
Dagnachew Lule
Bako Agricultural Research Center, P.O. Box 03, Bako, Ethiopia
Kassahun Tesfaye
Faculty of Life Science, Addis Abba University, Addis Abba, Ethiopia
Masresha Fetene
Faculty of Life Science, Addis Abba University, Addis Abba, Ethiopia
Santie de Villiers
Faculty of Life Science, Addis Abba University, Addis Abba, Ethiopia
International Journal of Agricultural Research
Year: 2012 | Volume: 7 | Issue: 6 | Page No.: 303-314







ABSTRACT
The gradual change in climatic conditions, particularly rainfall distribution in tropical and sub tropical regions of the world necessitate looking for productivity enhancement of stress tolerant crops such as finger millet as one option. Assessing genetic variation is a crucial for varietal development and genetic resource conservation. To this regard, a study was conducted at Gute and Arsi Negele (Ethiopia) during 2011 cropping season on one hundred and forty four finger millet landraces collected from different regions of Ethiopia, some introduced from Kenya, Eritrea, Zambia and Zimbabwe to evaluate the genetic diversity for quantitative traits at population level and eco-geographical regions of origin. The trend of quantitative trait diversity revealed that the highest genetic diversity were observed at the lowest level (among landrace populations) followed among the regions or countries of origin and least among altitude classes. This leads to suggest, taking more samples within a locality or population would be a better approach to capture the range of variation in finger millet population. Cluster analysis indicated that finger millet populations from neighboring regions of Ethiopia, neighboring African countries and proximity in altitude classes shared strong similarity. The similarity could be either due to fact that farmer’s selection criteria for a given traits might be similar particularly based on the adaptive role of traits for the environment, the primary seed source can be the same, or high tendency of seed exchange. Principal component analysis at populations level, geographical locations and agro-ecologies of origin indicated that grain yield per plant, thousand grain weight, days to heading, days to maturity, lodging index and biomass weight per plant were the most important traits contributing for the overall variability implying that breeding effort on those traits can meet the targeted objective.
PDF Abstract XML References Citation
Received: April 27, 2012;
Accepted: July 03, 2012;
Published: July 20, 2012
How to cite this article
Dagnachew Lule, Kassahun Tesfaye, Masresha Fetene and Santie de Villiers, 2012. Multivariate Analysis for Quantitative Traits in Finger Millet (Eleusine coracana subsp. coracana) Population Collected from Eastern and Southeastern Africa: Detection for Patterns of Genetic Diversity. International Journal of Agricultural Research, 7: 303-314.
DOI: 10.3923/ijar.2012.303.314
URL: https://scialert.net/abstract/?doi=ijar.2012.303.314
DOI: 10.3923/ijar.2012.303.314
URL: https://scialert.net/abstract/?doi=ijar.2012.303.314
INTRODUCTION
Finger millet (Eleusine coracana L.) is one of the most important cereal crops in large areas of the developing world especially Africa and India (Ekwamu, 1991). It is believed to be an allotetraploid (Babu et al., 2007). Finger millet can produce better yield than other crops under multiple stress such as drought, soil acidity and land marginality (Barbeau and Hilu, 1993; Upadhyaya et al., 2006). Moreover, it has high nutritional value and excellent storage qualities (Dida et al., 2007). In the semiarid tropics of Eastern Africa, it is the major staple food for millions of resource poor peoples and plays an important role in the dietary habits and economy of subsistence farmers.
Improvement in any crop usually involves exploiting the genetic variability in specific traits. Genetic variability is expressed as the genetic differences between species, sub species, varieties, population or individuals (Jarvis et al., 2000). Species with greater genetic diversity are more likely to be able to evolve in response to a changing environment than those with low genetic diversity. Populations that lack genetic diversity may experience low fertility and high mortality among offspring even in the environments that are fairly stable (Hunter, 1996). Studying the genetic variation of a crop species is essential for effective utilization of germplasm in plant breeding programs, devising appropriate sampling procedures for germplasm collection and conservation, obtaining some collections for efficient germplasm management and explicating the taxonomy, evolution and origin of crop species (Bekele, 1983, 1985; Demissie and Bjonstrand, 1996; Keneni et al., 2007; Varshney et al., 2007; Lule et al., 2011).
