
V. R. Pandey
Department of Genetics and Plant Breeding, Narendra Deva University of Agriculture and Technology, Kumarganj, Faizabad-224229, Uttar Pradesh, India
P. K. Singh
Department of Genetics and Plant Breeding, Narendra Deva University of Agriculture and Technology, Kumarganj, Faizabad-224229, Uttar Pradesh, India
O. P. Verma
Department of Genetics and Plant Breeding, Narendra Deva University of Agriculture and Technology, Kumarganj, Faizabad-224229, Uttar Pradesh, India
P. Pandey
Department of Genetics and Plant Breeding, Narendra Deva University of Agriculture and Technology, Kumarganj, Faizabad-224229, Uttar Pradesh, India
International Journal of Agricultural Research
Year: 2012 | Volume: 7 | Issue: 4 | Page No.: 169-184







ABSTRACT
Crop yield mostly depends on net result of yield contributing traits, action of genes and their interactions with environment. Hence, an experiment was undertaken with fifty two rice genotypes to know genetic relationship of various polygenic traits on yield. All the genotypes were grown in a randomized complete block design with three replications during Kharif 2009 under salt affected soil (pH of 9.2 and EC 1.61 dS m-1). Results showed that substantial amount of variability exists in the entire gene pool for all traits studied validated ample scope of selection. Magnitude of Phenotypic Coefficient of Variation (PCV) along with Genotypic Coefficient of Variation (GCV) was recorded high for biological yield per plant; moderate for grain yield per plant, spikelets per panicle and flag leaf area. However, days to 50% flowering, days to maturity, plant height, panicle length, spikelet fertility (%), 1000-grain weight and harvest-index had low estimates. High heritability coupled with high genetic advance in percent of mean were observed for biological yield per plant, grain yield per plant, spikelets per panicle and flag leaf area while, the 1000-grain weight and harvest-index suggesting preponderance of additive gene action in the expression of these characters. Grain yield per plant exhibited a very strong positive association with biological yield per plant, flag leaf area and spikelet fertility at phenotypic and genotypic level which indicated that these traits were the strongest associates of grain yield per plant. However, significant negative correlation was observed in case of harvest-index with biological yield per plant, and 1000-grain weight with effective tillers per plant. The genetic reasons for this type of negative association may be linkage or pleiotropy. Path analysis identified harvest-index, days to maturity and effective tillers per plant, 1000-grain weight and flag leaf area and panicle length as major direct contributors. Hence, these traits should be given much emphasis on the selection criteria for genetic improvement of rice yield in salt stress environment.
PDF Abstract XML References Citation
Received: February 14, 2012;
Accepted: April 11, 2012;
Published: June 21, 2012
How to cite this article
V. R. Pandey, P. K. Singh, O. P. Verma and P. Pandey, 2012. Inter-relationship and Path Coefficient Estimation in Rice under Salt Stress Environment. International Journal of Agricultural Research, 7: 169-184.
DOI: 10.3923/ijar.2012.169.184
URL: https://scialert.net/abstract/?doi=ijar.2012.169.184
DOI: 10.3923/ijar.2012.169.184
URL: https://scialert.net/abstract/?doi=ijar.2012.169.184
INTRODUCTION
Rice (Oryza sativa L.) is the most important staple food crop of the world because of being the major source of calories of more than half of the total global population. To focus attention on the importance of rice in global food security and the necessity to increase rice production and productivity, United Nations General Assembly in 2002 declared to celebrate the year 2004 as “International Year of Rice” with the theme of “Rice is Life” (FAO, 2004). This dedication of an International Year to Rice, a single crop, was not only to underline its importance as a fundamental commodity and primary food source for more than half of the world’s population, but also to create a focus on complex rice based ecosystems that influence issues of global concern such as food security development. More than 90% of the world’s rice is grown and consumed in Asia, known as rice bowl of the world, where 60% of the earth’s people and two third of world’s poor live (Khush and Virk, 2000).
Although, the average productivity of rice is much lower in India than the average productivity at world level (Siddiq, 2000). The development of high yielding, widely adapted pureline rice varieties coupled with advances in production technology in past four decades has enabled us to cope up with the demand of rice to a satisfactory level. However, rapidly increasing demand due to ever increasing Indian population has forced us to search for another quantum jump in rice production. Considering the rapidly decreasing availability of agricultural land due to urbanization and industrialization, utilization of different eco-systems and under exploited stress environments for rice production would be needed. Thus, adoption of high yielding rice varieties to various stress environments and under-utilized lands such as salt affected soils would be an important strategy to meet this challenge.
Rice is grown under varying eco-systems on a variety of soil under varying climatic and hydrological conditions ranging from waterlogged and poorly drained to well-drained situations (Chalam, 2006). Every year more and more land is becoming non-productive because of salt accumulation in soil in coastal as well as certain inland saline tracks. Salinity is a serious problem affecting one third of all irrigated land in the world (Munns, 2002; Tester and Davenport, 2003).
