
Efendi
Laboratory of Genetics and Plant Breeding, Department of Agrotechnology, Faculty of Agriculture, University of Syiah Kuala, Jl. Tgk. Krueng Kalee No. 3, Darussalam, Banda Aceh, 23111, Indonesia
Elly Kesumawaty
Laboratory of Genetics and Plant Breeding, Department of Agrotechnology, Faculty of Agriculture, University of Syiah Kuala, Jl. Tgk. Krueng Kalee No. 3, Darussalam, Banda Aceh, 23111, Indonesia
Sabaruddin Zakaria
Laboratory of Genetics and Plant Breeding, Department of Agrotechnology, Faculty of Agriculture, University of Syiah Kuala, Jl. Tgk. Krueng Kalee No. 3, Darussalam, Banda Aceh, 23111, Indonesia
Bakhtiar and Syafruddin
Laboratory of Genetics and Plant Breeding, Department of Agrotechnology, Faculty of Agriculture, University of Syiah Kuala, Jl. Tgk. Krueng Kalee No. 3, Darussalam, Banda Aceh, 23111, Indonesia
International Journal of Agricultural Research
Year: 2015 | Volume: 10 | Issue: 2 | Page No.: 74-82







ABSTRACT
The purpose of this research was to evaluate the performances of morpho-agronomic characters of rice landraces in developing new varieties that are adaptable to climate change and suitable for the organic SRI method. Sixteen genotypes of the selected local rice were treated to evaluate the phenotypes under the organic cultivation method. The observed rice landraces showed a phenotypic variation in morpho-agronomic performances. Among them was the plant height, leaves length, leaves width, number of tillers, productive panicles, heading date, panicles length, filled grains per spike, empty grains per spike, 1000 grains weight and grain yield potential, which were clearly affected by the genotypes. Sigupai showed the tallest genotype, while Pade Mas showed the shortest genotype with the highest numbers of the filled grains and the grains yield potential. Rom Mokot had higher numbers of the filled grains and the grain yield potential compared to Sigupai. It was found that Ciherang had the shortest and the narrowest leaf with the lowest grain yield potential, while the genotypes of Salah Mayang Ru showed the tallest leaf and higher grain yield potential compared to Ciherang. The genotype of Sikuneng had the lowest number of tillers, but it had the highest productive panicles with moderate grain yield potential. The shortest period of flowering found was for the Ciherang variety. Meanwhile, the Sikuneng produced flower at the longest period for heading. Hence, the results of this discovery are expected to provide an improved sustainability of intensified systems for rice breeding of crop resistance to environmental stress in the future.
PDF Abstract XML References Citation
Received: February 05, 2015;
Accepted: June 25, 2015;
Published: October 14, 2015
How to cite this article
Efendi, Elly Kesumawaty, Sabaruddin Zakaria and Bakhtiar and Syafruddin, 2015. Morpho-Agronomic Performances of Rice (Oryza sativa L.) Landraces under Organic Cultivation of SRI Method. International Journal of Agricultural Research, 10: 74-82.
DOI: 10.3923/ijar.2015.74.82
URL: https://scialert.net/abstract/?doi=ijar.2015.74.82
DOI: 10.3923/ijar.2015.74.82
URL: https://scialert.net/abstract/?doi=ijar.2015.74.82
INTRODUCTION
After being affected by the giant tsunami waves following the 9.1 earthquake on December 26, 2004, many of the natural resources in the Aceh province of Indonesia were damaged, including the loss of germplasm of rice in the low land areas (Efendi, 2012). This province has since been affected by climate changes as well. Sipayung (2007) analyzed that the daily temperature has been increasing as the highest point in Aceh is located at the northwestern tip of the Sumatera island. The surface temperature increases to about 1.9°C during the period of a hundred year (1900-2000) observations. The IPCC (2007) revealed that the high temperature is becoming one of the significant abiotic stresses limiting plant growth and productivity, especially as the global temperature is probable to increase by 1.8-4.0°C at the end of the 21st century. Therefore, both the impacts of global warming and the loss of germplasm in this province become a more serious problem for ensuring food security in the future.