Compared to other cereal crops such as wheat, maize, barley, rice and sorghum, comprehensive study on finger millet diversity using morphological or molecular markers are limited (Bezaweletaw et al., 2006; Upadhyaya et al., 2006). This study was initiated, therefore, to assess the patterns of genetic variability, identify major traits contributing for variations among population, regions and altitudes of origin for further utilization in the breeding programs and to design appropriate sampling procedures for germplasm collection and conservation.
MATERIALS AND METHODS
A total of 144 finger millet landraces collected from different finger millet producing regions of Ethiopia (Tigray, Amhara, Oromia, Benishangul-Gumuz and Southern Nation- Nationalities and peoples region) and some introduced from Kenya, Eritrea, Zambia and Zimbabwe were planted with six improved varieties at Arsi Negele (altitude of 1947 m.a.s.l., N: 07019" and E: 38°39") and Gute (altitude of 1906 m.a.s.l. N: 09°00" and E: 36°38") in 2011 main cropping season (Table 1).
| Table 1: | Regional and altitudinal distribution of finger millet landraces used for genetic diversity study |
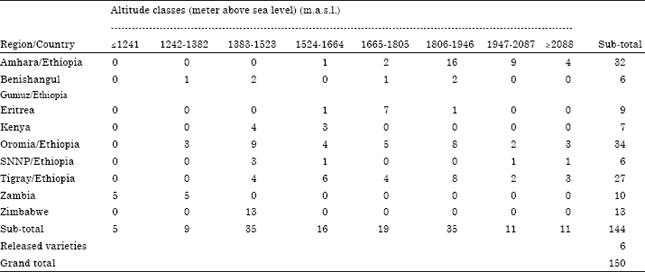 | |
They were used to group finger millet landraces with relative resemblance of agro-climatic origin using design was RCBD with two replication and plot size was single row of 2 m long and 50 cm between row spacing. Spacing between plants within row was adjusted to 10 cm. Each block was folded in to two. Improved finger millet varieties such as Boneya, Wama, Gute, Bereda, Tadesse and Padet were used. The first four were released from Bako Agricultural research center (Ethiopia) and the last two were released from Melkasa Agricultural research center (Ethiopia). Eight altitude classes the formula: K = 1+3.32 log10 n and W= (L-S)/K (Agrawal, 1996), where K= number of class interval, W = width of class interval, L = the largest value, S = the smallest value and n = sample size (in this case the number of landraces used in the study) (Table 1).
Data collection: Data were recorded for 17 quantitative morphological traits such as days to 50% heading, days to maturity, plant height (cm), culm length (cm), culm diameter (cm), finger length (cm), number of finger per main ear, total number of tiller, productive tiller number, number of grain per spikelet, number of culm branch per main plant, lodging index, ear weight (g), biomass weight per plant (g), harvest index (%), thousand grain weight and grain yield per plant (g) following finger millet descriptor (IBPGR, 1985).
Data analysis
Descriptive statistics: The major descriptive statistics such as mean, range, standard deviation and coefficient of variation for each traits were computed using MINITAB14 software (MINITAB, 2003).
Cluster analysis: Hierarchal clustering of average linkage method with squared Euclidian distance was performed using MINITAB14 software (MINITAB, 2003). Data of all quantitative traits were standardized to a mean of zero and a variance of one before clustering to avoid bias that arise due to differences in measurement scales. The distances between clusters were assessed using Mahalanobis’s distance (D2) for the standardized data. The concept of D2 statistics for measuring the divergence between the two population or groups of population was introduced by Mahalanobis (1936) and gives a result based on the magnitude of divergence and is independent of size of samples.
