
Aida A. Rizkalla
Department of Genetics and Cytology, Genetic Engineering and Biotechnology Division, National Research Center, Dokki, Egypt
A.M.F. Al-Ansary
Department of Genetics and Cytology, Genetic Engineering and Biotechnology Division, National Research Center, Dokki, Egypt
S.A.A. Attia
Department of Genetics and Cytology, Genetic Engineering and Biotechnology Division, National Research Center, Dokki, Egypt
A.A.A. Haiba
Department of Genetics and Cytology, Genetic Engineering and Biotechnology Division, National Research Center, Dokki, Egypt
J.E. Nasseef
Department of Genetics and Cytology, Genetic Engineering and Biotechnology Division, National Research Center, Dokki, Egypt
International Journal of Agricultural Research
Year: 2012 | Volume: 7 | Issue: 4 | Page No.: 205-214







ABSTRACT
Wheat anther culture produces homozygous doubled haploid plants, in a very short time span as comparing with traditional breeding methods. It is affected with several different factors, such as suitable spike stage as a donor for anthers, anthers cold pretreatment duration, rate of embryogenic callus formation and the time of anthers transferring from callus induction to regeneration medium. The effects of these factors on androgenic process were studied in this work the response of five F1 wheat hybrids, of ten varieties, differing in their salt tolerance, was investigated in anther culture system for haploid plantlets regeneration. Anthers were plated on P4S medium for callus induction, anthers were taken from different spikes ages (as in a distance between the top of the spike and flag leaf), different duration of cold pretreatment and different time of transferring anthers from callus induction to regeneration medium (190-2) were studied. Significant differences between genotypes on androgenic process were detected. Genotype influence was significant on the suitable spike length which has high percentage of proper stage of mid-uninucleate stage, consequently on both of callus induction and plant regeneration (green, albino and total plantlets). On the other hand, there was no significant influence in the embryogenic callus formation per 100 anthers between the hybrids under same duration (14, 28 and 56 days). The suitable duration for staying on induction callus medium was 28 days as the green regeneration plants increased. This is considered as accurate parameter for evaluation efficiency of androgensis for anther culture.
PDF Abstract XML References Citation
Received: January 27, 2012;
Accepted: April 03, 2012;
Published: June 21, 2012
How to cite this article
Aida A. Rizkalla, A.M.F. Al-Ansary, S.A.A. Attia, A.A.A. Haiba and J.E. Nasseef, 2012. Response of Some Egyptian and Introduced Wheat Hybrids to Androgenic Process. International Journal of Agricultural Research, 7: 205-214.
DOI: 10.3923/ijar.2012.205.214
URL: https://scialert.net/abstract/?doi=ijar.2012.205.214
DOI: 10.3923/ijar.2012.205.214
URL: https://scialert.net/abstract/?doi=ijar.2012.205.214
INTRODUCTION
Anther culture is a competitive technique for traditional methods that plant breeders use. This technique can produce haploids to improve breeding in both mono and dicotyledonous plants to get new lines characterizing with both high yield and disease resistance in a short time (Ramakrishnan et al., 2005). Production of haploid plants with androgenesis in cereals has been applied such as: wheat (Simmonds et al., 1993), Barely (Luckett and Smithard, 1992), Triticale (Charment and Bernard, 1984), Rice (Miah et al., 1985), Maize (Petolino and Thompson, 1987) etc. Doubled haploid production technology, inducing homozygosity, shortens breeding program and permitting variation between genotype within any generation which attracts breeders attention (Maria et al., 2006), reducing cost of cultivar development (Hu and Yang, 1986; Hu, 1997; Liu et al., 2002). In wheat breeding programs obtaining haploid plants resultant from anthers of F1 hybrids through anther culture that accelerates new cultivars development when reduces many years and permitting easier and more efficient selection process (Camergo et al., 1999; Ramos et al., 2000).
