
Abd El-Zaher M.A. Mustafa
Department of Botany, Faculty of Science, Tanta University, 31527 Tanta, Egypt
Abdelfattah Badr
Department of Botany, Faculty of Science, Tanta University, 31527 Tanta, Egypt
Mohammed A. El-Galaly
Department of Botany, Faculty of Science, Benha University, Benha, Egypt
Ahmed A. Mobarak
Department of Botany, Faculty of Science, Benha University, Benha, Egypt
Mervet G. Hassan
Department of Botany, Faculty of Science, Benha University, Benha, Egypt
International Journal of Botany
Year: 2006 | Volume: 2 | Issue: 3 | Page No.: 261-269







ABSTRACT
Seven populations of two species of Ocimum were analyzed using data from morphological and biochemical (seed proteins and isozymes) characters to measure genetic variation within and between populations and species of Ocimum in Egypt. Ten isozyme systems revealed 23 loci and a total of 49 alleles in the studied Ocimum populations. The constructed trees based on variation in morphological, seed proteins or isozyme data clearly demonstrated the existence of genetic diversity among and within populations of Ocimum that might be related to natural hybridization and fluctuations in environmental conditions. Seed proteins and isozyme polymorphism exhibited validity for studying genetic diversity and taxonomic relationships in Ocimum at both species and infra-specific levels. Moreover, Ocimum kilimandscharicum exhibited higher levels of genetic variation and also higher number of unique alleles than Ocimum basilicum.
PDF Abstract XML References Citation
How to cite this article
Abd El-Zaher M.A. Mustafa, Abdelfattah Badr, Mohammed A. El-Galaly, Ahmed A. Mobarak and Mervet G. Hassan, 2006. Genetic Diversity among Ocimum Populations in Egypt as Reflected by Morphological, Seed Proteins and Isozyme Polymorphism. International Journal of Botany, 2: 261-269.
DOI: 10.3923/ijb.2006.261.269
URL: https://scialert.net/abstract/?doi=ijb.2006.261.269
DOI: 10.3923/ijb.2006.261.269
URL: https://scialert.net/abstract/?doi=ijb.2006.261.269
INTRODUCTION
The genus Ocimum (basil) comprises same of the most popular herbs in the world. It belongs to the family Lamiaceae, subfamily Ocimoideae and includes over 150 different species and varieties distributed in tropical regions of Asia, Africa, Central and South America (Darrah, 1980). Because of its popularity, basil is often referred to as the "king of the herbs, being widely utilized due to its economic, nutritional, industrial and medicinal importance (Simpson and Conner, 1986; Simon et al., 1990; Khosla, 1995). Volatile compounds produced by sweet basil were shown to have allelopathic effects and influence the composition, distribution and spore germination of some fungal populations (Reuveni and Putiersky, 1984).
Inter-specific hybridization and polyploidy are of common occurrence within the genus Ocimum and have created taxonomic confusion and challenges (Anonymous, 1980). The morphological diversity within basil has been accenturated by centuries of cultivation with great variation in pigmentation, leaf shape and size (Anonymous, 1980). Using the morphological data in studying genetic diversity and taxonomic relationships of plants was previously scored for many plants (Bult and Kiang, 1992; Zviniene and Pank, 1996).
In addition to morphological traits, phytochemical analysis and chromosomal criteria, seed protein electrophoretic patterns have provided valid evidence for addressing genetic, taxonomic and evolutionary relationships in plants. Evidences provided by the separation of seed protein components in polyacrylamide gels are mostly informative at the species and infra-specific levels (Nei et al., 1978; Ladizinsky and Hymowitz, 1979; Cooke, 1984; Badr, 1995). Seed proteins data have therefore been applied to study species relationships in many plant genera (Boulter et al., 1970; Ladizinsky, 1979a, b; Sammour, 1989, 1994; Sammour et al., 1991; Paino et al., 1990; Saraswati et al., 1993; Badr, 1995) and genetic diversity at infraspecific levels (Badr et al., 1998; Al-Nowaihi et al., 2002; Badr et al., 2003).
Many previous applications have exploited the ability of electrophoresis to provide efficient data about the variation, heterozygosity and divergence of alleles at many gene loci coding enzymes in large population samples. Most isozyme data have been applied to species or infraspecific taxa rather than at higher taxonomic ranks (Kang and Chung, 2000; Badr et al., 2002; Mustafa et al., 2005).