Principal component analysis: Principal component analysis for 17 standardized quantitative traits were computed by using Agrobase (2000) computer software at population level, regions of collection and altitude classes to identify the most important traits contributing to the total variations observed among the populations, countries, regions and altitude zones. As suggested by Johnson and Wichern (1988), principal components with eigenvalues greater than one was considered.
RESULTS
Patterns of genetic variation: Wider ranges of variations were observed among finger millet population for all traits (Table 2). Among populations, days to maturity ranges from 143 days for accession 230103 of Eritrea collected from altitude of 1700 m.a.s.l. to 167 days for Acc. BKFM0018 of Ethiopia (Oromia region) collected from altitude of 1667 m.a.s.l. Likewise, plant height ranges from 41.13 cm for Acc.214991 of Zambia collected from 1330 m.a.s.l. to 103.35 cm for Acc. 215802 of Ethiopia (Amhara region) collected from 1950 m.a.s.l.
| Table 2: | Patterns of genetic variability for 17 quantitative traits among finger millet population, regions and altitudes of origin |
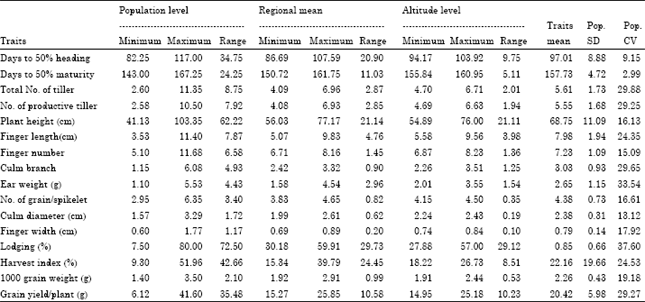 | |
For finger length, Acc. 229730 collected from Amhara region of Ethiopia attain maximum (11.4 cm), but Acc.203357 of Zimbabwe were the least (3.53 cm). Finger number per ear ranges from 5.1 for Acc. BKFM0028 of Ethiopia (Oromia region) to 11.68 for Acc.215802 of Ethiopia (Amhara region). For number of grains per spikelet, the highest (6.35) and the lowest (2.95) was observed for Acc. BKFM0042 and Acc. 216056, respectively, both from Oromia region of Ethiopia. Acc. 229724 of Benishangul Gumuz region (Ethiopia) attains the least thousand seed weight (1.4 g), but Acc. 203546 of Kenya was the highest (3.5 g). Grain yield per plant was highest (41.6 g) for Acc. 242132 of Amhara region (Ethiopia) collected from an altitude of 1910 m.a.s.l., but lowest (6.12 g) for Acc. 214991 of Zambia collected from 1330 m.a.s.l.
Among regions and countries of origin, the mean days to maturity, plant height, finger length and finger number were higher for populations from Ethiopia (B/Gumuz, Oromia, Amhara and Southern nations region, respectively). Collections from Eritrea were characterized as early maturing with smaller finger number per ear. Population from Zambia has shorter plant height and the least in grain yield per plant, but has higher number of grains per spikelet. Lowest number of grains per spikelet and thousand grain weight were recorded for B/Gumuz region (Ethiopia). The highest mean grain yield per plant and thousand grain weights were recorded for Kenyan’s (Table 2).
Among altitude classes, the least mean value for plant height, finger length, finger number per ear and grain yield per plant were observed in the range below 1241 m.a.s.l. but maximum for altitude class above 2088 m.a.s.l. (Table 2). In other words, the two extreme altitude classes were described for an extreme mean value of grain yield and yield related traits.
