
Awale Degewione
Jimma University, P.O. Box 307, Jimma, Ethioppia
Sentayehu Alamerew
Somali Regional State Pastoral and Agro-pastoral Research Institute, P.O. Box 398, Jijiga, Ethiopia
Getachew Tabor
Debre-Zeit Agricultural Research Center, P.O. Box 32, Debre Zeit, Ethioia
International Journal of Agricultural Research
Year: 2011 | Volume: 6 | Issue: 7 | Page No.: 517-536







ABSTRACT
The aim of this study was to estimate the extent of genetic variability and character association among bulb yield and related traits. Forty nine shallot accessions from major shallot growing regions of were tested at Debre-Zeit Agricultural Research Center, Central Ethiopia from 2009 to 2010 in simple lattice design with two replications. Variances component method was used to estimate phenotypic and genotypic variation, heritability and genetic advance. Association of traits was also estimated using standard method. The accessions differed significantly for most of the characters and relatively wide range of the mean for most of characters indicated the existence of variation among the tested accessions. High Phenotypic Coefficient of Variation (PCV) and Genotypic Coefficient Variation (GCV) were recorded for leaf diameter and percentage of bulb sprouting. High GCV along with high heritability and genetic advance was obtained from leaf diameter and percentage of bulb sprouting. Bulb yield was positively and significantly associated with plant height, leaf length, leaf sheath length, leaf sheath diameter, bulb length, bulb diameter, bulb dry weight, biological yield per plant and marketable yield per plant at both phenotypic and genotypic levels. Genotypic path-coefficient analysis revealed that bulb dry weight exerted maximum positive direct effect on bulb yield and also exhibited positive association with bulb yield per plant, suggesting their possible utilization to improve bulb yield per plant. D2 analysis showed the 49 shallot accessions grouped into six clusters. This makes the accessions to become moderately divergent. Principal component analysis showed that the first six principal components explained about 76.15% of the total variation. Over all, the study confirmed the presence of character diversity in Ethiopian shallot accessions and this could be exploited in the genetic improvement of the crop through hybridization and simple selection.
PDF Abstract XML References Citation
Received: February 20, 2011;
Accepted: May 11, 2011;
Published: July 16, 2011
How to cite this article
Awale Degewione, Sentayehu Alamerew and Getachew Tabor, 2011. Genetic Variability and Association of Bulb Yield and Related Traits in Shallot (Allium cepa Var. Aggregatum DON.) In Ethiopia. International Journal of Agricultural Research, 6: 517-536.
DOI: 10.3923/ijar.2011.517.536
URL: https://scialert.net/abstract/?doi=ijar.2011.517.536
DOI: 10.3923/ijar.2011.517.536
URL: https://scialert.net/abstract/?doi=ijar.2011.517.536
INTRODUCTION
Shallot (Allium cepa var. aggregatum Don., 2n = 16) is originated in tropical central or western Asia and has been cultivated from very early period (Tindall, 1983). The Allium spp. are distributed widely through the temperate, warm temperate and boreal zones of the northern hemisphere (Brewster, 2008). Typically, alliums are plants of open, sunny, dry sites in fairly arid climates (Brewster, 2008). As shallot and its relative species are generally open pollinated crops and have been cultivated for long time, a number of landraces and natural hybrids either intraspecific or interspecific probably are to be on the increase (Arifin and Okubo, 1996).
Tropical and sub-tropical shallots are preferred for their tolerance to the hot and humid tropical climate, better tolerance to pests and diseases and longer storage life than common onion (Rabinowitch and Kamenetsky, 2002). Locally adapted cultivars are grown either for their special flavor, green leaves or curative effects in Europe, USA and Asia (Fritsch and Friesen, 2002), in France (Brewster, 2008) and in Malaysia, Singapore and Indonesia (Susheela, 2007). Per capita consumption of shallot is 1.7 kg in rural areas and 5.9 kg in the towns (Currah and Proctor, 1990).