The disaster and climate change should be raised by adaptation and mitigation efforts to create a more resilient approach to obviate food crisis around the world. Resistance to abiotic stresses will become more important in the coming decades as farmers around the world have to cope with the effects of climate change and the growing frequency of extreme events. Breeding for adaptation to abiotic stress must be considered for the development of adaptive varieties in the future. Screening of rice collections for identification is very important to exploit the genetic diversity within rice germplasm. Manneh et al. (2011) revealed that the diversity of rice gene pool should be exploited for new donors of genes for climate change adaptation.
Therefore, this research attempted to study the morpho-agronomic performances of Acehness rice landraces under organic cultivtion system with the method of SRI (System of Rice Intensification) by using seeds collected from the post-tsunami affected areas in Aceh. Some research showed that morphological and agronomic traits, as well as reaction to abiotic stresses, that are known to be in the individual accessions increase the importance of the germplasm (Upadhyaya et al., 2008; Golam et al., 2011; El-Hendawy et al., 2012; Boriboonkaset et al., 2012; Hossain et al., 2013). Continuously, the morpho-agronomic performance leads to a more efficient use of germplasm in the collection, especially in developing the organic SRI method. There is a wide genetic variability available in rice among and between landraces leaving a wide scope for future crop improvement to be efficient (Rahman et al., 2009). Consequently, appropriate methods in programs of plant genetic improvement can reduce time and costs.
In this research, SRI technology under organic condition was developed. Cultivation of rice with SRI system showed a reduction in pests, diseases, grain shattering, unfilled grains and lodging, similar to the reduction of agricultural chemicals, water use and methane emissions of global warming (Efendi, 2011). The method of SRI enhances the growth and function of rice plants, root systems and elaborates the numbers and diversity of soil organisms that contribute to healthy plants and production (Randriamiharisoa et al., 2006; Uphoff, 2007, 2008; Kassam et al., 2011; Efendi et al., 2012). Thus, the purpose of this research was to evaluate the morpho-agronomic of local rice germplasm that could be used for developing new varieties that are adaptable to climate change and suitable for SRI method.
MATERIALS AND METHODS
Characterization of rice genotypes: A number of 112 samples of local rice varieties were collected during the year of 2008-2013 from fourteen districts in the Aceh province (Aceh Tamiang, Aceh Timur, Aceh Tengah, Gayo Luwes, Bener Meriah, Bireun, Pidie, Aceh Besar, Aceh Jaya, Aceh Barat, Nagan Raya, Aceh Barat Daya, Aceh Selatan and Simeulu). The samples of seeds were analyzed at the Laboratory of Seed Science and Technology, Faculty of Agriculture, Banda Aceh, Indonesia. Characterization of seed performances of each accession was described and catalogued to identify their accession traits. In addition, genotypic differences and similarities were also characterized and catalogued. Duplicate accessions were eliminated from the same variety or nearly identical variants of a variety. The duplicate accessions were removed ,when one variety has spread to several locations, resulting in morphological identical varieties and varieties with quite different names but essentially identical morpho-agronomic features.
Sixteen genotypes of the remaining samples were utilized in this experiment to evaluate their morpho-agronomic characters under organic cultivation of SRI practices. The local Acehnese rice genotypes were the focus of this research in selecting local rice germplasm based on their morpho-agronomic performances. The sixteen genotypes used in this research are: Acong, Aweuh, Bo Santeut, Bo 100, Pade Manggeng, Pade Bersilona, Pandrah, Pade Mas, Rom Mokot, Sigudang, Sanbei, Sigupai, Sipirok, Salah Mayang Ru, Sikuneng and Ciherang (the national variety used as the control genotype).