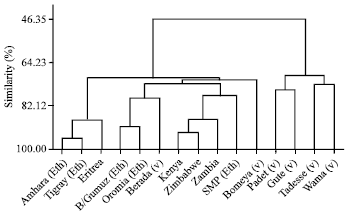
Cluster analysis: The cluster analysis of 144 finger millet landraces for the five African countries of origin showed that two clusters were formed at 50% similarity level (Fig. 1). Ethiopian and Eritrean populations grouped in the first cluster (left to right); Kenyan, Zambian and Zimbabwe’s populations grouped in the second cluster. At 75% similarity level, only Kenyan and Zimbabwe’s landraces share similarity for the 17 quantitative traits and the remaining 3 countries were separate or un-clustered.
 | |
| Fig. 1: | Similarities among 5 countries of origin for 144 finger millet landraces evaluated for 17 quantitative traits |
 | |
| Fig. 2: | The genetic relatedness of 144 F. Millet landraces for 17 quantitative traits among regions and countries of origin and 6 released varieties |
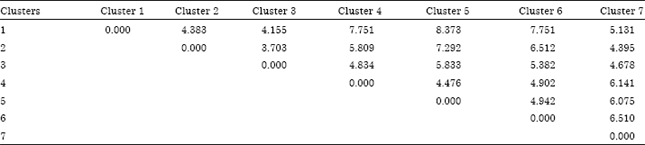
At 75% similarity level, populations from 5 major finger millet producing regions of Ethiopia, four African countries and six released varieties were grouped in to four clusters and three varieties remains solitary (Fig. 2). Populations collected from Tigray and Amhara regional state showed strong relatedness with Eritrean’s and hence fall in the first cluster. Landrace from Oromia and B/Gumuze regional state and Bereda variety were grouped in the second cluster; Kenyan, Zambian, Zimbabwe’s and Southern Nation Nationalities and Peoples region of Ethiopia were grouped in the third cluster. Released varieties such as Gute and Padet were clustered together in the fifth cluster, but Boneya, Wama and Tadesse remain solitary in the 4, 6 and 7th cluster, respectively. Mahalanobis’s genetic distance (D2) was relatively higher between first and the fifth cluster. The populations in first cluster were also showed wider genetic distance or dissimilarity to the fourth and the six clusters (Table 3).
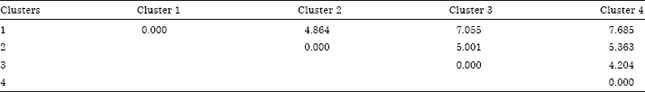
Altitudinally, three clusters and one solitary group were formed at 80% similarity level (Fig. 3). Proximity in altitude classes were clearly manifested in all the three clusters. Finger millet population from altitude level above 2088 remains solitary mainly due to its incomparable character such as the presence of highest plant height, finger length, finger number, culm branch, grain yield and harvest index. Relatively higher genetic distance (D2) was recorded between the two extreme altitude classes (Table 4).
| Table 3: | Mahalanobis’s distance (D2) between clusters for standardized data (inter-country and regional level) |
 | |
| Table 4: | Mahalanobis’s distance (D2) between clusters of altitude classes |
 | |
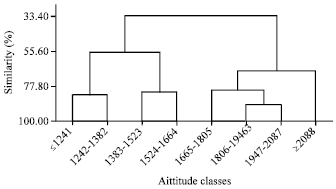 | |
| Fig. 3: | Dendrogram showing similarities among altitudes of collection for 144 F. Millet landrace evaluated for 17 quantitative traits |
Principal component analysis
Population based: The first five principal components having eigenvalue greater than one were extracted from the mean of 17 normalized quantitative traits of 144 finger millet populations (Table 5). A variance of 26.2, 15.3, 11.9, 9.0 and 6.2% were extracted from the first to the fifth components, respectively. 83.5% variation was extracted from the first eight principal components. Agronomic and phenotypic characters such as thousand grain weight, ear weight, finger length, productive tiller number, total tiller number and lodging index were the major contributors for the variation observed in the first principal component. The variation in the second principal component were mainly due to days to heading, days to maturity, biomass weight per plant, plant height and grain yield per plant. Likewise, finger number per main ear, culm diameter, harvest index, plant height and culm branch were among the major contributors to the variation in the third component. Total tiller number, productive tiller number and lodging index were the major contributor for the variation observed in the fourth component. The variability in the fifth component was attributed mainly due to number of grains per spikelet, biomass weight per plant and finger width.