Before eighteenth century, developing crops by doubled haploid plants was limited. Androgenesis process can be explained as formation of pseudo embryos (embryoids) which can geminate into plants giving genetically true doubled haploid (DH) plants. This is an important and attractive goal for plant breeders, geneticists and biotechnology researches. DH progeny enables getting inherited traits of different gene combinations (Konzak et al., 1987). Anther culture technique gives a very important advantage of obtaining more than thousand haploid plants/anther culture (Devaux, 1988). In comparing to other methods, all of it are restricted to only one plant/floret and consequently to a low number of responsive genotypes while others remain without changing. Therefore, the urgent need for methods help to increase androgenesis process of microspores in a great number of genotypes (Hu et al., 1995).
In wheat, may be through crosses androgenic response can be passed to progeny in great numbers as it is inherited (Henry and Buyser, 1981; Lazar et al., 1984; Moieni and Sarrafi, 1995). Thus, formation of haploids and their regeneration is affected by three related traits; embryo formation rate, their regeneration capability and ratio of green to albino plants (Henery and Buyser, 1985; Ghaemi et al., 1995; Moieni and Sarrafi, 1995). Regeneration frequency also is affected by the time of transferring anthers from induction medium to regeneration conditions (Zheng and Konzak, 1999; Redha et al., 2000).
Developing of microspore-anther culture methods was getting when stress treatment was applied basing on the transferring from preprogrammed gametophytic to sporophytic pathway for getting androgenesis (Touraev et al., 1996; Touraev et al., 1997; Hu and Kasha, 1999; Zhou and Konzak, 1997; Simonson et al., 1997; Reynolds, 1997; Zheng and Konzak, 1999; Ohnoutkoua et al., 2000).
In view of the above, present work was planned to study the androgenetic process of anther cultures for F1 hybrids of several wheat parents differing in salt tolerance level under different parameters i.e., (1) Proper stage of donor spikes development (in centimeters) as a morphological character (2) Duration of anthers cold pretreatment ( days) (3) Induction of embryogenic callus and embryos formation and (4) The time of transferring anthers from callus induction medium to regeneration medium.
MATERIALS AND METHODS
Plant materials and salt screening: Salt screening and field crossing were done in 2009 and 2010, laboratory work was extending till Dec. 2011.
Ten varieties of bread wheat (Triticum aestivum L.), six were local (Sids 4, Sakha 93, Gemmeiza 9, Sids 1, Sakha 8 and Giza 162) and four introduced (Golan 2, Cham 6, Bow and Mexipak 69), were chosen to test the transactions affected by levels of salinity. The experiment was conducted in Petri dishes containing quarter strength of Hoagland solution (Hoagland and Arnon, 1950) under five salinity levels: 0, 8,000, 10,000, 12,000 and 14,000 ppm of 2:1 (NaCl:KCl) as a source of salinity stress. Each treatment was performed by 5 petri dishes (15 cm in diameter) in 3 replicates, each one contained 100 seeds. The germination percentage was calculated after 2 weeks. Seeds were scored as germinated when the primary roots were greater than 3 mm and the shoots were longer than the seed itself. Seeds contaminated by fungi during the test were ignored (Ye et al., 1987).
| Table 1: | Crosses between selected parents for obtaining F1 hybrids as donor plants for anthers |
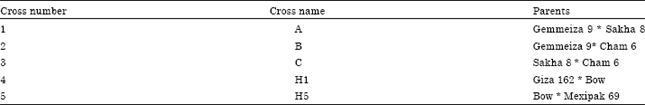 | |
On this base, six parents had been chosen, five of them were most resistant to salinity, while the sixth variety (Gemmeiza 9 is sensitive to salinity but high yielder) to set some crosses as shown in Table 1. The materials of this study were kindly provided by Dr. Samy. A.A. Attia who performed the crosses.
Laboratory work of anther culture, haploid production and regeneration was performed in Genetics and Cytology Department, Genetic Engineering and Biotechnology Division, National Research Center, Egypt.