In India most of the research bounded for irrigated and non-stress environments while a huge fraction of land under rice in country is salt affected, but the prospective for productivity gains in such areas has not been exploited. Rice is one of the most suitable crops for saline soil, though it is considered moderately sensitive to salinity. The common practice is to cultivate rice with elevated level of salt tolerance on salt affected marginal lands. Today, the most economic and sustained way to overcome the problems of food scarcity and salt stress is to develop salt tolerant varieties. Salinity affects rice growth in varying degrees at all stages starting from germination to maturation (Manneh, 2004). So far, conventional breeding methods for salt tolerance have been found ineffective due to the strong environmental effects on genotypic expression and the low narrow sense heritability of salt tolerance (Gregorio, 1997; Gregorio and Senadhira, 1993).
Collection of superior alleles into a single targeted genotype is necessary to formulate efficient and economic crop improvement programme (Wang and Wolfgang, 2007; Tripathi et al., 2011). Further, substantial genetic variability is essential for initiating an effective breeding programme and therefore, it becomes imperative to study the level of genetic variability to work out an efficient selection criterion (Reynolds et al., 2001). The available variability in a population can be partitioned into heritable and non-heritable components (Falconer, 1989) with the aid of genetic parameters such as coefficients of variation, heritability and genetic advance to serve as basis for selection of some outstanding genotypes from existing ones.
Correlation coefficient is a statistical measure, which is used to find out the degree (strength) and direction of relationship between two or more variables. The association between two variables, which can be directly observed, is termed as phenotypic correlation. It includes both genotypic and environmental effects. Genotypic correlation may be either due to pleiotropy action of genes or due to linkage or more likely both (Mode and Robinson, 1959; Falconer, 1989). The pleiotropy or linkage may involve two desirable traits or one desirable and one undesirable trait. The first situation enhances the genetic improvement, whereas the second hinders the progress. The coefficient of correlation expresses association between two variables but tells us nothing about the causal relations of variables. Therefore, the use of path-coefficient analysis is necessary. Path-coefficient is simply a standardized partial regression coefficient, which splits the correlation coefficient into the measures of direct and indirect effects (Wright, 1923). It reveals relative importance of yield-contributing characters thus, is useful in indirect selection. Keeping the above aspects, present experiment intended to construct selection criteria for rice yield improvement under salt stress environments.
MATERIALS AND METHODS
Experimental detail: The experimental material for the present investigation comprised of 50 genotypes of cultivated rice and 2 standard check varieties (Usar Dhan-3 and Sarjoo-52). The material, comprising exotic lines, exhibited wide spectrum of variation for various agronomic and morphological characters. All the genotypes were grown in salt affected soil under irrigated condition during Kharif, 2009 at Main Experiment Station of Narendra Deva University of Agriculture and Technology, Kumarganj, Faizabad (UP) India. This Experimental site is situated between 26.47°N latitude, 82.12°E longitude and at an altitude of 113 m above the mean sea level in the gangetic plains of eastern Uttar Pradesh. The soil type of experimental site was sandy loam, low in organic carbon, nitrogen and phosphorus and rich in potash with pH of 9.2 and EC 1.61 dS m-1. All entries were evaluated in Randomized Complete Block Design with three replications. Each plot consisted of 3 rows of 3 m length following between rows and within rows spacing of 20 and 15 cm, respectively. The recommended cultural practices were followed to raise a good crop.
Data collection: The observations were recorded on 10 randomly selected competitive plants in each plot. The characters studied were days to 50% flowering, days to maturity, flag leaf area (cm2), plant height (cm), effective tillers per plant, panicle length (cm), spikelets per panicle, spikelet fertility (%), 1000-grain weight (g), biological yield per plant (g), grain yield per plant (g) and harvest-index (%). The general reference for data collection was standard evaluation system for rice (Anonymous, 2002).
Statistical analysis: The data on twelve quantitative characters from the experiment were utilized for estimation of various genetic parameters. PCV and GCV were calculated by the formula given by Burton (1952), heritability in broad sense (h2) by Burton and de Vane (1953) and genetic advance i.e., the expected genetic gain were calculated by using the procedure given by Johnson et al. (1955). Correlation coefficient and path coefficient was worked out as method suggested by Searle (1961), Dewey and Lu (1959), respectively. The estimated values were compared with table values of correlation coefficient to test the significance of correlation coefficient prescribed by Fisher and Yates (1997).