Shallot is one of the major vegetable crops used as condiments in most Ethiopian cuisines. It is hardly possible to get a dish without this vegetable in every meal of a day (Currah and Proctor, 1990). It has long been growing in Ethiopia by subsistent farmers in the mid and high altitudes (1800 m.a.s.l to 2200 m.a.s.l) for flavoring of local foods and as a source of cash (Getachew et al., 2009). Although shallot has similar agro-ecological requirements with common onion it is better adapted to rain-fed (short growing seasons) and is relatively tolerant to leaf diseases (Currah and Proctor, 1990).
The estimated area under production of shallot and onion in the country in the 2007/2008 cropping season was 1803 ha with total production of 1751 tones of fresh bulbs (CSA, 2008). Despite its high economic importance, the yield of shallot under farmers’ conditions is very low (6 t h-1) compared to the 25 t ha-1 obtained under good management practices (Getachew et al., 2009). The wide gap in yield is attributed to lack of improved varieties, poor agronomic practices and soil fertility and diseases (bulb rot and downy mildew) and insect pests (onion thrips), etc in farmers’ fields (Getachew and Asfaw, 2000). Shallot grown in Ethiopia is diverse and includes: bolters and non-bolters, spreading and compact types, those with various bulb shapes, sizes and colors (Getachew and Asfaw, 2000). Likewise, several researchers (Singh, 1981; Kalloo et al., 1982; Barta et al., 1983; Abayneh, 2001) reported the existence of wide phenotypic and genotypic variations in vegetative, bulb yield and yield components and quality in onion. Mohanty (2001) recorded moderate to high estimates of heritability, genetic coefficients of variation and genetic gain for weight of bulb and number of leaves per plant which could be improved by simple selection (Bose et al., 2003).
In Allium spp., high estimates of heritability and genetic advance with respect to bulb weight, leaf length, leaf number, bulb length, bulb diameter have been reported (Kalloo et al., 1982; Dowker, 1990; Abayneh, 2001; Mohanty, 2001). Bulb diameter, bulb weight, bulb thickness, leaf length and days to flowering were found to be highly heritable in onion (Singh, 1981). Owen (1961) also reported the involvement of cumulative gene action and a fewer number of genes in controlling soluble solids in onion. Warid (1952) also found high heritability and the involvement of four to ten gene pairs and partial dominance of low soluble solids in onions.
In Allium cepa, positive correlations were observed between yield and leaf length (Dowker et al., 1976) and between neck thickness and bulb diameter (Patil and Kale, 1985). Mohanty, 2001) also reported significant and positive phenotypic and genotypic associations of bulb yield with plant high, number of leaves per plant, bulb diameter in onion but significant and negative correlation with neck thickness (Bose et al., 2003).
The knowledge of the extent of genetic variability present in the population is essential for further improvement of shallot. Similarly, information on the extent and nature of interrelationship among characters help in formulating efficient scheme of multiple trait selection, as it provides means of direct and indirect selection of component characters (Singh, 2006). Although, large numbers of shallot accessions are collected from major growing regions of Ethiopia by Deber-Zeit Agricultural Research Center (DZARC), research on variability and association among characters in these accessions are limited. Hence, the present study was undertaken with objectives to estimate the extent of variability for bulb yield and other related characters and the extent of correlation between pairs of characters at phenotypic and genotypic levels and thereby compare the direct and indirect effects of the characters on yield.
MATERIALS AND METHODS
Experimental site: The study was conducted at Debre-Zeit Agricultural Research Center (DZARC) in the central part of Ethiopia from 2009 to 2010 dry season using irrigation. DZARC is located at 38°55”N latitude, 8°44”E longitude and 1900 m.a.s.l, 45 km from Addis Ababa, Ethiopia. It receives an average rainfall of 851 mm per year with mean annual maximum and minimum temperature of 24.3°c and 8.9°c, respectively (DZARC, 1991). It has Vertisol soil.