Determining suitable spike age for anther culture: Anthers of F1 hybrid plants from primary and three secondary tillers were collected when microspores from the anthers in the central region of the spikes were at the mid to late uninucleate stage before the first pollen mitosis (Barnabas et al., 1991; Navarro-Alvarez et al., 1994; Redha et al., 1998). The developmental stage of microspores was checked by acetocarmine staining of anthers from the central region of the spike and subsequent light microscope observation. The sampled spikes at different ages as a distance between the top of the spike and flag leaf were O = 7.5, 10, 12.5 or 15 cm.
Cold pre treatment: Suitable spikes were placed in flasks containing tap water and stored in dark at 4°C for 2-7 or 14 days as an anther stress treatment.
Induction of embryogenic callus and embryos formation: Spikes were surface sterilized in 80% ethanol for 2 min intervals and the enclosed anthers were excised with fine forceps (Zheng and Konzak, 1999). Anthers were then placed aseptically in 50 mL glass jars containing 15 mL of P4S medium (Ouang et al., 1983) to induce embryogenic callus formation at 26±2°C in dark. The largest embryos (>2 mm) produced were then counted and transferred into regeneration medium 190.2 (Zhuang and Jia, 1983), green and albino plantlets were recorded.
The time of transferring anthers from callus induction medium to regeneration medium: The following parameters were recorded to show the effects of the time of transferring anthers from callus induction medium to regeneration medium: embryogenic callus formation plant/100 anthers, regenerated green plants/100 anthers, regenerated albino plants/100 anthers and total regenerated plants/100 anthers after 14, 28 or 56 days on induction medium (P4S) to regeneration medium (190-2). Cytological examination for haploids was done.
RESULTS AND DISCUSSION
Salt screening: As shown in Fig. 1, the ten different wheat varieties had emerged the effect of different levels of salinity on their germination rate were among the terms of higher and lower tolerance salinity. It is noticed that Mexipak 69, Bow Giza 162, Sakha 8 and Cham 6 gave the highest percentage of germination under all salinity levels.
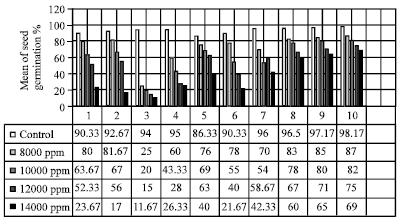 | |
| Fig. 1: | Seed germination percentages of the ten studied cultivars under five salinity levels. 1: Sids 4, 2: Sakha 93, 3: Gemmeiza 9, 4: Sids 1, 5: Sakha 8, 6: Golan 2, 7: Cham 6, 8: Giza 162, 9: Bow, 10: Mexipak 69 |
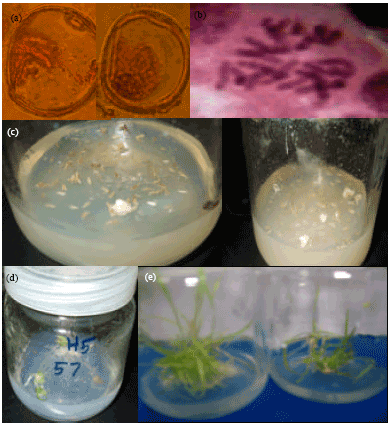 | |
| Fig. 2(a-e): | Developmental stages of anther culture, (a) Suitable stage of anther culture (mid-uninucleate stage), (b) Chromosome number of regenerated haploid plantlets 1n = 3x = 21, (c) Callus formation from anther culture, (d) Initiation of shoot tip, (e) Regenerated plantlets from embryogenic callus |
Suitable spike age for anther culture: According to Ding-Gang and Jun-Wen (1984) and Lantos et al. (2006) the most critical steps of wheat microspore anther cultures is the choice of ideal microspore development stage (Fig. 2a). It is the first important step of androgenesis induction.