Genetic variation and taxonomic relationships in the genus Ocimum were previously investigated using morphological, chemical, cytological and molecular traits (Khosla, 1995; Grayer et al., 1996; Paton and Putievsky, 1996). Estimating the genetic diversity levels both within and among populations of a crop is necessary for the best conservation of its gene pool (Votava et al., 2002). The genetic variation found within wild relatives of domesticated species offer novel gene complexes for strategic improvement of crop tolerance to biotic and abiotic stresses (Votava et al., 2002). Such variation can also have important implications for their conservation and management (Votava et al., 2002). However, no previous studies utilizing electrophoretic variation of seed proteins or isozymes in taxonomic, genetic or phylogenetic studies on Ocimum are known. The objectives of this study is to investigate genetic diversity in Ocimum populations in Egypt as inferred by variations in their morphology and electrophoretic profiles of both seed proteins and isozymes as a necessary step for best conservation of Ocimum gene pool in Egypt and for future improvement of crop characters.
MATERIALS AND METHODS
Samples collection: This study was conducted at Botany Department, Faculty of Science, Tanta University at 2003-2004. Complete mature plants were collected from a number of localities representing natural populations of two species of the genus Ocimum in Egypt. Ocimum basilicum was represented by 3 varieties, var. grand vant (OB1-OB3), var. parpurascens or dark opel basil (OP) and var. finoverde (OF1 and OF2), while O. kilimandscharicum was represented by only one accession (OK). Mature plants were collected during the flowering season for measuring morphological traits and mature seeds were collected for electrophoretic analysis of both seed proteins and isozymes.
Morphological traits: For each accession, the following morphological traits were scored; stem height, width, diameter and habit, internode number and length, leaf width, inflorescence length and width, number of flowers, root width, diameter and length and germination period. Each trait was scored as a mean value of ten individuals for each accession.
Seed proteins: From each population, a composite sample of mature dormant seeds was mixed with an equal weight of sterile fine sand, ground well in a mortar, then defated several times with acetone in the refrigerator until oils could not be observed. The meal was air-dried and samples of 400 mg were extracted overnight at 4°C in 1 mL 0.125 M Tris-borate buffer (pH 8.9) containing 2% β-mercaptoethanol. Extracts were centrifuged at 12000 × g for 20 min, 10% sucrose was added to the supernatant that was used directly for electrophoresis or kept at -20°C until use.
Dissociating polyacryamide gel electrophoresis (SDS-PAGE) was adopted after Laemmli (1970) with some modifications. Samples of 100 μL were applied onto main gels of 17% acrylamide concentration using bromophenol blue as a front dye. Electrophoresis was carried out using 25 mM tris/glycine buffer pH 8.3. The gel was stained overnight by slow shaking in 100 mL staining mixture (0.05% Coammassie Brillient Blue-R250 in 50% methanol, 7% glacial acetic acid and 43% distilled water) and destained by slow shaking in the stain solvent. The subunit molecular weight of the protein bands was determined according to Weber and Osborne (1969). Each band was considered as a character for which the presence or absence was coded 1 or 0 respectively in a data matrix for numerical analysis. Density of the protein bands were not considered.
Isozymes: Seeds were germinated and a leaf specimen (0.25 g) of one seedling was used to prepare samples for isozyme electrophoresis according to Marshall and Brown (1975). Twenty enzyme systems were assayed for activity; nevertheless, valid results were obtained for only ten enzyme systems including five esterases. The staining recipes of these enzymes are given in Table 1. About 4-6 individuals were used as replica from every accession for each enzyme (Table 2).