RESULTS
The highly significant mean squares for genotypes for the twelve characters in the analysis of the variance (Table 1) for the design of experiment indicated highly significant differences among the fifty-two rice genotypes for these characters in salt affected soil conditions.
| Table 1: | Analysis of variance for polygenic traits of rice under salt affected soil |
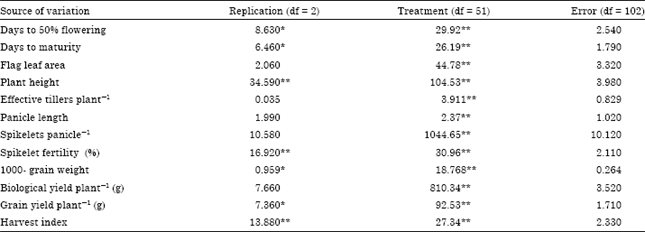 | |
| *, **Significant at 5 and 1% probability levels, respectively | |
| Table 2: | Genetic parameters for different polygenic traits in rice under salt affected soil |
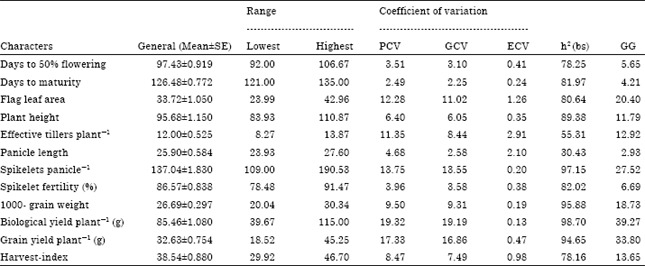 | |
| h2 (bs): Heritability in broad sense, GG: Genetic gain or genetic advance expected in percent of mean, GCV, PCV and ECV: Genotypic phenotypic and environmental coefficient of variation | |
The variances due to replications were highly significant in case of plant height, spikelet fertility and harvest-index and significant in case of days to 50% flowering, days to maturity, 1000-grain weight and grain yield per plant. This revealed existence of substantial variation for characters under study and validated ample scope of selection.
The highest estimates of phenotypic (PCV) and genotypic (GCV) coefficient of variation were observed in case of biological yield per plant (19.32, 19.19%) followed by grain yield per plant (17.33, 16.86%), spikelets per panicle (13.75, 13.55%) and flag leaf area (12.28, 11.02%). Effective tillers per plant possessed moderate PCV (11.35 %), with low GCV (8.44%). Remaining seven characters exhibited low estimates of PCV as well as GCV. The lowest estimates of PCV and GCV were observed for days to maturity (2.49, 2.25%). The estimates of Environmental Coefficient of Variation (ECV) were found to be below for all the twelve characters under study. The magnitude of phenotypic coefficient of variation was higher than the genotypic and environmental coefficient of variation for all the characters (Table 2).
The highest estimate of heritability in broad sense was recorded by biological yield per plant (98.70%), followed by spikelets per panicle (97.15%), 1000-grain weight (95.88%) and grain yield per plant (94.65%). In addition to the above four characters, the other traits exhibiting high estimates of heritability were plant height (89.38%), spikelet fertility (82.02%), days to maturity (81.97%), flag leaf area (80.64%), days to 50% flowering (78.25%) and harvest-index (78.16%). The effective tillers per plant (55.31%) showed moderate heritability (50% to 75%). Panicle length recorded low heritability besides having lowest value (30.43 %) of heritability among all traits under study (Table 2).
The highest value of genetic advance in percent of mean was shown by biological yield per plant (39.27%), while panicle length (2.93%) had lowest value. The characters exhibiting high estimates of genetic advance in percent of mean were grain yield per plant (33.38%), spikelets per panicle (27.52%) and flag leaf area (20.40%). The moderate estimates of genetic advance resulted in case of 1000-grain weight (18.73%), harvest-index (13.65%), effective tillers per plant (12.92%) and plant height (11.79%).
The grain yield per plant exhibited highly significant positive correlation (Table 3) with biological yield per plant (0.909**, 0.934**), flag leaf area (0.368**, 0.421**) and spikelet fertility (0.285*, 0.306*). Remaining eight characters showed non-significant correlations with grain yield per plant at both genotypic and phenotypic level. Days to 50% flowering exhibited highly significant positive correlation with days to maturity (0.947**, 0.922**). Flag leaf area recorded significant positive association with biological yield per plant (0.339*, 0.377*) and panicle length (0.333*, 0.494*), while spikelet fertility had significant positive correlation with days to 50% flowering (0.310*, 0.403*), plant height (0.292*, 0.368*) and days to maturity (0.282*, 0.392*). Spikelets per panicle possessed significant and positive association with effective tillers per plant (0.277*, 0.357*).
However, harvest-index showed highly significant and negative correlation with biological yield per plant (-0.566**, -0.625**) and 1000-grain weight with effective tillers per plant (-0.313*, -0.417*). The estimates of phenotypic and genotypic correlation coefficients for the rest of the character pairs were non-significant. It is very interesting to mention here that a close parallelism was observed in genotypic correlation in direction with their corresponding phenotypic correlation coefficients (Table 3).
The path-coefficient analysis (Table 4) was carried out by using phenotypic as well as genotypic correlation coefficients to resolve direct and indirect effects of different characters on grain yield per plant. At phenotypic level, harvest-index (1.892) exerted very high order positive direct effect on grain yield per plant, while biological yield per plant (-2.282) showed very high order negative direct effect on grain yield per plant. Substantial positive and negative direct effects on grain yield per plant were contributed by days to maturity (0.119) and days to 50% flowering (-0.143), respectively. The direct effects of other seven characters were too low to be considered for any consequence.