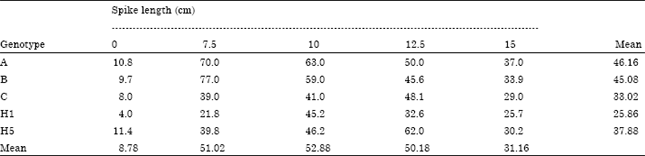
| Table 2: | Percentage of microspores/spike at mid-uninucleate stage induced at different spike stages of the five wheat hybrids (crosses) |
 | |
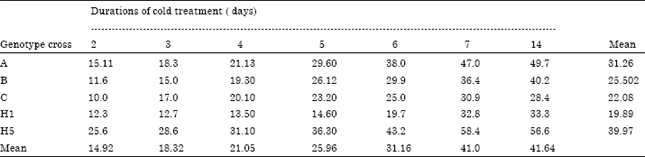
| Table 3: | Percentage of wheat embryogenic callus/100 anthers of the five crosses in response to cold pretreatment durations |
 | |
Data in Table 2 showed the percentage of microspores/spike at mid-uninucleate stage in the central part of spikes of the 5 used hybrids expressed in centimeter as distance between the top of the spike and the flag leaf auricles. Table 2 revealed that spikes at <10 cm stage for the five used F1 hybrids possessed the highest mean frequency of microspore cells in the mid-uninucleate stage (52.88) as compared to the other four spike ages, i.e., 0, 7.5, 12.5 or 15 cm where, their mean values reached 8.78, 51.02, 50.18 and 31.16, respectively. It seems that genotype effect has significant influence on the differences among the mean percentage of microspores/spike in the proper stage of the 5 genotypes. These result is in agreement with Ding-Gang and Jun-Wen (1984), Maluszynski et al. (2001) and Lantos et al. (2006), who stated that percentage of microspores in mid-uninucleate stage in a certain age of spike morphology depend on the genotype. So, correlation between developmental stage of microspores and morphological characters of spike such as distance between the top of the spike (last floret) and flag leaf auricles is important to define the suitable spike age for anther culture. These results also are supported by findings of Mehmet et al. (2008), who concluded that the spike length is significantly correlated with callus number and green plantlet.
Effect of cold pretreatment of spikes on callus induction: Low temperature ~4°C for several days, prior to normal culture procedure, was used under speculation that it might has a dual function: interrupting normal gametophytic development and the nursing effects of anther tissue on microspores (Fazar et al., 1985; Amstrong et al., 1987; Henry and Buyser, 1981). Table 3 showed the response of the 5 used crosses to cold treatment durations as percentage of callus produced from 100 anthers.
The results in Table 3 indicated that cold treatment of spikes for duration shorter than 7 days could not have enough impact on starving microspores to switch their normal pathway to form embryos. On the contrary, starving microspores for 7 or 14 days was highly recommended for the used crosses. Table 3 presented that the five crosses significantly varied as the percentage of cross H5 reached 58.4 and 56.6% under cold duration of 7 and 14 days, respectively.
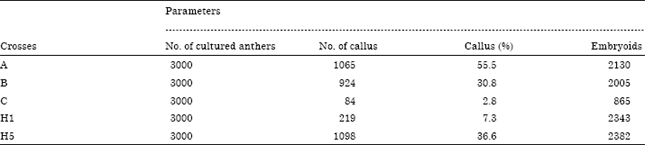
| Table 4: | Number and percentage of callus and embryoids from anthers of the five crosses on inductions (P4S) medium |
 | |
It was followed by cross A which reached 47 and 49.7%, respectively. While, cross C showed the lowest response, its percentages were 30.9 and 28.4, respectively, also the two crosses B and H1 produced intermediate levels of anther response. The diverse of the five genotypes responses for cold pretreatment were attributable to varying genotypes, pretreatment durations and the genotype x pretreatment durations. Present results are in agreement with the findings of Lazar et al. (1984), Amstrong et al. (1987) and Henry and Buyser (1981). In contrary, Khiabani et al. (2008) included that cold pretreatment was found ineffective on androgenic ability of genotypes where they showed low response to cold treatments.
