
Dhananjay Kumar Yadav
Division of Plant Pathology, Indian Agricultural Research Institute, 110012 New Delhi, India
Dinesh Singh
Division of Plant Pathology, Indian Agricultural Research Institute, 110012 New Delhi, India
LiveDNA: 91.17539
Narendra Kumar
Amity Institutes of Biotechnology, Amity University Haryana, Manesar, 122413 Gurgaon, India
LiveDNA: 91.2651
International Journal of Agricultural Research
Year: 2017 | Volume: 12 | Issue: 4 | Page No.: 172-180







ABSTRACT
Background and Objective: The Bacterial wilt caused by Ralstonia solanacearum (R. solanacearum) (Smith) Yabuuchi is a serious disease of tomato (Solanum lycopersicum L.) in tropical, subtropical and temperate areas of world including India and reduces the crop yields significantly. This study aimed to investigate the antibacterial activity through induction of defense-related enzymes by Bacillus amyloliquefaciens (B. amyloliquefaciens) DSBA-11 to manage bacterial wilt disease of tomato. Materials and Methods: Bacillus amyloliquefaciens DSBA-11was inoculated to two tomato cultivars Pusa Ruby (susceptible) and Arka Abha (moderate resistant) along with pathogen R. solanacearum UTT-25 at root zone of tomato under control conditions in Phytotronat 28°C and relative humidity was 67-75%. The one-way analysis of variance for biocontrol efficiency and yield of tomato was made by the SAS general linear model (GLM) procedure. Results: Minimum bacterial wilt intensity was recorded in Arka Abha (14.98%) treated with B. amyloliquefaciens DSBA-11 followed by Pusa Ruby (21.64%) which was significantly lower than the control treated only with R. solanacearum after 60 days of inoculation. Tomato seedlings of both cultivars cv. Pusa Ruby and Arka Abha treated with DSBA-11 produced significantly elevated production of superoxide dismutase (SOD), peroxidase (POD), polyphenol oxidase (PPO), phenylalanine ammonia lyase (PAL) and catalase(CAT) as compared to control. Conclusion: The enhancement of induced systemic resistance enzyme activities in two tomato cultivars which could involve in suppression of the wilt disease.
PDF Abstract XML References Citation
Received: September 03, 2017;
Accepted: October 09, 2017;
Published: October 20, 2017
How to cite this article
Dhananjay Kumar Yadav, Dinesh Singh and Narendra Kumar, 2017. Induction of Defense-related Enzymes by Bacillus amyloliquefaciens DSBA-11 in Resistant and Susceptible Cultivars of Tomato Against Bacterial Wilt Disease. International Journal of Agricultural Research, 12: 172-180.
DOI: 10.3923/ijar.2017.172.180
URL: https://scialert.net/abstract/?doi=ijar.2017.172.180
DOI: 10.3923/ijar.2017.172.180
URL: https://scialert.net/abstract/?doi=ijar.2017.172.180
INTRODUCTION
Tomato (Lycopersicon esculentum L.) is the world’s largest vegetable crop and known for its special nutritive value and also for it’s wide spread production. Tomato is one of the most important vegetable crops cultivated for its fleshy fruit and considered as an important commercial and dietary vegetable crop. Higher production of this crop is limited by the various pests and diseases in the field. The bacterial wilt disease caused by Ralstonia solanacearum (Smith) Yabuuchi affects 450 host species in 54 botanical families in tropical and subtropical regions of the world1,2. The disease causes very heavy losses varying from 2-90% in different climatic conditions and seasons in India2, particularly in October-November in coastal area3 and August-October in Northern and Eastern parts of India2. As, limited scope for chemical means of managing the disease, biological method using microbes is found very viable approach. Naturally occurring antagonistic rhizobacteria such as Bacillus spp.4, Pseudomonas spp.5 and Streptomyces spp.6 were found potential bioagent for management of bacterial wilt disease of tomato. Various mechanisms such as antibiosis, competition for nutrients and induction of resistance against disease in plants are involved in suppression of wilt disease by these microbes like other Bacillus spp. Bacillus amyloliquefaciens possess several advantages and makes them good candidates of biocontrol agent. Bacillus amyloliquefaciesns produces various antibacterial and antifungal antibiotics such as surfactin, iturin fengycine and polyketide antibiotics etc.7. The biocontrol agents suppress the bacterial wilt disease in plants through development of induced resistance. The polyphenol oxidsae, peroxidase, catalase, superoxide dismutase and phenylalanine ammonia lyase are the major enzymes responsive for decreasing disease response in tomato plants. The phenylalanine ammonia lyase (PAL) and polyphenol oxidsae (PPO) is the main SAR related enzymes in plants and their activities are related to plant resistance8,9. Bacillus subtilis strain AR12 induces resistance in tomato plants against bacterial wilt disease10. Induction of systemic resistance against various disease causing pathogens in crops such as banana, bean, rice and cucumber by species of Bacillus and Pseudomonas strains have been reported widely11.