Crop establishment and evaluation of morpho-agronomic characters: The rice cultivation of field experiment was conducted by using the method of SRI in the district of Aceh Besar. The experiment was conducted at the Experimental Station, Faculty of Agriculture, University of Syiah Kuala, Banda Aceh, Indonesia from October, 2013 to May, 2014. Geographically, the experimental area is located at 5°57’N latitude and 95°37’E longitude at the elevation of above 9.11 m of the sea level. A common procedure of SRI method was followed by cultivating plants in pots. The rice seeds were germinated and sown into a tray containing natural sandy loam soil with 10% of compost. Seedlings of seven days old were uprooted from the nursery trays carefully. Then, the seedlings were transplanted into the well-puddled experimental pots filled with 6 kg of soil containing 9:1 of sandy loam and compost. No chemical fertilizers were applied during both in land preparation and crop establishment. The first weeding was done at 14 Days After Transplanting (DAT), followed by the second and third weeding at 14 days interval after the first weeding was done. Irrigation was applied by intermittent irrigation with alternate wetting and drying from transplanting to the maximum tillering stage. A thin layer of water (2 cm) was kept on the plots from the Panicle Initiation (PI) to the hard dough stage. The water was removed from the plots during the ripening stage. The crop of each plot was harvested separately on different dates, when 90% of the grains became golden yellow in color. The principal morpho-agronomic characters that were systematically recorded on the plant and pot basis at different growth stages were as follows, i.e., plant height, leaves length, leaves width, number tillers, productive panicles, heading date, panicles length, filled grains per spike, empty grains per spike, 1000 grains weight and grain yield potential.
Experimental design and statistical analysis: The experiment was laid out in a Randomized Complete Block Design (RCBD) with 3 replications. Rice genotypes were designed as non-factorial RCBD. Drought stress was applied in a common SRI method and a two week desiccation was treated in vegetative stage, 30 days after transplanting, as a waterless stress condition. Rainfall was protected by using transparent plastic roof. The data were statistically analyzed by ANOVA (analysis of variance) technique. The significance of treatment effect was determined by using the F test. The significant difference between two treatments means and the Least Significant Difference (LSD) was estimated at the 5% probability level.
RESULTS AND DISCUSSION
Agro-morphologic performances of rice genotypes: Analysis of variance showed significant differences among the rice landraces for traits related to plant height, leaves length, leaves width, number of tillers, productive panicles, heading date, panicles length, filled grains per spike, empty grains per spike, 1000 grains weight and grain yield potential. The morpho-agronomic performances of rice genotypes clearly referred to a high genetic variation of the local rice. The characterized genotypes might provide an improved crop resistance to environmental stress in the future.
Plant height, leaves length and leaves width: Plant height was significantly affected by the observed genotypes. This result showed that plant height was clearly different among the genotypes (Table 1).
| Table 1: | Average plant height, leaves length, leaves width, number of tillers and productive panicles on different rice land races cultivated in organic SRI practices |
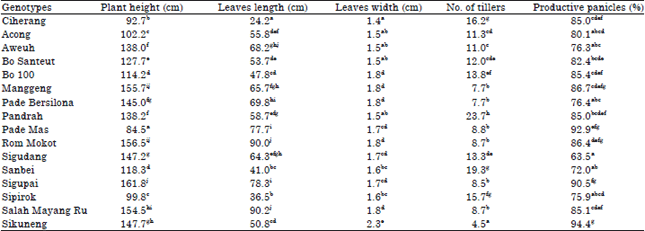 | |
| No. followed by the same letters in the same column are not significant at Duncan test (p<0.05) | |
The plant height ranged between 84.5-161.8 cm. Sigupai showed the tallest genotype, while, Pade Mas was the shortest genotype with the height was only 84.5 cm or 47.8% lower than the tallest genotypes. These results indicated that the identified genotypes of rice landraces have a high genetic variation. Some studies revealed that plant height is affected by a series of nodes and internodes of the genotypes. Internode elongation during vegetative growth is generally the culm which remains close to the ground and less develops. However, during reproductive growth, the uppermost internodes elongate to exert the panicle above the leaf sheaths (Smith and Dilday, 2003; El-Hendawy et al., 2012). Continuously, Hoshikawa (1989) discovered that the cotyledonary node is morphologically unique. The significant differences among the rice genotypes indicated the necessity to group them into clusters to identify the divergent groups (Bharadwaj et al., 2001; Yoshida and Horie, 2010).