Isozymes were separated on 8% PAGE vertical slab gels (16×18×0.2 cm) according to Wendel and Weeden (1989). The gel buffer composed of 45 mM Tris-HCl, 25 mM boric acid and 1 mM EDTA-Na2 pH 8.6 and the electrode buffer was composed of 0.18 M Tris-HCl, 0.1 M Boric acid and 4 mM EDTA-Na2. The gels were stained by shaking in the dark at 37°C in the appropriate staining solution (Table 1). After staining, the reaction was stopped by washing the gel 2-3 times with tap water. The gel was then kept in the fixing solution (glycerol and water 1:1 v/v) for 24 h and rinsed two times in tap water, then photographed. Interpretation of banding patterns followed standard principles (Wendel and Weeden, 1989; Murphy et al., 1996). Loci were numbered consecutively from the anodal end and alleles at each locus were labeled alphabetically at the same direction (Pasteur et al., 1988). Alleles were directly scored for each isozyme locus and allele frequencies were calculated. Based on allele frequencies, the following estimates of genetic variation were calculated for each population: (1) Proportion of polymorphic loci (P), (2) mean number of alleles per polymorphic locus (Kp) and (3) the mean number of alleles per locus (K).
| Table 1: | The enzymes assayed and their staining recipes |
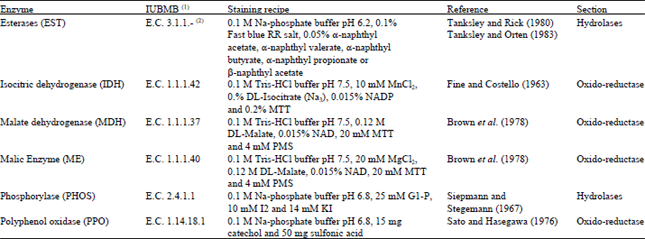 | |
| (1) Nomenclature Committee of the International Union of Biochemistry and Molecular Biology, (2) Non-specific enzyme | |
| Table 2: | Number of studied individuals (N), proportion of polymorphic loci (P), mean number of alleles per locus (K), mean number of alleles per polymorphic locus (Kp) and number of unique allels in accessions of Ocimum sp. |
 | |
Data analysis: Cluster analyses of the data obtained from morphology, seed proteins or isozyme polymorphism were made using the software SPSS for windows package (Version 10). A tree illustrating the genetic distance among populations was constructed based on Euclidean squared distance, Kulczynski Measure 2 and Dice similarity measure for morphological, seed proteins or isozyme data, respectively.
RESULTS
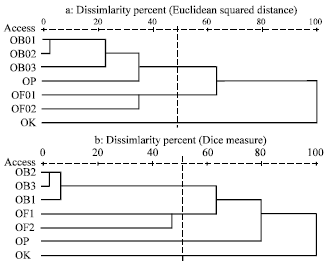
Morphological traits: Figure 1 and Table 3 illustrate the genetic distance among the studied accessions of Ocimum based on morphological traits, it can be noticed that farthest genetic distance was exhibited between O. kilimandscharicum and O. basilicum var. fino verde, while nearest distance was exhibited among the accession of the same variety or between sweet basil (OB) and dark opel basil (OP). O. basilicum var. grand vant and var. parpurascens exhibited similarity to O. kilimandscharicum more than to var. fino verde.
Seed proteins: The results obtained in this study showed a large variation in the number of protein bands among the studied accessions (Fig. 2 and Table 4), the least number of bands (12) was found in O. basilicum var. purpurascence (OP) and the highest number (36) for sweet basil (O. basilicum var. grand vant OB2) with molecular weight between 60 and 20 KDa.
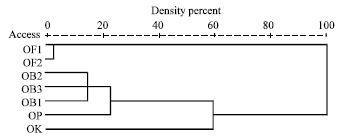 | |
| Fig. 1: | Dendrogram based on Ocimum morphological data |
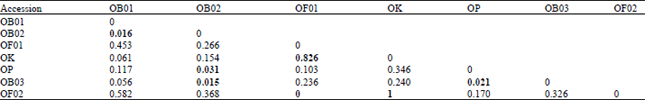
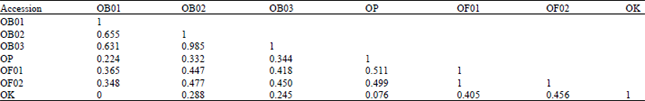
The accessions of O. basilicum exhibited four bands that could not be scored in O. kilimandscharicum, which showed four bands that could not be scored by O. basilicum. The cluster analysis using Kulczynski Measure 2 (Table 5) showed highest similarity within each variety and farthest genetic distance between OK and both OB01 and OP. These data were best represented in the dendrogram based on Kulczynski measure 2 similarity index (Fig. 3), which exhibited best discrimination between the two Ocimum species and also among the three varieties of O. basilicum.