Induction of embryogenic callus and embryos formation: Embryogenic callus formation (Fig. 2c) of the five crosses on induction medium (P4S), their percentages and No. of embryos are in Table 4 which showed that the percentages ranged from, the lowest percentage (2.8) for cross C and the highest (55.5) for the cross A. While crosses H5, B and H1 ranged in between the previous crosses where, their callus formation percentages were 36.6, 30.8 and 7.3%, respectively. These differences among crosses could be attributed to the effects of genetic make-up and or the culture composition, where genotype affected anther culture response significantly (Lu et al., 1991; Konieczny et al., 2003; Bagheri and Jelodar, 2008; Zheng et al., 2001).
Others suggest that these differences in androgenic response could be referred to the presence of several genes located on the different wheat genome (Szakacs et al., 1988; Sibikeeva and Sibikeev, 1996). Others referred the difference in androgenic responses, of used genotypes, to their differences in endogenous hormones levels , which in turn in different requirements for the culture medium.
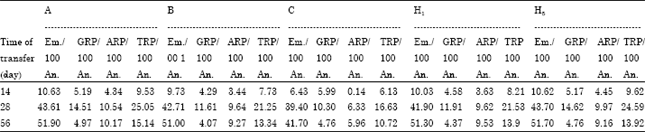
Effect of transferring wheat anthers (from induction to regeneration medium) on the androgenic response, of the five studied crosses: Data in Table 5, showed the percentages of embryogenic callus, green regenerated plants (Fig. 2d-e), albino regenerated plants and total number of regenerated plants/100 anther after 14, 28 or 56 days on induction medium (P4S) to regeneration medium (190-2). From Table 5, we notice that, there were no significant differences in the embryogenic callus formation per 100 anthers between the five studied crosses under the same duration (14, 28 or 56 days). On the other hand, we find that 28 days duration is the proper time of transferring for obtaining higher percentages of green regenerated plants as well as for albino and total regenerated plants (haploid 1n = 3x = 21, Fig. 2b).
| Table 5: | Percentages of embryogenic callus, green, albino and total regenerated plantlets per 100 anthers |
 | |
| Em.: Embryos, An: Anthers, GRP: Green regenerated plants, ARP: Albino regenerated plants, TRP: Total regenerated plants | |
Data showed great increment percentage of green regenerated plants at 28 days than 14 or 56 days, where for cross (a) the increment reached 2.8 and 2.9 fold, respectively; while for cross (b) it reached 2.7 and 2.85 fold; for cross (c) it was 1.71 and 2.16 fold, respectively; for cross (H1) it increased 2.6 and 2.72 fold; while for cross (H5) it was the highest increasing 2.82 and 3.07 fold.
In terms of regeneration, 28 days duration was the best as the percentage of green regenerated plants is considered as accurate parameter for efficiency evaluation of anther culture.
On the other hand, in comparing with 14 days, the percentages of the albino regenerated plants increased greater with 28 days, while the greatest increment was with 56 days for all the studied crosses which reflects as a problem for the application of anther culture technique in wheat breeding programs as Saidi et al. (1997) stated that, production of albino plants was affected by genotype and medium composition. They added that albinism might occur by mutation in chloroplast DNA. While Caredda et al. (1999) referred albinism to the probability of the abnormalities of plastids features during embryogenesis process.
The early transfer of anthers (that formed callus) from the induction medium to regeneration medium caused a strong reduction in the production of embryogenic callus, while the overall regeneration of plants/100 anthers showed an improvement for delaying. These results are in agreement with previous findings on wheat (Henery and Buyser, 1985; Redha et al., 2000). The reduction of embryogenic callus production with the early transfer, suggesting that, the earlier transfer interferes with the development of the embryos. However, it had a positive effect on the quality of the embryos and, as a result, the regeneration of the plants improved considerably. When the embryos are left for longer time (56 days) on induction medium, the quality of the embryos and their totipotency to regenerate may be lower (Redha et al., 2000).