Experimental materials: A total of forty-nine different shallot bulb accessions that include one local check and one standard check were used for the study. The majority of the accessions represent the national collection from different shallot growing regions of the country and that are maintained at DZARC (Table 1).
| Table 1: | Shallot accessions used in the study |
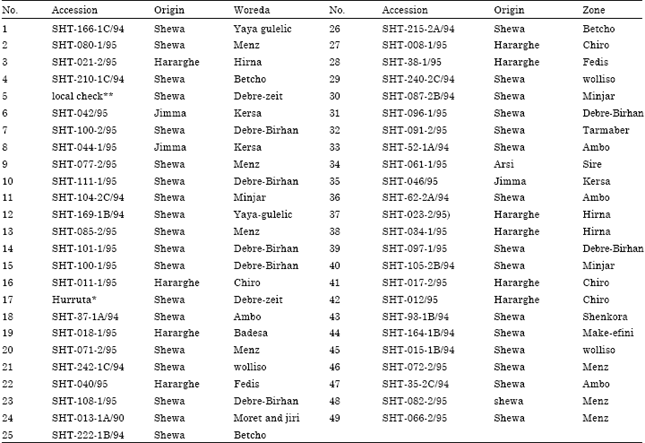 | |
| Source: Debre-Zeit Agricultural Research Center, *standard check, **Local check | |
Experimental design: The experiment was laid out in a 7x7 simple lattice design with two replications. Healthy and clean bulbs of each genotype were selected and planted on well prepared plots. The plot size was 2 m long and 1.8 m wide, consisting of 6 rows per plot. The bulbs were planted at spacing of 30 cmx20 cm between rows and plants, respectively. Phosphorus was applied in the form of Di-Amonium Phosphate (DAP) at the rate of 200 kg ha-1 at planting. Nitrogen was applied in the form of urea at the rate of 150 kg ha-1, in two splits, half at planting and the remaining half top-dressed 45 days after planting. All the other management practices were uniformly applied to all plots using recommended practices of DZARC (Getachew et al., 2009).
Data collected: Twenty two quantitative characters were recorded on eight randomly selected plants from the four middle rows of each plot by adopting IPGRI descriptor (IPGRI, 2001).
Data analysis: Data for quantitative characters were subjected to analysis of variance (ANOVA) for simple lattice design using Proc lattice and Proc GLM procedures of SAS version 9.2, (SAS Institute Inc., 2008). The difference between treatments means was compared using LSD at 1% and 5% probability levels.
Variability among the accessions was estimated using genotypic variances and coefficients of variations as suggested by Burton and de Vane (1953) as:
where, r is No. of replication, Msg is mean square due to genotypes (accessions), MSe is mean square of error (Environmental variance), Environmental variance (σ2e) is error mean square (Mse), Phenotypic variance (σ2p) is σ2g + MSe where, σ2g is genotypic variance and Phenotypic coefficient of variation (PCV).
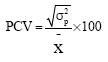 |
X is Mean of the character.
Genotypic coefficient of variation (GCV):
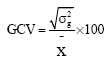 |
Broad sense heritability (H) for quantitative characters was computed using the formula suggested by Singh and Chaudhury, 1985) as follows:
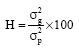 |
Genetic Advance (GA) for selection intensity (K) at 5% was computed according to Allard (1960) as given here under.
where, K is a constant (k = 2.056 at 5% selection intensity) and σp is phenotypic standard deviation on mean basis.
Character associations at genotypic and phenotypic levels were calculated from the genotypic, phenotypic and environmental covariances according to Miller et al. (1958). In path analysis, bulb yield per plant was taken as the resultant (dependent) variable while the rest of the characters were considered as casual (independent) variables. The direct and indirect effects of the independent characters on bulb yield per plant were estimated by the simultaneous solution of the formula suggested by Dewey and Lu (1959) and with statistical package developed by Doshi (1991).