Evaluated genotypes showed that leaves length and leaves width were significantly different among the genotypes (Table 1). Leaves length varied from 24.2-90.2 cm and leaves width ranged from 1.4-2.3 cm. It was found that Ciherang had the shortest and the narrowest leaf, while the genotypes of Salah Mayang Ru showed the tallest leaf (90.2 cm). Thus, the identified genotypes showed markedly different patterns of the leaf blade in their length and width. Farooq et al. (2010) revealed that leaf is an important morphological trait in rice. Leaf length and width are advantageous in enhancing the leaf area. Therefore, leaf characters were correlated with numbers of grains per panicle and grains weight per panicle. Accordingly, genetic improvement of trait to leaf performances is important for rice breeding (Tivet et al., 2001; Ohsumi et al., 2007; Wu et al., 2011).
Number of tillers, productive panicles and heading date: The number of tillers and productive panicles were significantly affected by the genotypes. The number of tillers ranged from 4.5-23.7 and productive panicles ranged from 63.5-94.4%. Table 1 shows that the highest number of tillers was found on the genotype of Pandrah (23.7) and was significantly different among the other genotypes. The genotype of Sikuneng, on the other hand, had the lowest number of tillers ,but with the highest productive panicles. Smith and Dilday (2003) explained that some cultivars have a maximum tiller number and are also observed to have a termination point for effective tillering. This condition of tiller number relates to the number of panicles at maturity. Yoshida (1981) revealed that some cultivars form tillers during the very early period, while others show delayed or sparse tillering. Performances of tillering are important to rice productivity because they affect the number of culms per square meter and grain yields per panicle (Jennings et al., 1989). A moderate number of vigorous early tillers are considered the most advantageous to produce a maximum yield (Counce et al., 1996). Thakur et al. (2011) proved that significant improvements were observed in number of tillers per hill under an aerobic condition. In other studies, Bian et al. (2013) analyzed that the number of final effective panicles was highly and significantly positive in correlation to the number of tillers. This suggests that distinct genetic systems might be responsible for the number of tillers which might feasibly improve the final yield. Yaqoob et al. (2012) also proved that grain yield also had a high correlation with the number of productive tillers. The results of heading observation showed a phenotypic variation in days to flowering as the plants responded to the organic SRI method. It was found that the days to the flowering of all genotypes were in different periods (average from 63.2-108.2 days). The shortest period of flowering was found for the Ciherang variety. Meanwhile, the Sikuneng produced the flowers at the longest period. The other varieties also showed significant difference of periods to flowering (Table 1). The days to flowering are related to panicle initiation during the reproductive growth of the rice plant. Blanco (1982) revealed that panicle initiation marks the onset of the reproductive phase. Additionally, Moldenhauer et al. (1994) explained that the first visible sign taking place is referred to as a thin green band and is briefly visible at the lower most internode prior to its elongation. During this period, environmental factors, such as; temperature extremes, drought, nutrient efficiencies, or toxicities, can reduce numbers of panicle branches and/or spikelets and reduce pollen viability. This directly affects the second and third yield components (Smith and Dilday, 2003). Guo et al. (2013) stated that the heading date and grain yield are key factors in determining the commercial potential of a rice variety.