CONCLUSION
Anthers of five F1 wheat hybrids, their parents differ in salt tolerance showed a variation in the productive response of callus formation and regeneration capacity (green, albino and total plant numbers) under different studied factors. It was concluded that androgenic response is affected mainly by genotype which has significant influence on the differences among the mean percentage of microspores/spike in the anther proper stage of the 5 used genotypes. While, embryogenic callus formation was better in response to 7 days of anthers cold pretreatment durations. Also the proper time of transferring calluses from callus induction medium to regeneration medium affected androgenic process.
REFERENCES
- Amstrong, T.A., S.G. Metz and P.N. Mascia, 1987. Two regeneration systems for the production of haploid plants from wheat anther culture. Plant Sci., 51: 231-237.
Direct Link - Bagheri, N. and N.B. Jelodar, 2008. Combining ability and heritability of callus induction and green-plant regeneration in rice anther culture. Biotechnology, 7: 287-292.
CrossRefDirect Link - Barnabas, B., P.L. Pfahler and G. Kovacs, 1991. Direct effect of colchicine on the microspore embryogenesis to produce dihaploid plants in wheat (Triticum aestivum L.). Tag. Theor. Applied. Genet., 81: 675-678.
CrossRef - Caredda, S., P. Devaux, R.S. Sangwan and C. Clemet, 1999. Differential development of plastids microspore embryogenesis in barely. Protoplasma, 208: 248-256.
CrossRefDirect Link - Devaux, P., 1988. Comparison of anther culture and the Hordeum bulbosum method for the production of doubled haploids in winter barely. Plant Breed., 100: 181-187.
CrossRefDirect Link - Ghaemi, M., A. Sarrafi and R. Morrise, 1995. Reciprocal substitutions analysis of embryo induction and plant regeneration from anther culture in wheat (Triticum aestivum L.). Genome, 38: 158-165.
PubMed - Ding-Gang, H. and O. Jun-Wen, 1984. Callus and plantlet formation from cultured wheat anthers at different developmental stages. Plant Sci. Lett., 33: 71-79.
CrossRefDirect Link - Henery, Y. and J. Buyser, 1985. Effect of 1B/1R translocation on anther culture ability in wheat (Triticum aestivum L.). Plant Cell Rep., 4: 307-310.
CrossRef - Hu, T.C., A. Ziaudin, E. Simion and K.J. Kasha, 1995. Isolated microspore culture of wheat (Triticum aestivum L.) in a defined medium I. Effects of pretreatment, isolation methods and hormones. In vitro Cell Dev. Biol-Plant, 31: 79-83.
CrossRef - Konieczny, R., A.Z. Czaplicki, H. Golczyk and L. Przymara, 2003. Two pathways of plant regeneration in wheat anther culture. Plant Cell Tissue Organ. Cult., 73: 177-187.
CrossRef - Lantos, C., S. Paricsi, A. Zofajova, J. Weyen and J. Pauk, 2006. Isolated microspore culture of wheat (Triticum aestivum L.) with hangarian cultivars. Acta Biol. Szegediensis, 50: 31-35.
Direct Link - Liu, W., M. Zheng and C. Konzak, 2002. Improving green plant production via isolated microspore culture in bread wheat (Triticum aestivum L.). Plant Cell Rep., 20: 821-824.
CrossRef - Lu, C.S., H.C. Sharma and H.W. Ohm, 1991. Wheat anther culture: Effects of genotype and environmental conditions. Plant Cell Tissue Organ. Cult., 24: 233-236.
CrossRef - Luckett, D.J. and R.A. Smithard, 1992. Doubled haploid production by anther culture for Australian barely breeding. Aust. J. Argic. Res., 43: 67-78.