RESULTS AND DISCUSSION
Analysis of variance: The analysis of variance (Table 2) for all of the characters studied, except number of leaves per plant and days to maturity which were significant (p<0.05), revealed highly significant (p<0.01) differences among the shallot accessions, indicating the existence of sufficient genetic variability among the accessions. Similarly, highly significant variations were observed in plant height, leaf length, bulb diameter, bulb length, bulb dry weight and biological yield per plant in onion (Abayneh, 2001); in number of leaves and leaf width in Iranian garlic (Baghalian et al., 2006) and in plant height, leaf length, bulb length and bulb diameter in garlic (Figliuolo et al., 2001). Similar to our findings, Dar and Sharma (2011) obtained highly significant difference among the genotypes for all the quality traits studied in tomatoes. Moreover, Alsemaan et al. (2011) reported the existence of genetic diversity within Rosa damascena cultivated in Syria which is in support of the present finding. In addition, Parthiban et al. (2011) found variability in Jatropha for some character studied which is in support of our results. Furthermore, Singh et al. (2011) also reported similar finding in field pea. In contart to our finding, Gichimu and Omondi (2010) studied on morphological characterization of five newly developed lines of Arabica coffee and two commercial cultivars in Kenya and they reported non significant difference for internodes length.
Mean performance of accessions: Negelle had the largest bulb (250.74 g/plant) while DZSHT35-2C/94 had the smallest bulb (140.84 g/plant) (Appendix 1). The highest bulb yield (250.74 g) per plant was also recorded in Negelle due to its large sized bulbs. Accession with the shortest plant height DZSHT072-2/95 (40.98 cm) had lower bulb yield per plant (144.96 g) while tall accessions Hurruta (51.60 cm), Negelle (51.12 cm), DZSHT164-1/94 (51.26 cm), DZSHT097-1/95 (53.13 cm), DZSHT061-1/95 (53.83 cm) and DZSHT082-2/95 (53.74 cm) had higher yield bulb per plant of 245.94 g, 250.74 g, 244.88 g, 221.28 g,244.97 g, 236.73 g, respectively.
The accessions also showed a difference of one month in days to maturity. Result in Appendix 2 showed that 40.82% of the accessions were early maturing (92-95 days), 36.73% medium maturing (96-100 days) and only 22.45% late maturing (101-118 days). The result is in agreement with the observation of Getachew and Asfaw (2000) indicating that the earliest shallot cultivars matured in 95 days while the latest took about 126 days.
| Table 2: | Analysis of variance for the 22 characters in shallot accessions tested at DZARC (2009/10) using simple lattice design |
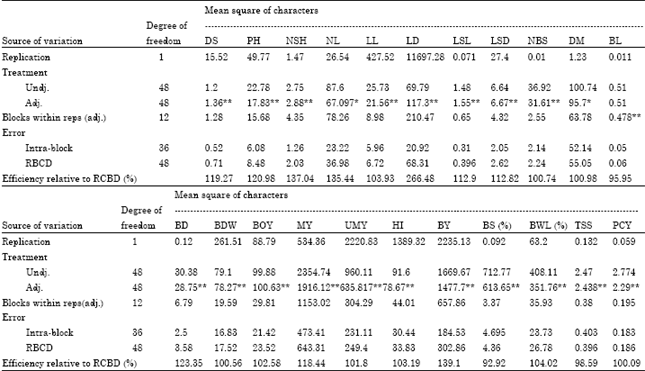 | |
| **and * indicates significant at 1% and 5% probability levels, respectively. BD: Bulb diameter, DBW: Bulb dry weight, BOY: Biological yield per plant, MY: Mmarketable yield per plant, UMY: Unmarketable yield per plant, HI: Harvest index, BY: Bulb yield per plant, BS (%): Percentage of bulb sprouting, BWL (% ): Percentage of bulb weight, TSS: Total soluble solid and PCY: Pungency. **and *indicates significant at 1% and 5% probability levels, respectively. DS: Days to sprouting, PH: Plant height, NSH: Number of shoots per plant, NL: Number of leaves per plant, LL: Leaf length, LD: Leaf diameter, LSL: Leaf sheath length, LSD: Leaf sheath diameter, NBS: Number of bulb splits per plant, DM: Days to maturity and BL: Bulb length | |
Variabilities in days to maturity and other traits were also reported earlier by Rabinowitch (1988) in onion.