Biological yield per plant (1.720), flag leaf area (0.697), spikelet fertility (0.539), days to maturity (0.420), effective tillers per plant (0.393), days to 50% flowering (0.378), spikelets per panicle (0.377), panicle length (0.232) and 1000-grain weight (0.169) exerted high order positive indirect effects on grain yield per plant via harvest-index. Plant height (-0.218) showed high order negative indirect effect on grain yield per plant via harvest-index. Harvest-index (-2.074), followed by flag leaf area (-0.775), spikelet fertility (-0.481), effective tillers per plant (-0.392), days to maturity (-0.375), days to 50% flowering (-0.369), spikelets per panicle (-0.357) and panicle length (-0.303) exhibited high order negative indirect effects on grain yield per plant via biological yield per plant.
| Table 3: | Estimates of correlation coefficients between different polygenic traits of rice under salt affected soil |
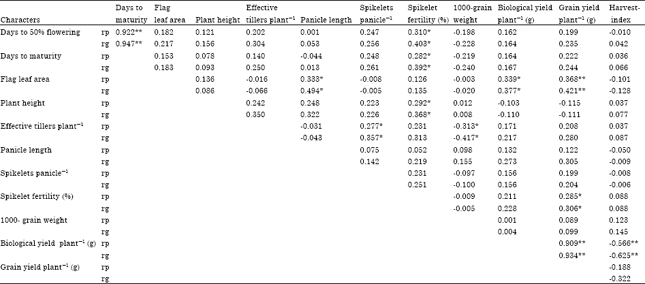 | |
| *,**significant at 5 and 1% probability level, respectively, rp and rg: Phenotypic and genotypic correlation, respectively | |
| Table 4: | Direct and indirect effects of polygenic traits on grain yield in rice under salt affected soil |
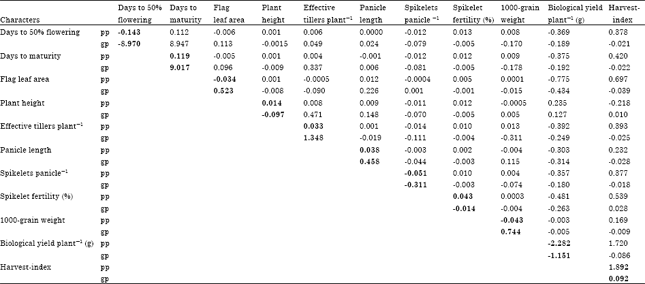 | |
| *Residual effect: 0.056, bold figures indicate the direct effects, pp and gp: Path coefficient at phenotypic and genotypic level, respectively | |
In contrast, plant height (0.235) made substantial positive indirect contribution on grain yield per plant via biological yield per plant. Days to 50% flowering (0.112) contributed considerable positive indirect effect on grain yield per plant via days to maturity which showed high order negative indirect effect (-0.135) via days to 50% flowering on grain yield per plant. The rest of the estimates of indirect effects obtained in path analysis at phenotypic level were negligible. The estimate of residual factors (0.056) obtained in the phenotypic path analysis was very low.
It was revealed that very high order positive direct effect on grain yield per plant exhibited by days to maturity (9.017) at genotypic level. Effective tillers per plant (1.348), 1000-grain weight (0.744), flag leaf area (0.523) and panicle length (0.458) exerted high order positive direct effects on grain yield per plant. Days to 50% flowering (-8.970) showed very high negative direct contribution towards grain yield per plant, while direct effects of biological yield per plant (-1.151) and spikelets per panicle (-0.311) were also high order negative. The direct effects of rest of the characters on grain yield were negligible. The estimate of residual factor in this analysis was also low (0.118).
DISCUSSION
The phenotypic, genotypic and environmental coefficients of variation were computed to assess the nature and magnitude of existing variability in the experimental material. The high magnitude of Phenotypic Coefficient of Variation (PCV) along with Genotypic Coefficient of Variation (GCV) was recorded for biological yield per plant. This indicated greater scope of obtaining high selection response for this trait owing to presence of high genetic variability. The existence of high genetic variability for biological yield per plant in rice is in conformity with the findings of earlier workers (Singh and Chaudhary, 1996; Mishra and Verma, 2002; Karthikeyan et al., 2007; Panwar et al., 2007; Pandey et al., 2009, 2010; Yadav et al., 2011).
The moderate estimates of coefficients of variation at genotypic and phenotypic level were observed for grain yield per plant, spikelets per panicle and flag leaf area. Kumar and Ramesh (2008) observed moderate to high PCV and GCV values for grain yield per plant while Singh et al. (2008) found moderate PCV and GCV estimates for grain yield per plant and number of effective tillers per plant. The three characters exhibiting moderate PCV and GCV values, as mentionable above, are likely to allow reasonable scope of improvement through selection due to moderate genetic variability available in the rice lines evaluated.
