Woodward, 1996. A nitrogen led model of grass growth. Ann. Bot., 77: 167-177.
Direct Link - Simpson, R.J., H. Lambers and M.J. Dalling, 1983. Nitrogen redistribution during grain growth in wheat (Triticum aestivum (L.)) IV. Development of a quantitative model of the translocation of nitrogen to the grain. Plant Physiol., 71: 7-14.
Direct Link - Vos, J. and H. Biemond, 1992. Effects of nitrogen on the development and growth of the potato plant. 1. Leaf appearance, expansion growth, life spans of leaves and stem branching. Ann. Bot., 70: 27-35.
CrossRefDirect Link