The present investigation was undertaken to study induction of defense related enzymes against bacterial wilt caused by R. solanacearum to control the disease in tomato plants. The main objectives of this study was to evaluate the induction of various defense related genes encoding proteins implicated in strengthening of plant cell walls by biocontrol agents in response to infection by R. solanacearum.
MATERIALS AND METHODS
The whole study was carried out at Bacteriology Department, Division of Plant Pathology, Indian Agricultural Research Institute, Pusa, New Delhi, India during the period of July, 2015 to April, 2016.
Bacterial culture and Ralstonia solanacearum UTT-25: The Bacillus amyloliquefaciens DSBA-11 (Accession No. KF850150) and Ralstonia solanacearum UTT-25 (Accession No. KM921781) was obtained from Plant Bacteriology Laboratory Division of Plant Pathology, ICAR- IARI New Delhi, India. They were cultured in Luria Bertani (LB) agar and CPG media and stored in 50% glycerol at -80°C for further use.
Bacterial cultures and tomato seedlings: Bacillus amyloliquefaciens DSBA-11 was cultured in sterile LB agar plate at 28±1°C for 48 h. The cells were diluted with sterile distilled water upto 108 CFU mL–1. Ralstonia solanacearum UTT-25 was grown on CPG agar medium at 28±1°C for 48 h and the grown bacterial cells were harvested by sterile distilled water maintained the inoculums load 108 CFU mL–1. Tomato seeds of both cvs. Pusa Ruby (susceptible) and Arka Abha (moderately resistant) were grown in seed starter trays of 96 wells having the mixture of peat mass, vermiculli and sand in the ratio of 2:1:1 under controlled condition in Phytotron at 28±2°C, watered daily. One month old tomato seedlings were transplanted in 6 inch autoclave pot having 1 kg of soil mixture. Five days after transplanting, the plants were treated with B. amyloliquefaciens DSBA-11 and R. Solanacearum UTT-25 simultaneously. One set of control inoculated only with R. solanacearum UTT-25 was also maintained with three replications. The 6 treatments were included as T-1 and T-4: Arka Abha and Pusa Ruby were treated with water, T-2 and T-5 both cultivars of tomato Pusa Ruby and Arka Abha treated with B. amyloliquefaciens DSBA-11and R. solanacearum UTT-25, T-3 and T-6 the plants of Pusa Ruby and Arka Abha were treated with only R. solanacearum UTT-25. 25 mL of each B. amyloliquefaciens and R. solanacearum were inoculated at root zone of plants. At the time of harvesting, survived plants were sampled to get wilt intensity, fresh and dry weight. The wilt intensity was investigated at 5 days intervals upto 60 days. The calculation was done the average percentage wilt for each treatment. Disease rating was recorded by using following scale: 1= No symptoms, 2= One leaf wilted, 3 = 2-3 leaves wilted, 4 = 4 or more leaves wilted, 5: Whole plant wilted (dead plant). Calculate the wilt incidence 30 days after inoculation, using the following equation12.
Wilt intensity (%) (I) = [Σ (ni×vi)÷(V×N)] 100
where, ni is the number of plants with respective disease rating, vi is the disease rating (1, 2, 3, 4 or 5), V is the highest disease rating and N is the number of plants observed. Biocontrol efficacy was calculated as described by Guo et al.5. Growth promoting efficacy was calculated based on dry weight of root and shoot of plant as described by Singh et al.4.