Late maturing accessions (Hurruta, Negelle, DZSHT100-1/95 and DZSHT061-1/95) have high yield per plant compared to the early maturing ones (DZSHT164-2B/94), in agreement with the observations of Boswell (1984). The variation may offer an opportunity to develop varieties for agro-ecologies that differ in their moisture patterns. Among the forty nine accessions, DZSHT169-1B/94, DZSHT164-1B/94, DZSHT091-2/95, DZSHT008-1/95 and DZSHT082-2/95 were found to be high yielding as well as early maturating.
Cognizant of the crucial importance of shelflife to ensure an extended availabilityof bulbs, the accessions were stored in a diffused light store at an average minimum and maximum temperature of 10.76°C and 27.85°C and relative humidity of 47.01% for 12 weeks. A significant variation was observed among the accessions: DZSHT164-1B/94, an early maturing (95 days) and high yielder (244.88 g) accession, also had low percentage of bulb weight loss (43.75 percent) and bulb sprouting (8 percent) (Appendix 2). Similarly, Galvan et al. (1997) obtained differential responses of onion cultivars in their storability. Similarly, Brewster, 2008) reported that some shallot strains probably represent the extreme of storability, while the delicate green shoots of Chinese chives can only be stored for 2-3 days.
On the other hand, high bulb weight loss (77.99 percent) and bulb sprouting (49 percent) coupled with high bulb yield per plant (244.97 g) was recorded in DZSHT061-1/95 indicating that such high yielding varieties may only be produced if they can be consumed immediately. The utilization of their high yielding potential needs to bring in low sprouting and weight loss traits from accession such as DZSHT164-1B/94 and DZSHT015-1B/94. The high bulb weight loss and percentage of bulb sprouting record in DZSHT077-2/94, DZSHT072-2/95, DZSHT017-2/95, DZSHT101-1/95 and DZSHT104-2C/94 indicated that direct selection based on yield traits could lead to is inferior varieties with short shelflife (Appendix 2).
In general high yielding accessions such as Negelle, Hurruta, DZSHT100-1/95, DZSHT061-1/95 and DZSHT011-1/95 had high bulb weight loss ranging from 63.67 to 77.99% in storage. On the other hand, the low yielding shallot accessions DZSHT040/95 and DZSHT053-1A/90 were among the accessions with long shelflife (only about 40% weight loss). Hence, breeders should combine high yieldand long shelflife to develop cultivars that benefit farmers, traders and consumers.
The pyruvate level estimated in the shallot accessions ranged between 7.37 and 12.49 μ molg-1 of fresh weight in DZSHT046/95 and DZSHT101-1/95, respectively (Table 6 and Appendix 2). Similar levels were reported by Baghalian et al., 2006) in garlic. Low, medium and high pungent onions with0-3, 3-7 and above 7 μ mol-1 pyruvic acid per gram of fresh weight, respectively, were reported by Shock et al. (2004). But almost all of the shallot accessions investigated in the present study were qualified as highly pungent. The lowest and the highest levels of total soluble sugars were recorded in DZSHT35-2/94 (10.6%) and in DZSHT222-1B/94 (16.4%) Appendix 1).
Balance between pyruvate (pungency) and Total Soluble Sugars (TSS) determines the preference of consumers. Shallot accessions with high levels of sugars and pyruvate were DZSHT044-1/95, DZSHT222-1B/94 and DZSHT087-2B/94. However, DZSHT164-1B/94 was found to have low level of pyruvate but high sugar content, qualifying a character desired in sweet onions.