Effective tillers per plant showed moderate PCV with low GCV which indicated chances of obtaining some improvement through selection. Low estimates of PCV and GCV parameters were observed for days to 50% flowering, days to maturity, plant height, panicle length, spikelet fertility (%), 1000-grain weight and harvest-index. Khedikar et al. (2004) also reported low estimates of genotypic and phenotypic coefficients of variation for days to 50% flowering ad plant height. The occurrence of low estimates of genotypic and phenotypic coefficient of variation indicated that selection directly based on these traits would not be much rewarding.
The estimates of phenotypic coefficient of variation were slightly higher than the corresponding genotypic coefficient of variation for all the 12 characters under study. Singh and Chaudhry (1996), Pattanayak and Gupta (1999) and Kumar et al. (2001) also reported that phenotypic coefficients of variability were higher than their respective genotypic coefficients of variability for all the traits studied by them.
The low estimates of environmental coefficients of variation were possessed by all the characters. This indicated comparatively lesser contribution of environmental factors in expression of variability in case of all the traits under study, which represents a favourable situation for better efficiency of selection for these characters in salt affected soil conditions.
The fundamental principle involved in plant breeding is the application of selection on the genetic variability available in breeding materials for various characters in order to develop improved genotypes possessing higher economic yield and value than existing ones. Obviously, genetic variability is the raw material on which selection acts to bring improvement in genetic architecture of plants. The proportion of genetic variability which is transmitted from parents to offspring is reflected by heritability (Lush, 1949). Heritability in broad sense and genetic advance in percent of mean as direct selection parameters provide index of transmissibility of traits which gives indication about the effectiveness of selection in improving the characters (Johnson et al., 1955).
The high estimates of heritability in broad sense with high genetic advance in percent of mean were observed for biological yield per plant, grain yield per plant, spikelets per panicle and flag leaf area. High estimates of heritability and genetic advance have also been reported earlier for grain yield per plant (Sawant and Patil, 1995; Anand et al., 1998; Nayak et al., 2002; Yadav et al., 2002; Mahto et al., 2003; Chand et al., 2004; Sharma and Sharma, 2005; Devi et al., 2006; Manickavelu et al., 2006; Kumar et al., 2007; Bagheri et al., 2008; Sarangi et al., 2009), spikelets per panicle (Yadav et al., 2002; Sharma and Sharma, 2005; Suman et al., 2005; Devi et al., 2006), flag leaf area (Sharma and Sharma, 2005; Manna et al., 2006; Sarangi et al., 2009) and biological yield per plant (Thakur et al., 2000; Suman et al., 2005; Kumar et al., 2007; Pandey et al., 2010; Yadav et al., 2011). The four characters, mentioned above, having high values of heritability and genetic advance in percent of mean emerged as ideal traits for improvement through selection. These traits also showed moderate to high GCV and PCV values which suggested that the lines evaluated and/or segregating generations derived from them may provide very high response to selection for the characters exhibiting high heritability along with high genetic advance owing to their high transmissibility and variability.
The 1000-grain weight, harvest-index and plant height showed high heritability in broad sense coupled with moderate genetic advance in percent of mean with low PCV and GCV values. This indicated possibility of obtaining reasonable response to selection in these traits owing to their high transmissibility even if variability was of lower degree. Bughio et al. (2009) reported high heritability with moderate genetic advance for plant height and 1000-grain weight. Effective tillers per plant had moderate estimates of heritability, genetic advance in percent of mean and PCV with low GCV which indicated chances of obtaining considerable selection response due to moderate transmissibility and variability.
The high estimates of heritability with low genetic advance, PCV and GCV value were recorded for days to 50% flowering, days to maturity and spikelet fertility which suggested that the chances of obtaining desirable selection response for improving these three traits by the breeding efforts based on the genotypes evaluated are bleak. The apparent lack of genetic variability in spite of high transmissibility makes these traits unreliable indices for selection. High heritability with low genetic advance was also recorded by Gupta et al. (1999) for days to 50% flowering and Madhavilatha et al. (2005) for days to maturity and spikelet fertility percentage. Panicle length was the only trait which showed very low estimate of heritability, genetic advance in percent of mean, GCV and PCV values to reveal existence of very low transmissibility and variability for this character in the material of the present investigation. Thus, improvement in panicle length through selection procedures would be a difficult and tedious job in respect of breeding populations developed involving rice genotypes evaluated in the present study.
The grain yield or economic yield, in almost all the crops, is referred to as super character which results from the multiplicative interactions of several other characters that are termed as yield components. Thus, genetic architecture of grain yield, in rice as well as other crops, is based on the balance or overall net effect produced by various yield components directly or indirectly by interacting with one another. Therefore, identification of important yield components and information about their association with yield and also with each other is very useful for developing efficient breeding strategy for evolving high yielding varieties. In this respect, the correlation coefficient which provides symmetrical measurement of degree of association between two variables or characters, help us in understanding the nature and magnitude of association among yield and yield components.