Prepration of enzyme extracts: About 0.5 g leaf tissue of tomato cvs. Pusa Ruby and Arka Abha were collected from each treatments for enzyme extraction at intervals of 0, 24, 48, 72 and 96 h. Leaf tissue was homogenized in 0.1 M sodium phosphate buffer (20 mL), β mercaptoethanol (0.2 mL) and 1.0 g of insoluble polyvinyl pyrrolidone (PVPP) and centrifuged at 12000 rpm for 30 min at 4°C. The supernatant was measured and as per the ammonium sulphate concentration. After 1 h supernatant was centrifuged at 12000 rpm for 30 min at 4°C and pellets were resuspended in extraction buffer (3 mL), for the assay of the scavenging enzyme viz. Super oxide dismutase (SOD), catalase (CAT), peroxidase (POD), polyphenol oxidase (PPO) and phenyl alanine ammonia lyase (PAL) and photosynthetic pigments.
Determination of photosynthetic pigments: For photosynthetic pigments, Chlorophyll A, Chlorophyll B and β-carotene were estimated as described by Wintermans et al.13. About 0.5 g fresh leaves were grounded in a pestle and mortar and extracted by 15 mL of 80% acetone (1:100 w/v) and 0.5 g calcium carbonate. The mixture was filtered through a glass funnel and the residue was washed with a small volume of acetone and completed to 25 mL. The optical density (OD) of a constant volume of filtrate was measured at a wave length of 662, 644 and 440 nm for chlorophyll A, chlorophyll B and carotene, respectively.
The following equations were used:
Chl. A = 9.784 E.662-0.99 E.644 = mg L–1
Chl. B = 21.426 E.644-4.65 E.662 = mg L–1
Carotene = 4.695 E.440 - 0.268 (Chl.A- Chl.B) = mg L–1
where, E is the optical density at the wave length indicated
Estimation of PPO, POD and CAT activity: The activity of polyphenol oxidase, peroxidase and catalase were measured according to method described by Kar and Mishra14. For PPO activity, 2.0 mL of potassium phosphate buffer, 1.0 mL of pyragallol with 0.1 mL of enzyme extract were mixed together. For POD above described mixture and 1.0 mL of H2O2 were mixed and incubated at room temperature for 3 min and then immediately OD was taken at 480 and 420 nm for PPO and POD, respectively. One unit of enzyme activity was described by the change in absorbance of 0.1 min–1 and calculation of specific activity was done by using the equation.
(A×3/0.1)/3 = Specific activity min–1 mg–1 protein
where, A is the difference of absorbance of sample and the control (OD S-OD C).
For CAT activity, 1.0 mL of hydrogen peroxide and distilled water, 0.5 mL of enzyme extract with 2.0 mL of phosphate buffer were mixed together and OD was taken at 240 nm by using UV-VIS Spectrophotometer, Hitachi (U-2900). Absorbance was recorded at every 60 seconds against corresponding control of each sample. Specific activity calculation was done by using the formula
(A×3)/0.5) = Specific activity min–1 mg–1 protein in enzyme extract
where, A is the specific activity.
Estimation of SOD activity: Superoxide dismutase (SOD) activity was estimated using a nitro-blue tetrazolium (NBT) method as described by Beauchamp and Fridovich15. The 3.0 mL reaction mixture contained 0.1 M phosphate buffer (pH 7.8), 13 mM methionine, 0.1 mM EDTA, 75.0 μM NBT, 100 μL enzyme extract and 2 μM of riboflavin was added at last. The reaction mixtures were put in incubator and shaker in light for 10 min. The reaction was stopped by switching off the light and the tubes were covered with black cloth. The SOD activity was measured by optical density OD560. The OD value of reaction solution, the reduction of NBT by 50% was considered as 1 unit for enzyme activities.
Calculation of specific activity of SOD was done by using the equation:
(100-(OD S/OD LC) 100)/50 = x
where, x/mg protein in enzyme extract = Specific activity in units, 1 unit is defined as the 50% reduction in the blue color formed by NBT/30 min mg–1 protein, OD S is the OD T-OD C, OD T is the absorbance of sample, OD C is the absorbance of dark control and OD LS is the absorbance of light control.