Isozymes: Ten enzyme systems and 23 loci were resolved and scored in the studied seven accessions of Ocimum One of these ten enzymes (IDH) was represented by only one locus while the remaining nine enzymes were represented by 2-3 loci each.
| Table 3: | The genetic distance among Ocimum accessions based on morphological data |
 | |
| Table 4: | The protein bands in the electrophoregram of the studied Ocimum accessions |
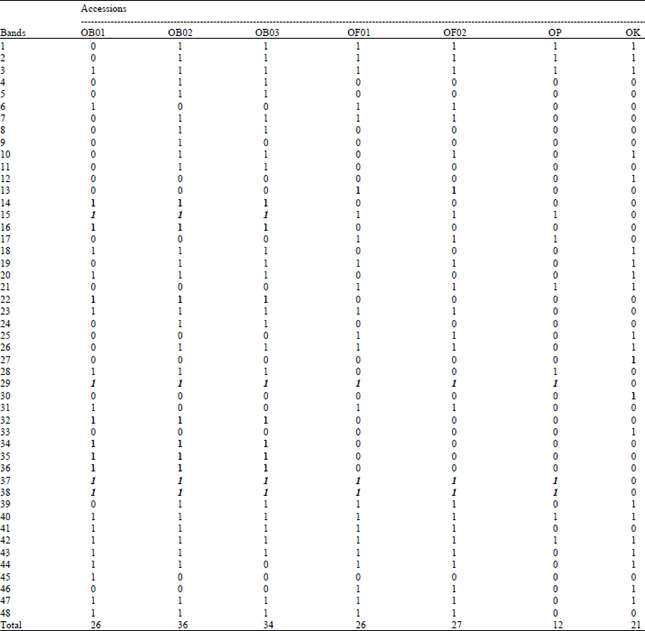 | |
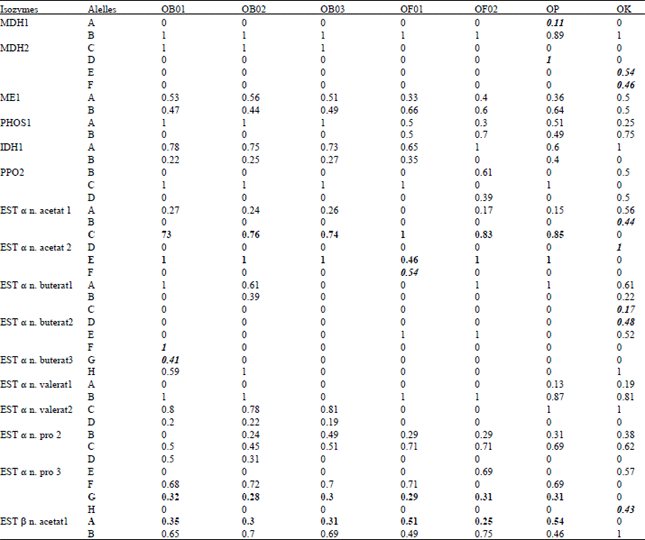
Seven of the 23 loci were monomorphic in all the studied accessions of Ocimum (ME 2, PHOS 2, PPO1, EST α-naphthyl valerate 3, EST α-naphthyl propunate 1 and EST β-naphthyl acetate 2 and 3), while the remaining 16 loci were polymorphic (Table 6 and 7). Twelve unique alleles were scored in the studied accessions of Ocimum sp. (Table 2 and 6), seven of which in O. kilimandscharicum, two in each of the accessions OB1 and OP and only one in OF1.
| Table 5: | The genetic similarity index among the accessions of Ocimum sp. based on seed protein data. (Kulczynski Measure 2) |
 | |
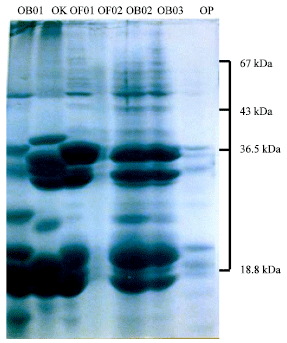 | |
| Fig. 2: | Seed protein electrophoretic profile of Ocimum accessions |
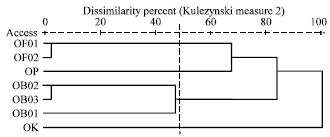 | |
| Fig. 3: | Dendrogram based on electrophoretic data of Ocimum seed proteins |
The accession representing O. kilimandscharicum exhibited higher values of genetic variation parameters (P, K, Kp) than the accessions of O. basilicum (Table 2).