The genotypic correlation coefficients between different characters were generally similar in sign or nature to the corresponding phenotypic correlation coefficients in the experiment. The estimates of genotypic correlation coefficients showed close parallelism in direction with their corresponding phenotypic correlation coefficients. However, genotypic correlations were higher in magnitude than the corresponding phenotypic correlations. The occurrence of higher estimates of genotypic correlations than the corresponding phenotypic correlations between yield and yield components in rice has also been reported by Reddy et al. (1997).
In the present study, a very strong positive association of grain yield per plant was observed with biological yield per plant and flag leaf area at genotypic as well as phenotypic level. Grain yield per plant also recorded strong positive association with biological yield per plant at environmental level. Spikelet fertility was another trait having strong positive association with grain yield per plant at phenotypic as well as genotypic level. Thus, biological yield per plant, followed by flag leaf area and spikelet fertility emerged as most important associate of grain yield per plant in rice under salt affected soil. The positive association of grain yield per plant with biological yield per plant, flag leaf area and spikelet fertility has also been observed by previous workers (Panwar et al., 1997; Bagali et al., 1999; Sarawgi et al., 2000; Shivani and Reddy, 2000; Prasad et al., 2001; Sashidhar et al., 2005; Bughio et al., 2009; Sarangi et al., 2009; Pandey et al., 2009; Yadav et al., 2011).
Harvest-index showed very strong negative correlation with biological yield per plant at phenotypic as well as genotypic level. This association appears logical because higher biomass production may reduce the harvest-index if partition of photosynthates towards the reproductive phase is of lower degree.
Spikelet fertility exhibited significant and positive correlation at phenotypic as well as genotypic level with days to 50% flowering, days to maturity and plant height, besides having similar association with grain yield per plant. This suggested that late flowering, late maturing and taller genotypes had high spikelet fertility contributing towards higher grain yield. Similarly, flag leaf area exhibited positive and significant phenotypic and genotypic correlations with panicle length and biological yield per plant along with grain yield per plant. Thus, the genotypes having higher flag leaf area appear to produce higher grain yield per plant and biological yield per plant. The estimates of correlation coefficients mentioned above represent a highly favourable situation for providing correlated response through selection leading to improvement in grain yield per plant and its most important associates like spikelet fertility and flag leaf area through other traits to which these characters had positive association such as panicle length and biological yield with flag leaf area and days to maturity, days to 50% flowering and plant height with spikelet fertility.
The highly significant and positive genotypic and phenotypic correlation between days to 50% flowering and days to maturity between them appears logical as late onset of flowering in likely to push genotypes towards late maturity. Zafar et al. (2006) have also observed positive correlation between days to flowering and days to maturity. Effective tillers per plant had strong positive and negative association at genotypic as well as phenotypic level with spikelets per panicle and 1000-grain weight, respectively.
Furthermore, grain yield per plant showed positive association at genotypic and phenotypic level with biological yield, flag leaf area and spikelet fertility. The occurrence of only positive associations of significant level of grain yield with some of its component traits and positive association between most of the yield components reveals less complex inter-relationships between yield and yield components. Such situation is favourable from breeding point of view because selection for one trait may bring correlated response for improvement of other traits which are positively associated with it. The existence of positive associations between grain yield and yield components in most of the cases for providing rapid improvement of characters due to correlated response during selection is in agreement with the reports of Bagali et al. (1999), Chandra and Das (2000), Panwar (2006), Raut et al. (2009), Reddy et al. (1997), Pandey et al. (2009) and Yadav et al. (2011).
Path-coefficient analysis is a tool to partition the observed correlation coefficient into direct and indirect effects of yield components on grain yield to provide clear picture of character associations for formulating efficient selection strategy. Path analysis differs from simple correlations in that it points out the causes and their relative importance, whereas the latter measures simply the mutual association ignoring the causation.
In the present study, the path-coefficient analysis was carried out at phenotypic as well as genotypic level. At phenotypic level, harvest-index exhibited very high positive direct effect on grain yield per plant, followed by considerable positive direct effect by days to maturity on grain yield per plant. Surek et al. (1998), Bagali et al. (1999), Meenakshi et al. (1999), Mishra and Verma (2002), Saif-ur-Rasheed et al. (2002), Madhavilatha et al. (2005) and Yadav et al. (2008) have also found high direct contribution of harvest-index on grain yield per plant in rice, Singh and Chaudhary (2006) and Kotal et al. (2010) in wheat, while Chaudhury and Das (1998) and Yadav et al. (2008) recorded high order positive direct effects of days to maturity on grain yield. Thus, harvest-index, followed by days to maturity emerged as most important direct contributor towards the grain yield. In contrast, biological yield per plant exerted very high negative direct effect on grain yield per plant, followed by substantial negative direct effect by days to 50% flowering at phenotypic level. Direct effects of remaining seven characters on grain yield per plant were too low to be considered important.