Filled grains per spike, empty grains per spike and 1000-grains weight: The yield components were significantly influenced by the genotypes of rice landraces (Table 2). In this study, the highest percentage of filled grains per spike (86.7%) was observed from the genotype of Pade Mas. It was 67.5% higher compared to the lowest percentage of filled grains per spike (28.2%) from the genotype of Sipirok. Inconsistently, the lowest percentage of empty grains per spike (13.3%) was also observed from the genotype of Pade Mas.
| Table 2: | Average heading date, filled grains, empty grains, 1000 grain weight and yield grain potential on different genotypes of rice plants cultivated in organic SRI practices |
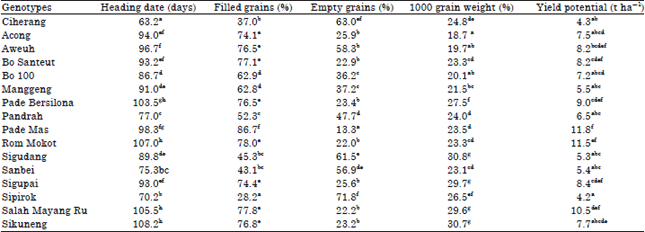 | |
| No. followed by the same letters in the same column are not significant at Duncan test (p<0.05) | |
It was also 78.90% lower compared to the higher percentage of empty grains per spike (71.8%) from the genotype of Sipirok. Tuyen and Prasad (2008) explained that the difference of yield parameters among rice genotypes varied among all morphological traits. Continuously, Liu et al. (2014) stated that the effect of the genotype on phenotype performances across environments become much more significant than that of the effects of environment and genotype-environment interactions. Desirable rice genotypes could then be selected by breeders based on the expected yield and adaptability to various environments.
The results of grains weight also showed a phenotype variation in grains weight of the observed rice landraces (Table 2). This study found that 1000 grains weight of the rice landraces was significantly different among the genotypes. The smallest size of grains was found for the accession of Acong (18.7 g–1 1000 grains), whereas, the Sigudang genotype showed the biggest size of grains (30.8 g–1 1000 grains). It was assumed that the grain size is also affected by the ripening process during the reproductive phase that was a genetic factor of the genotypes. Yoshida (1981) stated that grain growth during ripening is characterized by the increase in size and weight of kernels as starch and sugars are translocated from culms and leaves. Sarwar et al. (2012) found that there are significant differences regarding the values of morphological characters of rice grains. Grain size is the most important factor which influences the yield of rice quality. The grain has a strong correlation with 1000-grain weight. Maji and Saibu (2012) observed that a high coefficient of variation is found on 1000 grain weight of rice.
Grain yield potential: Table 2 shows that genotypes affected grain yield potential significantly. The highest result was obtained from the genotype of Pade Mas (11.8 t ha–1). In contrast, the lowest result was found from the genotype of Ciherang (4.3 t ha–1) it was 63.6% lower than the highest yield of Pade Mas. It was assumed that the Pade Mas as rice landrace had higher yield. Mishra and Slokhe (2011) explained that greater root length density and chlorophyll content of the flag leaf and the duration of grain filling affect the yield-contributing parameters in all trials. These relationships can significantly improve rice plant physiological efficiency and, hence, grain yield. Uphoff et al. (2011) observed that SRI practices make soil conditions more aerobic and promote greater root growth, as well as larger, more diverse communities of beneficial soil biota. These below-ground changes support more productive phenotypes above-ground for practically all rice genotypes (cultivars) tested so far. In another research, Thakur et al. (2011) found that grain yield increased and significant improvements were observed in the morphology of SRI plants in terms of root growth, plant/culm height, tiller number per hill, tiller perimeter, leaf size and number, leaf area index, specific leaf weight and open canopy structure. These phenotypes improvements of the SRI crop were accompanied by physiological changes: Greater xylem exudation rate, crop growth rate, mean leaf elongation rate and higher light interception by the canopy. So, the results of the rice landraces evaluation showed considerable promise as a material research with various good characters, for example, high yield potential, high levels of tolerance to stresses such as weeds, drought, acidity and blast for further evaluation. The progenies may provide an improved sustainability of intensified systems through durable crop resistance to biotic and abiotic stress.