Detection of PAL activity: The PAL activity of tomato leaf sample was assayed as described by Sadasivam and Manickam16. One unit of the enzyme was defined as increase in absorbance of 1 unit min–1. The OD290 was determined per 15 min until it kept steady and one unit enzyme was defined by the change in absorbance of 0.1 h–1.
Statistical analysis: The One-way analysis of variance for biocontrol efficiency and yield of tomato was made by the SAS general linear model (GLM) procedure (SAS Institute, Version 6, Cary NC). Mean comparison was conducted by a least significant difference (LSD) test (p = 0.05). Standard error and a LSD result were recorded. The values of enzyme activities were expressed as means of three measurements (±SE) from three plants having the same treatment.
RESULTS
Biocontrol efficacy and PGPR activity of B. amyloliquefaciens DSBA-11: Bacillus amyloliquefaciens DSBA-11inhibited the occurrence of bacterial wilt under green house conditions. Table 1 shows the biocontrol efficacy of DSBA-11 against bacterial wilt and growth promotion on tomato in the greenhouse was found better. Biocontrol efficacy of B. amyloliquefaciens DSBA-11 was 76.80% in Arka Abha and 73.97% in Pusa Ruby were recorded after 30 days of inoculation of the pathogen. Disease intensity was 21.64% in Pusa Ruby and 14.98% in Arka Abha treated by DSBA-11 while 83.15% in Pusa Ruby and 64.97% in Arka Abha treated only with R. solanacearum. Bacillus amyloliquefaciens can suppress the growth of R. solanacearum was also significantly suppressed by treating with the bioagents B. amyloliquefaciense DSBA-11. Thus the suppression of wilt disease in tomato treated with B. amyloliquefaciens (DSBA-11) may be due to the activity of secondary metabolites compounds produced by B. amyloliquefaciens cells in soil and by development of resistance in plants. The biomass of the average fresh weight of survived tomato plants treated by DSBA-11 was recorded higher in both tomato cultivars Pusa Ruby and Arka Abha with GPE 120.75 and 126.38%, respectively as compared to control treated with R. solanacearum (Table 1).
DSBA-11 induces the production of defense resistance related enzymes in tomato leaves: The bioagent B. amyloliquefaciens DSBA-11treated plants significantly increased gradually SOD production in both tomato cvs. Pusa Ruby and Arka Abha as compared to inoculated only with R. solanacearum.
| Table 1: | Reduction of bacterial wilt disease intensity and enhancement of biomass of tomato plants treated with antagonistic Bacillus amyloliquefaciens DSBA-11 |
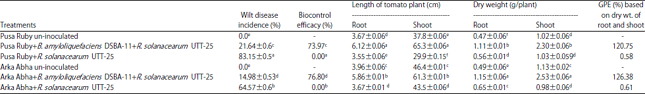 | |
| Means±SE (standard error) followed by the same letter do not differ significantly according to Duncan’s multiple range test at p = 0.05. Scheffe post hoc test. Means sharing different alphabetical (a, b, c, d, e) superscripts in a column significantly different (p<0.05) | |
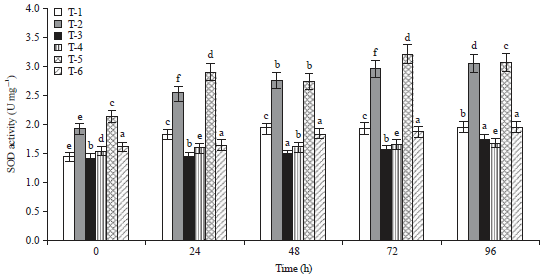 | |
| Fig. 1: | Total activity of SOD 0-96 h after inoculation in tomato |