Bulb dry weight also showed wide variation among the accessions ranging between 10.37 g in DZSHT034-1/95 and 33.93g in DZSHT097-1/95. Marketable yield per plant also varied from 73.57 g in DZSHT080-1/95 to 216.74 g in Hurruta indicating that Hurruta can be used for further breeding to improve marketability of bulbs (Appendix 1) (Table 6, Appendix 2). In general, the range and the mean in this study suggested the existence of wide variations among the shallot accessions for all of the characters studied and their considerable potential in the improvement of shallot.
Phenotypic and genotypic variations: We recorded phenotypic variances ranging between 0.271 and 1193.765 and genotypic variances ranging between 0.42 and 721.335 for traits considered in this study (Table 3). Relatively higher phenotypic variance values of 1193.765 for marketable yield and 831.115 for bulb yield per plant were obtained in our study. Similarly, the genotypic variances for these characters were almost as high, indicating that the genotype could be reflected by the phenotype and the effectiveness of selection based on the phenotypic performance for these characters. This corroborates with the findings of Hosamani et al. (2010) considering bulb yield only for both the variances.
Phenotypic Coefficients of Variation (PCV) ranged from 7.23% for plant height to 68.32% for percentage bulb sprouting. The Genotypic Coefficients of Variation (GCV) ranged from 4.66% in days to maturity to 67.13% in percentage bulb sprouting (Table 3). Deshmukh et al. (1986) classified PCV and GCV values as high (>20%), medium (10-20%) and low (<10%). Accordingly, high PCV and GCV were observed in characters like leaf diameter, number of bulb splits per plant, biological yield per plant, marketable yield per plant, unmarketable yield per plant, bulb dry weight, bulb weight loss and bulb sprouting. The high PCV and GCV indicated that selection may be effective based on these characters and their phenotypic expression would be good indication of the genotypic potential (Singh et al., 1994).
| Table 3: | Estimate of ranges, mean, phenotypic (PV) and genotypic (GV) component of variances, broad sense heritability, and genetic advance as percent of mean of 22 characters of shallot accessions at DZARC (2009/10) |
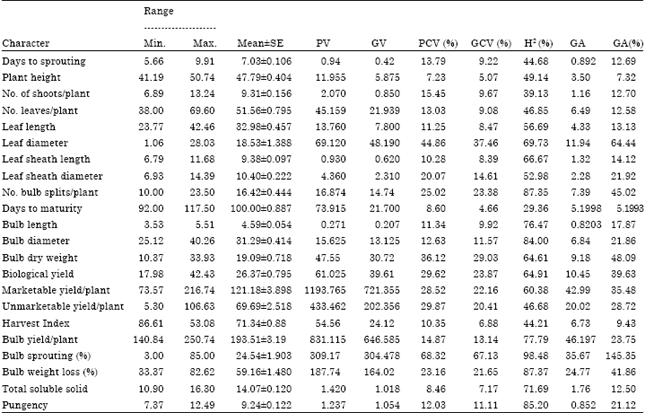 | |
| PV: Phenotypic variance, GV: Genotypic variance, H2 (%): Broad sense heritability, GCV (%): Genotypic coefficient of variation, PCV (%): Phenotypic coefficient of variation, GA (%): Genetic advance, GAM: Genetic advance as percent of mean | |
Similarly, Kassahun (2006) reported high PCV and GCV estimates for bulb dry weight, weight of bulb and biological yield per plant in garlic. It is also in conformity with the findings of Getachew and Asfaw (2000) in shallot. Medium PCV and GCV were displayed in bulb diameter, leaf sheath diameter, bulb yield per plant, number of shoots per plant, bulb length and pungency. Further more, medium and low PCV and GCV, respectively, were observed in characters like leaf length, harvest index, number leaves per plant and days to sprouting, agreeing with findings of Singh (1981) in onion.
On the other hand, plant height, total soluble solid and days to maturity showed low GCV and PCV, indicating less scope of selection as they are under the influence of environment.