Indirect effects of days to 50% flowering, days to maturity, flag leaf area, effective tillers per plant, panicle length, spikelets per panicle and spikelet fertility on grain yield per plant were high order positive via harvest-index and high order negative via biological yield per plant. Similarly, plant height has considerable positive and negative indirect effect on grain yield per plant via biological yield per plant and harvest-index, respectively. Days to maturity showed considerable negative direct effect on grain yield per plant via days to 50% flowering. The above results indicates that the seven traits mentioned above in this paragraph showed contrasting high order positive or negative indirect effects on grain yield per plant at phenotypic level via one character or another. These seven traits can be considered as important but highly tricky indirect yield attributes whose handling may require very careful and balancing approach.
Very high order positive indirect effect of biological yield per plant and substantial positive indirect effect of 1000-grain weight on grain yield per plant via harvest-index were noted a phenotypic level. The estimates of indirect effects of these two traits via other characters were negligible but biological yield per plant had very high negative direct effect on grain yield per plant. Thus, biological yield per plant and 1000-grain weight may also be important indirect yield components. The complexity arising due to contrasting negative direct effect with positive indirect effect in case of biological yield per plant renders true role of biological yield per plant as indirect yield contributor difficult. On the basis of path-coefficient analysis at phenotypic level, the remaining estimates of indirect effects not discussed above were too low to be considered of any consequence.
The very high and positive direct effect of days to maturity has followed effective tillers per plant, 1000-grain weight, flag leaf area and panicle length to suggest that these five characters can be considered as important direct contributors of grain yield per plant. The high order positive direct contribution of days to maturity was also recorded by Chaudhury and Das (1998) and Yadav et al. (2008), while Madhavilatha et al. (2005), Khan et al. (2005), Agahi et al. (2007), Karad and Pol (2008) and Yadav et al. (2008) observed high order positive direct effects of effective tillers per plant on grain yield. Rather et al. (1997), Verma (1997), Mahto et al. (2003) and Agahi et al. (2007) reported high positive direct effects of 1000-grain weight on grain yield, while Reddy et al. (1997) noted high direct contribution to grain yield by fag leaf area. The high order positive direct effects of panicle length on grain yield have also been reported earlier by Chaudhury and Das (1998), Mishra and Verma (2002), Karad and Pol (2008) and Yadav et al. (2008). In contrast, very high negative direct effect of days to 50% flowering, followed by high order negative direct effects of biological yield per plant and spikelets per panicle were observed. The direct effects of remaining three characters were negligible.
Almost all the characters exhibiting high order positive indirect effects via one character showed high order negative indirect effects via another character. For example, days to 50% flowering, days to maturity, flag leaf area, effective tillers per plant, panicle length, spikelets per panicle and spikelet fertility showed high order positive indirect effects via harvest-index and high order negative indirect effects via biological yield per plant in path analysis at phenotypic level. Similarly, in genotypic path analysis, days to maturity, flag leaf area, effective tillers per plant, panicle length and 1000-grain weight showed high order positive direct effects on grain yield per plant but these traits had contrasting positive or negative indirect effects on grain yield per plant via days to maturity and days to 50% flowering.
CONCLUSION
Commencing the experimental findings it could be accomplished that most of characters were governed by additive gene action such as biological yield, grain yield, spikelets per panicle and flag leaf area, 1000-grain weight, harvest-index and plant height indicates that these traits are least influenced by environment hence; selection may be effective through these characters. Grain yield per plant exhibited a very strong positive association with biological yield per plant, flag leaf area and spikelet fertility at phenotypic and genotypic level which indicated that these traits were the strongest associates of grain yield per plant. However, significant negative correlation was observed in case of harvest-index with biological yield per plant and 1000-grain weight with effective tillers per plant. The genetic reasons for this type of negative association may be linkage or pleiotropy. Path analysis identified harvest-index, days to maturity and effective tillers per plant, 1000-grain weight and flag leaf area and panicle length as major direct contributors. Thus a genotype with higher magnitude of these traits could be either selected from existing genotypes or evolved by breeding program for genetic improvement of yield in rice. Further, the occurrence of negative as well as positive indirect effects by yield components on grain yield via one or other character simultaneously, presents a complex situation where a compromise is required to attain proper balance of different yield components in determining ideotype for high grain yield for salt affected soil in rice.
REFERENCES
- Agahi, K., M.H. Fotokian, and E.Farshadfar, 2007. Correlation and path coefficient analysis for some yield-related traits in rice genotypes (Oryza sativa L.). Asian J. Plant Sci., 6: 513-517.
CrossRef - Bagheri, N., N.B. Jelodar and A. Ghanbari, 2008. Diallel analysis study of yield and yield-related traits in rice genotypes. Int. J. Agric. Res., 3: 386-396.
CrossRefDirect Link - Bughio, H.R., M.A. Asad, I.A. Odhano, M.A. Arain and M.S. Bughio, 2009. Heritability, genetic advance and correlation studies of some important traits in rice. Int. J. Biol. Biotechnol., 6: 37-39.
Direct Link - Burton, W.G. and E.H. Devane, 1953. Estimating heritability in tall fescue (Festuca arundinacea) from replicated clonal material. Agron. J., 45: 478-481.