CONCLUSIONS
The agro-morphology performances of rice landraces clearly referred to a high genetic variation of traits related to plant height, leaves length, leaves width, number of tillers, productive panicles, heading date, panicles length, filled grains per spike, empty grains per spike, 1000 grains weight and grain yield potential. Sigupai showed the tallest genotype, while Pade, Mas and Rom Mokot had higher numbers of the filled grains and the grains yield potential compared to Sigupai. Ciherang had the shortest and the narrowest leaf with the lowest grain yield potential, while Salah Mayang Ru showed the tallest leaf and had higher grain yield potential compared to Ciherang. The genotype of Sikuneng had the lowest number of tillers but it had the highest productive panicles with moderate grain yield potential.
ACKNOWLEDGMENTS
Thanks to the Directorate General of Higher Education, Ministry of Education and Culture of the Republic of Indonesia that has funded this research in accordance with the budget funds in 2012-2014 and to all parties, who have contributed to this research, especially to Hotna Riris Simanjuntak, Ridwan Anhar and Jefri, as technical assistants who helped planted cultivation and maintenance of rice plant.
REFERENCES
- Bian, J., H. He, H. Shi, C. Zhu and X. Peng et al., 2013. Dynamic QTL detection and analysis of tiller number before and after heading in japonica rice. Aust. J. Crop Sci., 7: 1189-1197.
Direct Link - Boriboonkaset, T., C. Theerawitaya, A. Pichakum, S. Cha-um, T. Takabe and C. Kirdmanee, 2012. Expression levels of some starch metabolism related genes in flag leaf of two contrasting rice genotypes exposed to salt stress. Aust. J. Crop Sci., 6: 1579-1586.
Direct Link - Counce, P.A., T.J. Siebenmorgen, M.A. Poag, G.E. Holloway, M.F. Kocher and R. Lu, 1996. Panicle emergence of tiller types and grain yield of tiller order for direct-seeded rice cultivars. Field Crops Res., 47: 235-242.
CrossRefDirect Link - Efendi, 2011. The system of rice intensification (SRI) as technology innovation to improve the productivity of rice (Oryza sativa L.) in post-tsunami affected-area of Aceh Province. Proceedings of the 6th Annual International Workshop and Expo on Sumatra Tsunami Disaster & Recovery in Conjunction with 4th South China Sea Tsunami Workshop, November 22-24, 2011, Syiah Kuala University, Banda Aceh.
- Efendi, 2012. Selection of Acehnese germplasm of rice (Oryza sativa L.) using SRI approach in the post-tsunami affected area of Aceh Province, Indonesia. Proceedings of the 2nd Annual International Conference Unsyiah and 8th IMT-GT UNINET Biosciences Conference, November 22-24, 2012, Syiah Kuala University, Banda Aceh.
- Efendi, Halimursyadah and H.R. Simanjuntak, 2012. The response of local Acehnese germplasm of rice to growth and production in aerobic condition. Agrista, 16: 114-121.
Direct Link - El-Hendawy, S., C. Sone, O. Ito and J.I. Sakagami, 2012. Differential growth response of rice genotypes based on quiescence mechanism under flash flooding stress. Aust. J. Crop Sci., 6: 1587-1597.
Direct Link - Farooq, M., N. Kobayashi, O. Ito, A. Wahid and R. Serraj, 2010. Broader leaves result in better performance of indica rice under drought stress. J. Plant Physiol., 167: 1066-1075.
CrossRefPubMedDirect Link - Golam, F., Y.H. Yin, A. Masitah, N. Afnierna, N.A. Majid, N. Khalid and M. Osman, 2011. Analysis of aroma and yield components of aromatic rice in Malaysian tropical environment. Aust. J. Crop Sci., 5: 1318-1324.
Direct Link - Hossain, M.A., M.R. Ismail, M.K. Uddin, M.Z. Islam and M. Ashrafuzzaman, 2013. Efficacy of ascorbate-glutathione cycle for scavenging H2O2 in two contrasting rice genotypes during salinity stress. Aust. J. Crop Sci., 7: 1801-1808.