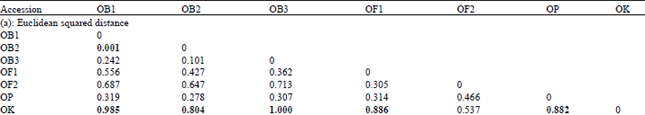
Cluster analysis of the allele frequency data using Euclidean squared distance and Dice similarity measure (Table 8) showed farthest genetic distance between O. kilimandscharcum and both of the accessions of O. basilicum except the accession OF02. Minimal Euclideane distances were noticed within each variety or between var. purpurascence (OP) and var. grandvant (OB01, OB02 and OB03) of O. basilicum.
 | |
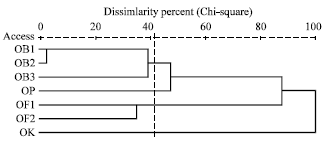
| Fig. 4: | Dendrogram Based on electrophoretic data of Ocimum isozymes |
 | |
| Fig. 5: | Dendrogram based on data of morphology and electrophoretic polymorphism of seed proteins and isozymes |
The dendrograms based on either Euclidean squared distance or Dice similarity measure (Fig. 4) showed best discrimination of the accessions under study at both species and infra-specific levels. At 50% similarity, the accessions were classified according to Dice similarity measure into four categories representing the three cultivars of O. basilicum and the accession of O. kilimandscharcum while they were grouped into three categories according to squared Euclidean distance aggregating the two varieties grandvant and purpurascence in one group.
Cluster analysis of the combined data (morphology, seed proteins and Isozymes) depending on Chi-square between sets of frequencies (Table 9 and Fig. 5) showed farthest genetic distance between O. kilimandscharcum and each of the accessions of O. basilicum and also between the two varieties of O. basilicum; grandvant and finoverde.
| Table 6: | Allele frequency of polymorphic loci of the studied Ocimum samples; unique alleles are shaded and italic |
 | |
| Table 7: | Number of polymorphic and monomorphic loci for each isozyme studied |
 | |
| Table 8: | The genetic relationships among Ocimum accessions based on isozyme polymorphism |
 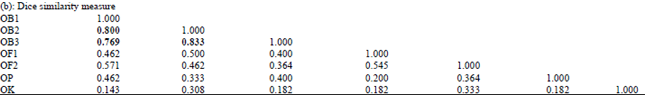 | |
| Table 9: | The genetic distance (Chi-square) among Ocimum accessions (total data) |
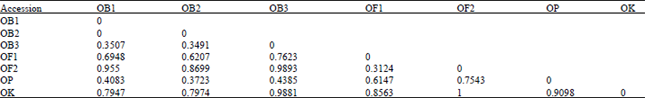 | |
Nearest genetic distances were revealed among the accessions of sweet basil. At 45% dissimilarity, the accessions were grouped into four categories representing the three varieties of O. basilicum and the accession of O. kilimandscharcum
DISCUSSION
The importance of morphological variation among the examined populations of Ocimum is demonstrated by the topology of the dendrogram based on Euclidean distance that showed discrimination of the two species under study. The separation of the two accessions of O. basilicum var. fino verde from all other accessions in one cluster is correlated with clear differences in most of the scored morphological characters. Grouping the accessions of each cultivar in one cluster in spite of the different environmental conditions of each accession reflects a genetic basis of the plant form in Ocimum.