CrossRef - Maluszynski, M., I. Szarejko, P. Barriga and A. Balcerzyk, 2001. Heterosis in crop mutant crosses and production of high yielding lines using doubled haploid systems. Euphytica, 120: 387-398.
CrossRefDirect Link - Miah, M.A.A., E.D. Earle and G.S. Khush, 1985. Inheritance of callus formation ability in anther cultures of rice, Oryza sativa L. Theor. Applied Genet., 70: 113-116.
Direct Link - Moieni, A. and A. Sarrafi, 1995. Genetic analysis for haploid regeneration responses of hexaploid wheat anther cultures. Plant Breed., 114: 247-249.
CrossRef - Ohnoutkoua, L., J. Novotny, E. Mullerova, J. Vagera and L. Kucera, 2000. Is a cold pretreatment really needed for induction of in vitro androgensis in barely and wheat. Proceedings of the COST Action on 824 Biotichnological Approaches for Utilisation of Genetic Cells, July 1-5, 2000, Bled, Slvenia, pp: 33-37.
- Petolino, J.F. and S.A. Thompson, 1987. Genetic analysis of anther culture response in maize. Theoret. Appl. Genet., 74: 284-286.
CrossRef - Redha, A., S.M.S. Islam, B. Buter, P. Stamp and J.E. Schmid, 2000. The improvement in regenerated doubled haploids from anther culture of wheat by anther transfer. Plant Cell Tiss. Organ. Cult., 63: 167-172.
CrossRef - Sibikeeva, Y.E. and S.N. Sibikeev, 1996. Genetic analysis of anther culture response in wheat carrying alien translocation. Tag. Theor. Applied Gene., 92: 782-785.
CrossRef - Simonson, R.L. P.S. Baenziger and V.D. Gustafson, 1997. Wheat anther culture as affected by various cultural changes and supplements. J. Appl. Genet., 38: 381-392.
Direct Link - Szakacs, E., G. Kovaes, J. Pauk and B. Barmabas, 1988. Substitution analysis of callus induction and plant regeneration from anther culture in wheat (Triticum aestivum L.). Plant Cell Rep., 7: 127-129.
CrossRef - Touraev, A., A. Indrianto, I. Wratschko, O. Vicente and E. Heberle-Bors, 1996. Efficient microspore embryogenesis in wheat (Triticum aestivum L.) induced by starvation at high temperature. Sex Plant Reproduc., 9: 209-215.
CrossRef - Touraev, A., O. Vicente and E. Heberle-Bors, 1997. Initiation of microspore embryogenesis by stress. Trends Plant Sci., 2: 297-302.
CrossRef - Ye, J.M., K.N. Kao, B.L. Harvey and B.G. Rossangel, 1987. Screening salt-tolerant barely genotypes via F1 anther culture in salt stress media. Tag. Theor. Applied Genet., 74: 426-429.
CrossRef - Zheng, M.Y. and C.F. Konzak, 1999. Effect of 2,4-Dichlorophenoxy acetic acid on callus induction and plant regeneration in anther culture of wheat (Triticum aesitivum L.). Plant Cell Rep., 19: 69-73.
CrossRefDirect Link - Zheng, M., W. Liu, Y. Weng, E. Polle and C. Konzak, 2001. Culture of freshly insolated wheat (Triticum aestivum L.) microspores treated with inducer chemicals. Plant Cell Rep., 20: 685-690.
CrossRef - Zhou, H. and C.F. Konzak, 1997. Influence of genetic and environmental factors on anther culture response of wheat. J. Applied Gen., 38: 339-406.
Direct Link - Ramakrishnan, S.H., S. Saravanan, C.R. Anandakumar and J.R. Kannanbapu, 2005. In vitro androgenesis in rice (Oryza sativa L.). Asian J. Plant Sci., 4: 600-602.
CrossRefDirect Link