Phenotypic coefficients of variation were found to be higher than genotypic coefficients of variation for all characters studied (Table 3). Similar results were reported by Melke and Ravishankar, (2006) in twenty six onion accessions and by Pramoda and Gangaprasad (2007) in four F2 onion populations. In most cases, the two values differ slightly indicating less influence of environmental factors. Results of the present study concur with that of Korla et al. (1981) and Kassahun (2006) in garlic and Abayneh (2001) and Hosamani et al. (2010) in onion. Wide differences between PCV and GCV values were observed in leaf diameter, bulb dry weight, biological yield per plant, marketable yield per plant and unmarketable yield per plant which may indicate significant influence of environmental factors on these traits. Therefore, it would be appropriate to consider the above characters depending on the objective of shallot improvement program.
Estimates of heritability (H2) in the broad sense: he values of estimated broad sense heritability for the studied characters were found between 29.36% for days to maturity to 98.48% for bulb sprouting (Table 3). According to Pramoda and Gangaprasad (2007) heritability estimates can be low (<40%), medium (40-59%), moderately high (60-79%) and very high (= 0%). Heritability estimates were very high for percentage bulb sprouting (98.48%), percentage of bulb weight loss (87.37%), number of bulb splits per plant (87.30%), pungency (85.20%) and bulb diameter (84.00%), indicating the possibility of success in selection. The very high heritability estimates of pungency obtained in the present study is in agreement with that of Fasika et al. (2008). Total soluble solids (71.69%), bulb yield per plant (77.79%), biological yield per plant (64.91%), bulb dry weight (64.9 1%), (64.61%), marketable yield per plant (60.38%), leaf diameter (69.73%), leaf sheath length (66.67%) and bulb length (76.47%) exhibited moderately high heritability estimates. Abayneh (2001) observed similar results in biological yield and total soluble solids in onion. These characters, therefore, may respond effectively to selection pressure.
Moderate heritability estimates were observed for unmarketable yield per plant (46.68%), days to sprouting (44.68%), number of leaves per plant (46.85%), harvest index (44.21%), leaf length (56.69%), leaf sheath diameter (52.98%) and plant height (49.14%). On the other hand, low heritability estimates were also observed for days to maturity (29.36%) and number of shoots per plant (39.13%) indicating the limited scope for improvement of these characters through selection. Similarly, Pike (1986), Abayneh (2001), Mohanty (2001) and Fasika et al. (2008) observed moderate to high heritability estimates for bulb yield per plant in onion.
Estimates of expected genetic advance (GA): The expected genetic advance expressed as a percentage of the mean by selecting the top 5% (high yielder) of the accessions, varied between 5.20% for days to maturity and 145.35% for percentage of bulb sprouting (Table 3) indicating that selecting the top 5% of the base population could result in an advance of 5.20 to 145.35 percent over the respective population mean.
Genetic advance as percentage of mean was maximum for percentage of bulb sprouting followed by leaf diameter, bulb dry weight, number of bulb splits per plant and percentage of bulb weight loss and biological yield per plant. Likewise, genetic advance was maximum for bulb yield per plant, marketable yield per plant, percentage of bulb weight loss, percentage of bulb sprouting and unmarketable yield per plant. This is in close agreement with findings of Kassahun (2006) in garlic.
Burton (1952) suggested that genetic coefficient of variation together with heritability estimate would give the best picture of the amount genetic advance to be expected from selection. Similarly, Johnson et al. (1955) and Johnson and Hernandez (1980) reported that high genotypic coefficients of variation along with high heritability and high genetic advance provide better information than each parameter alone.
High genetic coefficient of variation, heritability and genetic advance were found in biological yield per plant, bulb dry weight, percentage of bulb sprouting, leaf diameter, number of bulb splits per plant and percentage of bulb weight loss. These characters could be useful basis of selection. Sandhu and Korla (1976) and Melke and Ravishankar (2006) obtained similar results in percentage of bulb sprouting. Besides, Jalata et al. (2011) also reported that high heritability coupled with high genetic advance was recorded in thousand kernel weight in Ethiopian barley.