CrossRefDirect Link - Chandra, K. and A.K. Das, 2000. Correlation and interrelation of physiological parameters in rice under rainfed transplanted condition. Crop Res. Hisar, Assam Agric. Univ., 19: 251-254.
Direct Link - Dewey, D.R. and K.H. Lu, 1959. A correlation and path-coefficient analysis of components of crested wheatgrass seed production. Agron. J., 51: 515-518.
CrossRefDirect Link - Gregorio, G.B. and D. Senadhira, 1993. Genetic analysis of salinity tolerance in rice (Oryza sativa L.). Theor. Applied Genet., 86: 333-338.
CrossRefDirect Link - Johnson, H.W., H.F. Robinson and R.E. Comstock, 1955. Estimates of genetic and environmental variability in soybeans. Agron. J., 47: 314-318.
CrossRefDirect Link - Khan, A.J., F. Azam, A. Ali, M. Tariq and M. Amin, 2005. Inter-relationship and path coefficient analysis for biometric traits in drought tolerant wheat (Triticum aestivum L.). Asian J. Plant Sci., 4: 540-543.
CrossRefDirect Link - Kotal, B.D., A. Das and B.K. Choudhury, 2010. Genetic variability and association of characters in wheat (Triticum aestivum L.). Asian J. Crop Sci., 2: 155-160.
CrossRef - Kumar, S.T., R. Narasimman, R. Eswaran, C.P.S. Kumar and A. Anandan, 2007. Studies on genetic variability, heritability and genetic advance in segregating generations of rice (Oryza sativa L.). Int. J. Plant Sci., 2: 48-51.
Direct Link - Lush, J.L., 1949. Heritability of quantitative characters in farm animals. Hereditas, 35: 356-375.
CrossRefDirect Link - Manickavelu, A., R.P. Gnanamalar, N. Nadarajan and S.K. Ganesh, 2006. Genetic variability studies on different genetic populations of rice under drought condition. J. Plant Sci., 1: 332-339.
CrossRefDirect Link - Mode, C.J. and H.F. Robinson, 1959. Pleiotropism and the genetic variance and covariance. Biometrics, 15: 518-537.
Direct Link - Munns, R., 2002. Comparative physiology of salt and water stress. Plant Cell Environ., 25: 239-250.
CrossRefDirect Link - Pandey, P., P.J. Anurag, D.K. Tiwari, S.K. Yadav and B. Kumar, 2009. Genetic variability, diversity and association of quantitative traits with grain yield in rice (Oryza sativa L.). J. Biosci., 17: 77-82.
CrossRefDirect Link - Prasad, B., A.K. Patwary and P.S. Biswas, 2001. Genetic variability and selection criteria in fine rice (Oryza sativa L.). Pak. J. Biol. Sci., 4: 1188-1190.
CrossRefDirect Link - Saif-ur-Rasheed, M., H.A. Sadaqat and M. Babar, 2002. Inter-relationship among grain quality traits of rice (Oryza sativa L.). Asian J. Plant Sci., 1: 245-247.
Direct Link - Sarangi, D.N., B. Pradhan, P. Sial and C.H.P. Mishra, 2009. Genetic variability, correlation and path-coefficient analysis in early rice genotypes. Environ. Ecol., 27: 307-312.
Direct Link - Searle, S.R., 1961. Phenotypic, genetic and environmental correlations. Biometrics, 17: 474-480.
CrossRefDirect Link - Singh, L., D. Ram and B. Singh, 2008. Genetic variability and character association in paddy. Progr. Res., 3: 209-210.
Direct Link - Singh, G.P. and H.B. Chaudhary, 2006. Selection parameters and yield enhancement of wheat (Triticum aestivum L.) under different moisture stress conditions. Asian J. Plant Sci., 5: 894-898.
CrossRefDirect Link - Tester, M. and R. Davenport, 2003. Na+ tolerance and Na+ transport in higher plants. Ann. Bot., 91: 503-527.
CrossRefDirect Link - Tripathi, S.N., S. Marker, P. Pandey, K.K. Jaiswal and D.K. Tiwari 2011. Relationship between some morphological and physiological traits with grain yield in bread wheat (Triticum aestivum Lem. Thell.). Trends Applied Sci. Res., 6: 1037-1045.
CrossRefDirect Link - Wang, J.K. and P.H. Wolfgang, 2007. Simulation modeling in plant breeding: Principles and applications. Agric. Sci. China, 6: 908-921.
CrossRef - Yadav, S.K., P. Pandey, B. Kumar and B.G. Suresh, 2011. Genetic architecture, inter-relationship and selection criteria for yield improvement in rice (Oryza sativa L.). Pak. J. Biol. Sci., 14: 540-545.
CrossRef - Zafar, N., S. Aziz and S. Masood, 2006. Phenotypic divergence for agro-morphological traits among landrace genotypes of rice (Oryza sativa L.) from Pakistan. Int. J. Agric. Biol., 6: 335-339.
Direct Link