Means±SE (standard error) followed by the same letter do not differ significantly according to Duncan’s multiple range test at p = 0.05. Scheffe post hoc test means sharing different alphabetical (a, b, c, d, e) superscripts in a column significantly different (p<0.05) | |
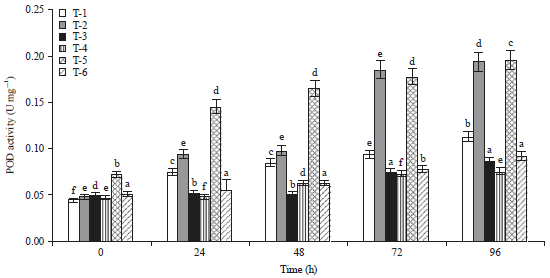 | |
| Fig. 2: | Total activity of POD 0-96 h after inoculation in tomato |
Means±SE (standard error) followed by the same letter do not differ significantly according to Duncan’s multiple range test at p = 0.05. Scheffe post hoc test means sharing different alphabetical (a, b, c, d, e) superscripts in a column significantly different (p<0.05) | |
It was noted that the highest amount of SOD activity was recorded in Arka Abha (3.08 U g–1) treated with bioagent followed by Pusa Ruby (3.07 U g–1) after 96 h of inoculation (Fig. 1). It is very interesting to observe that SOD activity was higher level even after 4 days of the treatments. As a major defensive enzyme SOD showed notably high activity. The rapid increase of POD activity was observed during first 2 days after bioagent treatments in both the cultivars. However, POD activity was found minimum (0.196 U g–1) in Arka Abha treated with bioagent as compared to Pusa Ruby (0.194 U g–1) (Fig. 2). This is probably because of the production of excess H2O2 by increasing POD in DSBA-11 treated plants. The highest PPO activity was also found in Arka Abha (0.210 U g–1) treated with biocontrol agent and in Pusa Ruby (0.129 U g–1) after 48 h of the pathogen inoculation was recorded after that it was decreased in all the treatments (Fig. 3). Additionally, it is also found that the activity of PPO was also stimulated by DSBA-11 treatment. The PAL activity in tomato cultivars was continuously increased in biocontrol agent treated plants upto 96 h. Figure 4 revealed that maximum production of PAL production was observed in bioagent treated plants in Pusa Ruby (749.3 U g–1), followed by Arka Abha (735.6 U g–1) after 96 h of pathogen inoculation. The PAL activity is one of the most important ISR enzymes which induce resistance in the plants against disease.
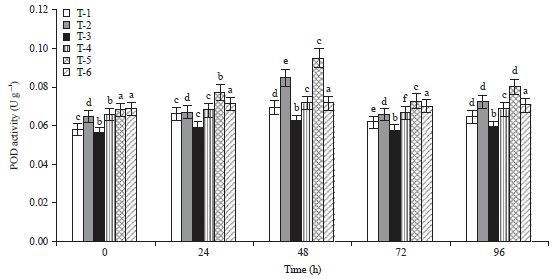 | |
| Fig. 3: | Total activity of PPO 0-96 h after inoculation in tomato |
Means±SE (standard error) followed by the same letter do not differ significantly according to Duncan’s multiple range test at p = 0.05. Scheffe post hoc test means sharing different alphabetical (a, b, c, d, e) superscripts in a column significantly different (p<0.05) | |
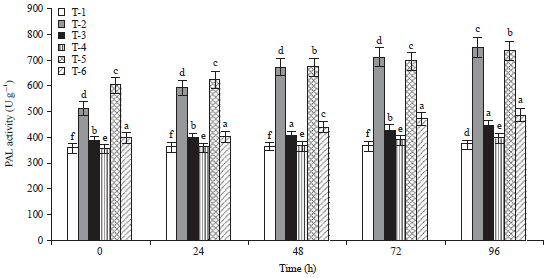 | |
| Fig. 4: | Total activity of PAL 0-96 h after inoculation in tomato |
Means±SE (standard error). Same letter do not differ significantly according to Duncan’s multiple range test at p = 0.05. Different alphabetical (a, b, c, d, e) superscripts in a column significantly different (p<0.05) | |
It was shown to catalyze the deamination of L-phenylalanine to produce cinnamic acid.