| Table 5: | Principal component analysis for 17 quantitative traits of 144 finger millet landraces |
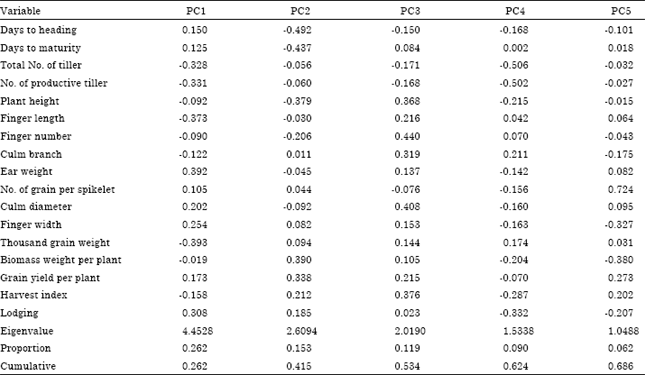 | |
| PC: Principle components | |
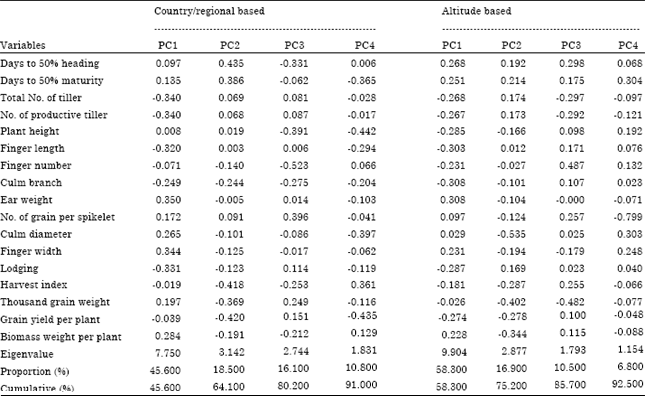
Geographical location based PCA: A variance of 45.6, 18.5, 16.1 and 10.8% were extracted from the first four principal components with eigenvalue greater than one, and 91.0% of the total variance was explained by these components (Table 6). Ear weight, finger width, lodging index, total tiller number, productive tiller number and finger length were the major contributors for the variation in the first principal component. Days to heading, harvest index, grain yield per plant and days to maturity for the second; finger number per main ear, number of grain per spikelet, plant height and days to heading for the third; plant height, culm diameter, grain yield per plant, harvest index and days to maturity for the fourth were the major traits contributing for the variation observed in the respective principal components (Table 6). The relative positions of regions of origin and improved varieties on the first two principal axis (Fig. 4) clearly reflected the hierarchical clustering affinities and the different geographical location of germplasm source and released varieties have been scattered in the entire four quadrants (Fig. 4).
Clinal/altitudinal based PCA: The total variance of 92.5% was observed for the first four principal components having eigenvalue greater than one (Table 6). 58.3% was attributed to the first component mainly due to ear weight, culm branch per plant and finger length as the major input. Culm diameter, thousand grain weight and biomass weight per plant was the major contributor for the variation observed in the second cluster. Finger number per main ear and thousand grain weights were the major input for the variation in the third component. Likewise, number of grain per spikelet, culm diameter and days to maturity for the fourth component. The eight altitude classes were distributed among four quadrants and their relative position on the principal axis partly confirm the cluster result of altitude zones (Fig. 5).