The large variation in the number of protein bands among the studied Ocimum accessions reflects the high level of genetic variation among and within Ocimum populations in Egypt. The presence of certain protein bands characteristic to certain varieties or species, in addition to the topology of the tree illustrating the genetic distances among the studied Ocimum accessions based on the data obtained from the electrophorertic analysis of the seed proteins, supports the validity of the used technique (seed protein electrophoresis) as a tool for cultivar identification as well as studying genetic diversity and taxonomic relationships in Ocimum at both specific and infra-specific levels. Electrophoresis of seed proteins was previously used for cultivar identification for many other plants; as Vicia faba (Stegemann et al., 1980) and Linum usitatissimum (Sammour, 1988). It was also utilized for studying genetic diversity and addressing taxonomic relationships in Glycine soja (Bult and Kiang, 1992), Lens (Ladizinsky, 1979a, b; Sammour, 1994), Vicia faba (Sammour, 1989), Lotus (Sammour et al., 1991), Sesbania (Saraswati et al., 1993) and Mentha (Badr et al., 2003).
The dendrograms based on data obtained from isozyme polymorphism and allele frequency according to Euclidean distance or Dice measure reflect the intra-and inter-populational genetic variation in the genus Ocimum in Egypt. The higher genetic polymorphism in a population or a species is dependent on the amount of sexual reproduction, whereas low levels of genetic variation are often associated with asexual propagation (Micales and Bonde, 1995; Mustafa et al., 2005). According to this view, the results obtained in this study that showed lower genetic variation parameters for O. basilicum than those of O. kilimandscharicum might reflect the relative equilibrium state between sexual and vegetative reproduction that tends towards higher percentage of sexual propagation in O. kilimandscharicum. The topology of the tree based on the isozyme data in addition to such large number of unique alleles reflect the validity of the isozyme data to study genetic diversity and taxonomic relationships at both species and infra-specific levels in Ocimum. Such discrimination at the species and infra-specific levels using electrophoretically assayed isozyme variation was previously scored by many authors for different plant genera (Schmit et al., 1996; Kang and Chung, 2000; Batista et al., 2001; Badr et al., 2002; Mustafa et al., 2005). The very few number of cultivar diagnostic alleles within O. basilicum suggests that the cultivars under study might be hybrids or have recently derived from an ancestor harboring high levels of genetic diversity (Kang and Chung, 2000; Mustafa et al., 2005). This derivation might be due to intra-specific natural hybridization and the subsequent dispersion of pollen grains and hybrid fruits or seeds. Inter-specific hybridization within Ocimum in Egypt may be also indicated by the farther genetic distance within O. basilicum than between some of its accessions and O. kilimandscharicum. The inter- and intra-specific natural hybridization was previously reported for Ocimum (Anonymous, 1980; Khosla and Sobti, 1984) and other plant genera (Aparicio et al., 2000; Ellstrand and Schierenbeck, 2000; Gobert et al., 2002; Mustafa et al., 2005).
The grouping of the accessions of each cultivar in one cluster in each of the trees based on data inferred from electrophoretic seed protein or isozyme polymorphism or both with morphological data may indicate the impact of environmental conditions on genetic variation rather than genetic divergence in Ocimum populations.
REFERENCES
- Al-Nowaihi, A.S., S.F. Khalifa, A. Badr and S.M. Sharawy, 2002. Species relationships of Astragalus L. in Egypt based on storage seed protein electrophoretic criteria. Proceedings of the 2nd International Conference on Biology Science, Volume 2, April 27-28, 2002, Tanta Univeristy Egypt, pp: 111-121.
- Aparicio, A., R.G. Albaladejo, M. Porras and G. Ceballos, 2000. Isozyme evidence for natural hybridization in Phlomis (Lamiaceae): Hybride origin of the rare P. x margaritae. Ann. Bot., 85: 7-12.
Direct Link - Badr, A., 1995. Electrophoretic studies of seed proteins in relation to chromosomal criteria and the relationships of some taxa of Trifolium. Taxon, 44: 183-191.
Direct Link - Badr, A., H. Sayed-Ahmed, L.E. Watson and A. El-Shanshouri, 2002. Ancestors of white clover (Trifolium repens L.), as revealed by isozyme polymorphisms. Theor. Appl. Genet., 106: 143-148.
CrossRefPubMedDirect Link - Badr, A., A.M.A. Mustafa, M.A El-Galaly, A. Mobarak and M.G. Hasan, 2003. Genetic diversity among Mentha populations in Egypt as reflected by morphological and electrophoretic variation. Proceedings of the 1st Egyptian-Syrian Conference Agriculture and Food in Arabic Nations, December 8-11, 2003, Menia University, Egypt, pp: 269-286.