Direct Link - Kassam, A., W. Stoop and N. Uphoff, 2011. Review of SRI modifications in rice crop and water management and research issues for making further improvements in agricultural and water productivity. Paddy Water Environ., 9: 163-180.
CrossRefDirect Link - Guo, L., Z.H. Zhang and J.Y. Zhuang, 2013. Quantitative trait loci for heading date and their relationship with genetic control of yield traits in rice (Oryza sativa). Rice Sci., 20: 1-12.
CrossRefDirect Link - Liu, C.G., X.Q. Zhou, D.G. Chen, L.J. Li, J.C. Li and Y.D. Chen, 2014. Natural variation of leaf thickness and its association to yield traits in indica rice. J. Integr. Agric., 13: 316-325.
CrossRefDirect Link - Maji, A.T. and A.A. Shaibu, 2012. Application of principal component analysis for rice germplasm characterization and evaluation. J. Plant Breed. Crop Sci., 4: 87-93.
Direct Link - Mishra, A. and V.M. Salokhe, 2011. Rice root growth and physiological responses to SRI water management and implications for crop productivity. Paddy Water Environ., 9: 41-52.
CrossRefDirect Link - Ohsumi, A., A. Hamasaki, H. Nakagawa, H. Yoshida, T. Shiraiwa and T. Horie, 2007. A model explaining genotypic and ontogenetic variation of leaf photosynthetic rate in rice (Oryza sativa) based on leaf nitrogen content and stomatal conductance. Ann. Bot., 99: 265-273.
CrossRefDirect Link - Rahman, M.S., M.R. Molla, M.S. Alam and L. Rahman, 2009. DNA fingerprinting of rice (Oryza sativa L.) cultivars using microsatellite markers. Aust. J. Crop Sci., 3: 122-128.
Direct Link - Sarwar, A.G., M.A. Ali and M.A. Karim, 2012. Correlation of grain characters in rice (Oryza sativa L.). J. Nat. Sci. Foundat. Sri Lanka, 26: 209-215.
Direct Link - Thakur, A.K., S. Rath, D.U. Patil and A. Kumar, 2011. Effects on rice plant morphology and physiology of water and associated management practices of the system of rice intensification and their implications for crop performance. Paddy Water Environ., 9: 13-24.
CrossRefDirect Link - Tivet, F., B.D.S. Pinheiro, M. de Raissac and M. Dingkuhn, 2001. Leaf blade dimensions of rice (Oryza sativa L. and Oryza glaberrima Steud.). Relationships between tillers and the main stem. Ann. Bot., 88: 507-511.
Direct Link - Upadhyaya, H.D., C.L.L. Gowda and D.V.S.S.R. Sastry, 2008. Plant genetic resources management: Collection, characterization, conservation and utilization. J. SAT Agric. Res., 6: 1-16.
Direct Link - Uphoff, N., A. Kassam and R. Harwood, 2011. SRI as a methodology for raising crop and water productivity: Productive adaptations in rice agronomy and irrigation water management. Paddy Water Environ., 9: 3-11.
CrossRefDirect Link - Wu, L.L., Z.L. Liu, J.M. Wang, C.Y. Zhou and K.M. Chen, 2011. Morphological, anatomical and physiological characteristics involved in development of the large culm trait in rice. Aust. J. Crop Sci., 5: 1356-1363.
Direct Link - Yaqoob, M., R. Anjum, M. Hussain and M.J. Shah, 2012. Genetic diversity analysis and character association in some Chinese hybrid rice under dry conditions. Pak. J. Agric. Res., 25: 249-256.
Direct Link - Yoshida, S., 1981. Fundamentals of Rice Crop Science. 1st Edn., International Rice Research Institute, Phillipines, ISBN: 971-104-052-2, Pages: 267.
Direct Link - Yoshida, H. and T. Horie, 2010. A model for simulating plant N accumulation, growth and yield of diverse rice genotypes grown under different soil and climatic conditions. Field Crops Res., 117: 122-130.
CrossRefDirect Link