The H2O2 contents of tomato treated with B. amyloliquefaciens DSBA-11 were increased at the top level at 96 h after pathogen inoculation in both the cultivars Pusa Ruby and Arka Abha. Catalase contents of tomato treated with DSBA-11 were 7.22 U g–1 in Pusa Ruby and 7.83 U g–1 in Arka Abha at 96 h after pathogen inoculation (Fig. 5). In the present study revealed enhanced activities of defense related enzyme POP, POD, PAL and accumulation SOD and CAT in tomato plants treated with bio-control agents and challenged with R. solanacearum.
Chlorophyll content: Table 2 revealed that significant variations in chlorophyll content between healthy and non inoculated plants of cv. Pusa Ruby (susceptible) and cv. Arka Abha (moderately resistant) was recorded. However, both the cultivars treated with the R. solanacearum UTT-25 and B. amyloliquefaciens DSBA-11showed variations in chlorophyll content such as chlorophyll A, chlorophyll B and the carotene. While the DSBA-11 treated plants showed higher the all chlorophyll content as compared to only R. solanacearum treated plants. In summary B. amyloliquefaciens (DSBA-11) had good efficiency in bacterial wilt disease suppression and biomass increase under green house conditions.
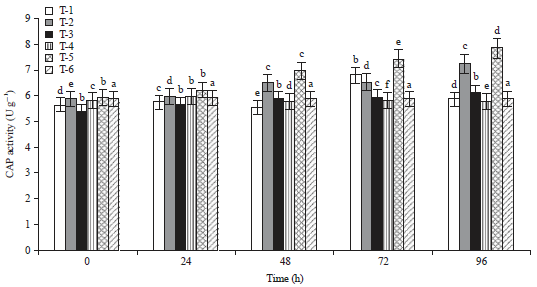 | |
| Fig. 5: | Total activity of CAT 0-96 h after inoculation in tomato |
Means±SE (standard error) followed by the same letter do not differ significantly according to Duncan’s multiple range test at p = 0.05. Scheffe post hoc test Means sharing different alphabetical (a, b, c, d, e) superscripts in a column significantly different (p<0.05) | |
DISCUSSION
In this study it was isolated a new member of bio control agenet of B. amyloliquefaciens DSBA-11which suppressed R. solanacearum causing bacterial wilt disese. Interestingly, it was found evidences that DSBA-11 could induce different ISR related enzyme. Tomato seedlings of both the cultivars cv. Pusa Ruby and Arka Abha produce significantly high amount of SOD, PPO, POD, CAT and PAL after treated with DSBA-11. The main mechanisms of bioconrol agents include the production of antibiotics, competition, plant growth promotion and induction of SAR and ISR17. Generally it was believed that beneficial bacteria can make the plant more tolerant to pathogen by stimulating ISR18. In this study, the activities of resistance related enzymes including SOD, POD, PPO, PAL and CAL considerably increased after treated by DSBA-11. It is proved that H2O2 appears to be a key element involved in disease resistance to pathogen1. Yamasaki et al.19 reported that biocontrol agent induced production of SOD against pathogen interaction of plants. However, the excess production of H2O2 will lead to the production of POD that scavenges H2O2. Additionally, author also found that the activity of PPO was also stimulated by DSBA-11 treatment. The PPO function as catalyzing the oxygen dependent oxidation of phenols to quinines and are assumed to be involved in plant defense against pests and pathogen20. PAL was also enhanced during DSBA-11 treatment. It was shown to catalyze the deamination of L- phenylalanine to produce cinnamic acid. Present results from green house experiments showed that the wilt incidence against bacterial wilt of tomato was as high 83.15% in Pusa Ruby while 64.57% in Arka Abha as compared to B. amyloliquefaciens (DSBA-11) treated plants which were 14.98% in Arka Abha and 21.64% in Pusa Ruby. The yield and biomass increased by B. amyloliquefaciens (DSBA-11) were plenty (Table 1). Jetiyanon et al.21 reported that three strains B. pumilus SE34 and SE49 and Pseudomonas fluorescens 89b-61 could induce systemic resistance on cucumber for colletotrichum orbiculare. B. pumilus SE34 and SE49 induced HR response in cucumber while P. fluorescens 89B-61 did not. Induced systemic resistance (ISR) of plants against pathogens is a world wide spread phenomenon. Interaction between plants and pathogens can lead either to a successful infection (compatible response) or resistance (incompatible response)22. The action of ISR is based on the defense mechanism that is activated by inducing agents23. In the present study revealed enhanced activities of defense related enzyme POP, POD, PAL and accumulation SOD and CAT in tomato plants treated with bio-control agents and challenged with R. solanacearum. Bacillus amyloliquefaciense can suppress the growth of R. solanacearum as it was also significantly suppresssed by treating with the bioagents B. amyloliquefaciense DSBA-11. Thus the suppression of wilt disease in tomato treated with B. amyloliquefaciens (DSBA-11) may be due to the activity of secondary metabolites compounds produced by B. amyloliquefaciens cells in soil. Infection by R. solanacearum is one of the major stress stimuli that plants often encode24. In summary B. amyloliquefaciens (DSBA-11) had good efficiency in bacterial wilt disease suppression and yield increase under green house conditions. Induced systemic resistance (ISR) on tomato by enriching the activity of oxidant and anti oxidant enzymes related to plant defense response at different stages.