- Batista, F., A. Banares, J.C. Castelis, E. Carque, M.M. Gomez and P.A. Sosa, 2001. Allozyme diversity in three endemic species of Cistus (CISTACEAE) from the canary Islands: Intraspecific and Interspecific comparisons and implications for genetic conservation. Am. J. Bot., 88: 1582-1592.
Direct Link - Brown, A.H.D., E. Nevo, D. Zohari and O. Dagan, 1978. Genetic variation in natural populations of wild barley (Hordeum spontaneum). Genetica, 49: 97-108.
CrossRefDirect Link - Bult, C.J. and Y.T. Kiang, 1992. Electrophoretic and morphological variation within and among natural populations of the wild soybean, Glycine soja. Bot. Bull. Acad. Sin., 33: 111-112.
Direct Link - Ellstrand, N.C. and K.A. Schierenbeck, 2000. Hybridization as a stimulus for the evolution of invasiveness in plants. Proc. Natl. Acad. Sci. USA., 97: 7043-7050.
Direct Link - Gobert, V., S. Moja, M. Colson and P. Taberlet, 2002. Hybridization in the section Mentha (Lamiaceae) inferred from AFLP markers. Am. J. Bot., 89: 2017-2023.
CrossRefDirect Link - Grayer, R., G. Kite, F. Goldstone, S. Bryan and E. Putievsky, 1996. Infraspecific taxonomy and essential oil chemotypes in sweet basil (Ocimum basilicum L.). Phytochemistry, 43: 1033-1039.
CrossRef - Kang, S.S. and M.G. Chung, 2000. High levels of allozyme variations and low allozyme divergence within and among species of Hemerocallis (Liliaceae). Am. J. Bot., 87: 1634-1646.
Direct Link - Khosla, M.K. and S.N. Sobti, 1984. Hybridization between different geographical races of Ocimum. Nucleus, 27: 156-159.
Direct Link - Ladizinsky, G. and T. Hymowitz, 1979. Seed protein electrophoresis in taxonomic and evolutionary studies. Theor. Applied Genet., 54: 145-151.
CrossRefDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Mustafa, A.Z.M.A., A. Badr, M.A. El-Galaly, A.A. Mobarak and M.G. Hassan, 2005. Genetic diversity among Mentha populations in Egypt as reflected by isozyme polymorphism. Int. J. Bot., 1: 188-195.
CrossRefDirect Link - Paton, A. and E. Putievesky, 1996. Taxonomic problems and cytotaxonomic relationships between and within varieties of Ocimum basilicum and related species (Labiatae). Kew-Bull., 51: 509-524.
Direct Link - Sammour, R.H., M.A. Hamoud and A.S. Haidar, 1991. Seed protein variation in relation to cytological features of some species in genus Lotus. Cytologia, 65: 289-291.
Direct Link - Schmit, V., D.G. Debouk and J.P. Baudoin, 1996. Biogeographical and molecular observations on Phaseolus glabellus (Fabaceae, Phaseolinae) and its taxonomic status. Taxon, 45: 493-501.
Direct Link - Simon, J.E., J. Quinn and R.G. Murray, 1990. Basil: A Source of Essential Oils. In: Advances in New Crops, Janick, J. and J.E. Simon (Eds.). Timber Press, Portland, pp: 484-489.
Direct Link - Stegemann, H., A.E. Shehata and M. Hamza, 1980. Broad bean proteins (Vicia faba L.). electrophoretic studies on seeds of some German and Egyptian cultivars. Z. Acker Und Pflanzenbau, 149: 447-453.
Direct Link - Tanksley, S.D. and C.M. Rick, 1980. Genetics of esterases in species of Lycopersicon. Theoret. Applied Genet., 56: 209-219.
CrossRefDirect Link - Votava, E.J., G.P. Nabhan and P.W. Bosland, 2002. Genetic diversity and similarity revealed via molecular analysis among and within an in situ population and ex situ accessions of chiltepin (Capsicum annuum var. glabriusculum). Conserv. Genet., 3: 123-129.
CrossRefDirect Link - Weber, K. and M. Osborn, 1969. The reliability of molecular weight determinations by dodecyl sulfate-polyacrylamide gel electrophoresis. J. Biol. Chem., 244: 4406-4412.
Direct Link