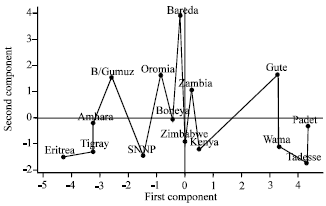 | |
| Fig. 4: | The relative position of countries/regions of origin and released varieties on the first two principal component |
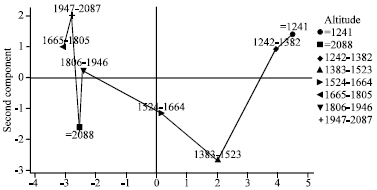 | |
| Fig. 5: | The relative position of eight altitude classes of origin on the first two principal component coordinates |
| Table 6: | Principal component analysis for 17 quantitative traits of finger millet based on geographical location and agro-ecologies of germplasm source |
 | |
| PC: Principle components | |
DISCUSSION
Patterns of diversity: The wider ranges of variation for several traits observed among finger millet populations, regions of origin and altitude classes are essential for effective collection, conservation and sustainable improvement of finger millet by combining the desirable traits together. The variation for days to heading and days to maturity offers great flexibility for developing improved varieties suitable for various agro-ecologies of countries or regions which have variable length of growing period and also to use in various cropping systems. It also guide breeder to develop a variety which escape late season drought by improving traits which correlate to days to maturity in the required direction. The variation in plant height, culm diameter, culm branch and tillering capacity indicates the possibility to combat lodging problem. Variation in number of finger per main ear, finger length, number of grains per spikelet, harvest index, thousand grain weight and grain yield per plant implies that the possibility to create a variety with higher grain yield and/or other biological yields.
Moreover, the trend of quantitative trait diversity in the present study showed that the highest genetic diversity were observed among landrace populations followed by among regions or countries of origin and least among altitude classes. This leads to suggest that taking more samples within a locality or population would be a better approach to capture the range of variation in finger millet population. In line with the present study, Bekele (1984) found that the total phenotypic variation for bread wheat was highest at the lowest level (within localities), followed by difference among populations within a region, and the least among regions. Besides, the extent of genetic variation in tef landraces is highest among accessions, followed by within regions and with altitudes but lower among regions and among altitudes (Lule et al., 2011). Contradicting results were reported for bread wheat, where diversity was higher among district than among population within district (Tadesse et al., 1991).
Cluster analysis: The result for cluster analysis at the level of countries, regions and altitude classes of origin indicated that neighboring regions, countries and proximity in altitude classes shared strong similarity. The similarity could be either due to fact that farmer’s selection criteria for a given traits might be similar particularly based on the adaptive role of traits for the environment, the primary seed source can be the same, or high tendency of seed exchange. Other possibilities could be inter and intra-country/regional migration and gene flow. Even if the geographical location of Kenya is at distant to Zambia and Zimbabwe, populations from those countries showed strong similarity. The most probable reasons can be, migration and gene flow was not limited by distance, the primary seed source for these countries can be the same, eco-geographic similarity for the adaptation of similar populations, the combination of those and other factors. In support of the present study, finger millet landraces collected from Kenya, Tanzania and Uganda were grouped together, but landraces from Ethiopia and Burundi were in a separate cluster each (Reddy et al., 2009). The finding of Ayana (2001) indicated that sorghum accessions collected from more neighboring regions were grouped together. Other supportive results were also reported by different authors (Bezaweletaw et al., 2006; Tsehaye and Kebebew, 2002).
Principal component analysis: Principal component analysis in the present study confirmed the existence of high genetic diversity in finger millet landraces since all traits have their own contribution in a variable degree for the variation observed in every components and since the entire variation was explained in several PCs. Based on the mean of the first two principal components of the three Eigenvectors (population level, country/regional and altitudinal level), grain yield per plant, thousand grain weight, days to heading, days to maturity, lodging index and biomass weight per plant were the most important traits contributing for the overall variability observed among landraces, geographical locations and agro-ecologies. This is an important backup for breeder to investigate for high yielding (both in food and feed aspect), agro-ecology and climate condition (rainfall) based maturity groups and lodging resistant varieties through conventional breeding.
Several authors indicated that different agronomic and phenotypic traits for the different crops have contribution for the overall variability observed between study germplasms and geo-ecological zones of origin (Lule et al., 2011; Negash et al., 2005; Assefa et al., 1999, 2003; Ayana and Bekele, 1998; Quendeba et al., 1995).