| Table 2: | Effect of artificial inoculation with R. solanacearum and B. amyloliquefaciens on chlorophyllA, chlorophyll B and carotene contents in the two tomato cultivars Pusa Ruby and Arka Abha after inoculation under greenhouse conditions |
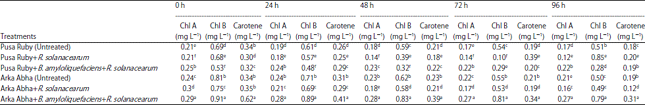 | |
Means±SE (standard error) followed by the same letter do not differ significantly according to Duncan’s multiple range test at p = 0.05. Scheffe post hoc test. Means sharing different alphabetical (a, b, c, d, e) superscripts in a column significantly different (p<0.05) | |
CONCLUSION
The present investigation shows the induction of systemic resistance by B. amyloliquefaciens against R. solanacearum in tomato screening for defense related enzymes for bacterial wilt disease control. Finally, peroxidase (POD), polyphenol oxidase (PPO), superoxide dismutase (SOD), phenylalanine ammonia lyase (PAL) and catalase (CAT) enzymes activities can be used as biochemical markers to reveal the resistance or susceptibility nature of tomato cultivars against bacterial wilt disease of tomato caused by R. solanacaerum.
SIGNIFICANCE STATEMENTS
The present study discovers the induction of systemic resistance by B. amyloliquefaciens against R. solanacearum in tomato screening for defense related enzymes for bacterial wilt disease control. And screening of different ISR enzyme like peroxidase (POD), polyphenol oxidase (PPO), superoxide dismutase (SOD), phenylalanine ammonia lyase (PAL) and catalase (CAT) activities in susceptibility and resistance tomato cultivars Pusa Ruby and Arka Abha. These enzymes activities can be used as biochemical markers to reveal the resistance or susceptibility nature of tomato cultivars against bacterial wilt disease of tomato caused by R. solanacaerum.
ACKNOWLEDGMENTS
The authors are thankful to the Indian Council of Agricultural Research, New Delhi, to giving assistance under outreach project on Phytofura for conducting various experiments. The authors are also thankful to the Head, Division of Plant Pathology, Indian Agricultural Research Institute, New Delhi, for help throughout experimentation.
REFERENCES
- Singh, D., S. Sinha, D.K. Yadav, J.P. Sharma and D.K. Srivastava et al., 2010. Characterization of biovar/races of Ralstonia solanacearum, the incitant of bacterial wilt in solanaceous crops. Indian Phytopath., 63: 261-265.
Direct Link - Singh, D., D.K. Yadav, G. Chaudhary, V.S. Rana and R.K. Sharma, 2016. Potential of bacillus amyloliquefaciens for biocontrol of bacterial wilt of tomato incited by Ralstonia solanacearum. J. Plant Pathol. Microbiol., Vol. 7.
CrossRefDirect Link - Guo, J., Y. Guo, L. Zhang, H. Qi and Z.D. Fang, 2001. Screening for biocontrol agents against Ralstonia solanacearum. Chin. J. Biol. Control, 17: 101-106.
Direct Link - El-Abyad, M.S., M.A. El-Sayed, A.R. El-Shanshoury and N.H. El-Batanony, 1996. Effect of culture conditions on the antimicrobial activities of UV-mutants of Streptomyces corchorusii and S. spiroverticillatus against bean and banana wilt pathogens. Microbiol. Res., 151: 201-211.
CrossRefDirect Link - Chen, X.H., A. Koumoutsi, R. Scholz, K. Schneider and J. Vater et al., 2009. Genome analysis of Bacillus amyloliquefaciens FZB42 reveals its potential for biocontrol of plant pathogens. J. Biotechnol., 140: 27-37.
CrossRefDirect Link - Finger, A., 1994. In‐vitro studies on the effect of polyphenol oxidase and peroxidase on the formation of polyphenolic black tea constituents. J. Sci. Food Agric., 66: 293-305.
CrossRefDirect Link - Soares, A.G., L.C. Trugo, N. Botrel and L.F. da Silva Souza, 2005. Reduction of internal browning of pineapple fruit Ananas comusus L.) by preharvest soil application of potassium. Postharvest Biol. Technol., 35: 201-207.
CrossRefDirect Link - Li, S.M., G.G. Hua, H.X. Liu and J.H. Guo, 2008. Analysis of defence enzymes induced by antagonistic bacterium Bacillus subtilis strain AR12 towards Ralstonia solanacearum in tomato. Ann. Microbiol., 58: 573-578.
CrossRefDirect Link - Harish, S., M. Kavino, N. Kumar, D. Saravanakumar, K. Soorianathasundaram and R. Samiyappan, 2008. Biohardening with plant growth promoting rhizosphere and endophytic bacteria induces systemic resistance against Banana bunchy top virus. Applied Soil Ecol., 39: 187-200.
CrossRefDirect Link - Wintermans, J.F.G.M. and A.S. de Mots, 1965. Spectrophotometric characteristics of chlorophylls A and B and their phenophytins in ethanol. Biochim. Biophys. Acta (BBA)-Biophys. Incl. Photosynth., 109: 448-453.
CrossRefDirect Link - Kar, M. and D. Mishra, 1976. Catalase, peroxidase and polyphenoloxidase activities during rice leaf senescence. Plant Physiol., 57: 315-319.
CrossRefDirect Link - Beauchamp, C. and I. Fridovich, 1971. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem., 44: 276-287.
CrossRefPubMedDirect Link - Ramarathnam, R., W.G.D. Fernando and T. de Kievit, 2011. The role of antibiosis and induced systemic resistance, mediated by strains of Pseudomonas chlororaphis, Bacillus cereus and B. amyloliquefaciens, in controlling blackleg disease of canola. BioControl, 56: 225-235.
CrossRefDirect Link - Van der Ent, S., S.C.M. van Wees and C.M.J. Pieterse, 2009. Jasmonate signaling in plant interactions with resistance-inducing beneficial microbes. Phytochemistry, 70: 1581-1588.
CrossRefDirect Link - Yamasaki, H., N. Sakihama and N. Ikehara, 1997. Flavonoid-peroxidase reaction as a detoxification mechanism of plant cells against H2O2. Plant Physiol., 115: 1405-1412.
PubMedDirect Link - Li, L. and J.C. Steffens, 2002. Overexpression of polyphenol oxidase in transgenic tomato plants results in enhanced bacterial disease resistance. Planta, 215: 239-247.
CrossRefDirect Link - Heil, M. and R.M. Bostock, 2002. Induced systemic resistance (ISR) against pathogens in the context of induced plant defences. Ann. Bot., 89: 503-512.
CrossRefDirect Link - Maurhoffer, M., C. Hase, J.P. Matraux and G. Defago, 1994. Induction of systemic resistance of tobacco to tobacco necrosis virus by the root-colonizing Pseudomonas fluorescens strains CHAO: Influence of the gacA gene and pyoverdine production. Phytopathology, 84: 139-146.
Direct Link - Murthy, K.N., F. Uzma, Chitrashree and C. Srinivas, 2014. Induction of systemic resistance in tomato against Ralstonia solanacearum by Pseudomonas fluorescens. Am. J. Plant Sci., 5: 1799-1811.
CrossRefDirect Link