CONCLUSION AND RECOMMENDATION
Varietal development, genetic resource collection and conservation depend on the presence of genetic variability in specific characters. The wider genetic variations at lowest level (among populations) than at higher level (among countries and regions of origin) as revealed from descriptive analysis, agro-ecological proximity and the similarity observed between neighboring regions as observed from cluster analysis leads to suggest that taking more samples within a locality or population would be a better approach to capture the range of variation in finger millet population. The similarity manifested for eco-geographical proximity as revealed from cluster analysis and principal coordinates analysis could be either selection by farmers were in fever of same traits across location, the seed source was same originally or it can be due to adaptive role of the traits in the environment. Principal component analysis indicated that all traits considered in the current study have contribution for the overall genetic variations observed among populations and eco-geographical locations of origin.
ACKNOWLEDGMENTS
The authors acknowledge the SIDA-supported Bioinnovate collaborative research project for funding the research, the Biology Department of Addis Ababa University, Bako Agricultural research center and Arsi Negele Agricultural research sub-center for facilitating field trials. We thank the Institute of Biodiversity Conservation for providing landraces with their available passport data.
REFERENCES
- Assefa, K., M. Arnulf and H. Tefera, 2003. Multivariate analysis of diversity of tef (Eragrostis tef (Zucc.) Trotter) germplasm from western and south western Ethiopia. Hereditas, 138: 228-236.
PubMed - Assefa, K., S. Ketema, H. Tefera, H.T. Nguyen and A. Blum et al., 1999. Diversity among germplasm lines of the Ethiopian cereal tef Eragrostis tef (Zucc.) Trotter. Euphytica, 106: 87-97.
CrossRef - Ayana, A. and E. Bekele, 1998. Geographical patterns of morphological variation in sorghum (Sorghum bicolor (L.) Moench) germplasm from Ethiopia and Eritrea: Qualitative characters. Hereditas, 129: 195-205.
CrossRefDirect Link - Babu, B.K., N. Senthil, S.M. Gomez, K.R. Biji, N.S. Rajendraprasad, S.S. Kumar and R.C. Babu, 2007. Assessment of Genetic diversity among fingermillet (Eleusine coracana L. Gaertn) accession using molecular markers. Genet. Res. Crop Evol., 54: 399-404.
CrossRef - Bekele, E., 1984. Analysis of regional patterns of phenotypic diversity in the Ethiopian tetraploid and hexaploid wheats. Hereditas, 100: 131-154.
CrossRefDirect Link - Bekele, E., 1983. Some measures of genetic diversity analysis on landrace population of Ethiopian Barley. Hereditas, 98: 127-143.
CrossRef - Demissie, A. and A. Bjornstad, 1996. Phenotypic diversity of ethiopian barleys in relation to geographical regions, altitudinal range and agro-ecological zones: As an aid to germplasm collection and conservation strategy. Hereditas, 124: 17-29.
CrossRefDirect Link - Ekwamu, A., 1991. Influence of head blast infection on seed germination and yield components of finger millet (Eleusine coracana L. Gaertn). Trop. Pest Manage., 37: 122-123.
CrossRef - Mahalanobis, P.C., 1936. On the generalised distance in statistics. Proc. Nat. Inst. Sci. India, 2: 49-55.
Direct Link - Quendeba, B., G. Ejeta, W.W. Hannan and A.K. Kumar, 1995. Diversity among African pearl millet landrace populations. Crop Sci., 35: 919-924.
Direct Link - Varshney, R.K., K. Chabane, P.S. Hendre, R.K. Aggarwal and A. Graner, 2007. Comparative assessment of EST-SSR, EST-SNP and AFLP markers for evaluation of genetic diversity and conservation of genetic resources using wild, cultivated and elite barleys. Plant Sci., 173: 638-649.
CrossRefDirect Link - Upadhyaya, H.D., C.L.L. Gowda, R.P.S. Pundir, V.G. Reddy and S. Singh, 2006. Development of core subset of finger millet germplasm using geographical origin and data on 14 quantitative traits. Gen. Res. Crop Evo., 53: 679-685.
Direct Link