Association of characters: Estimates of phenotypic and genotypic correlation coefficients between each pair of characters are presented in Tables 4 and 5. The results showed that, in most cases, the genotypic correlation coefficients were higher than the phenotypic correlation coefficients which indicated the inherent association among various characters independent of environmental influence. The results are consistent with the reports of Kalloo et al. (1982) and Kassahun (2006) in garlic, Shimeles (2000), Abayneh (2001) and Hosamani et al. (2010) in onion.
Based on phenotypic and genotypic correlation coefficients, bulb yield per plant showed positive and significant association with plant height, leaf length, leaf sheath length, leaf sheath diameter, bulb length, bulb diameter, bulb dry weight, biological yield per plant and marketable yield per plant at phenotypic and genotypic levels. that the results imply that improvement of these characters could improve the capacity of the plants to synthesize and translocate photosynthates to the organ of economic value, the bulb Similar findings were reported by Vavidel et al. (1981) and Pandian and Muthukrishnan (1982) in shallot, Mulungu et al. (1998), Kalloo et al. (1982), Mahantesh et al. (2007) and Abayneh (2001) in onion, Lee et al. (1977) in garlic, Nikhila et al. (2008) in robusta coffee, Umamaheswari and Mohanan (2011) in vanilla and Akansha Singh, et al. (2011) in field pea considering plant height only. Contrary to results of the present study, Badshah and Umar (1999) reported negative correlation between yield and plant height in garlic.
Bulb yield per plant had positive and significant associations with leaf diameter and total soluble solids at phenotypic level. The same trait also displayed non-significant and positive association with the same traits at genotypic level, in contrast with the findings of Darbyshire and Henry (1981) who reported negative and significant correlation of bulb yield per plant with total soluble solids in onion. Bulb yield per plant displayed negative and significant association with harvest index, percentage of bulb sprouting, percentage of bulb weight loss and number of bulb splits per plant at genotypic level, indicating the difficulty in simultaneous improvement of these traits.
Bulb dry weight had positive and significant association with biological yield per plant, marketable yield per plant, percentage of bulb sprouting and percentage of bulb weight loss, plant height, leaf length, leaf sheath length, leaf sheath diameter, bulb length and bulb diameter at phenotypic and genotypic levels. An increase in these characters hastened the production of dry matter in the shallot accessions, in agreement with the findings of Abayneh (2001) and Havey (1993) in onion. On the other hand, bulb dry weight had negative and significant association with number of leaves per plant, number of bulb splits per plant and harvest index at phonotypic level, in conformity with the results of Abayneh (2001) and Havey (1993) in onion.
Total soluble solids had positive and significant association with leaf sheath, leaf length and leaf sheath diameter at both geneotypic and phenotypic levels but had similar association with number of shoots per plant only at phenotypic level. However, its association with percentage of bulb weight loss at phenotypic level was negative and significant. Bulb dry weight had negative and significant association with number of leaves per plant, number of bulb splits per plant, unmarketable yield per plant and harvest index at genotypic level suggesting that an increase in bulb dry weight could result from decrease in all these traits. In contrast, total soluble solids had positive and significant association with plant height, number of leaves per plant, leaf length, marketable yield per plant, number of bulb splits per plant and harvest index. However, it had negative and significant association with percentage of bulb sprouting indicating the practical difficulty of simultaneous improvement of both traits due to lack of closely linked genes that cause co-variation in the traits (Falconer, 1989). This was not in line with the work of Nikhila et al. (2008) who reported that length of primary branches , number of primary branches, intermodal length and bush spread are the character that should be given premium importance while carrying out crop improvement programmes in robusta coffee.
| Table 4: | phenotypic correlation coefficients (Pr) of the 22 quantitative characters of shallot accessions at DZARC (2009/10) |
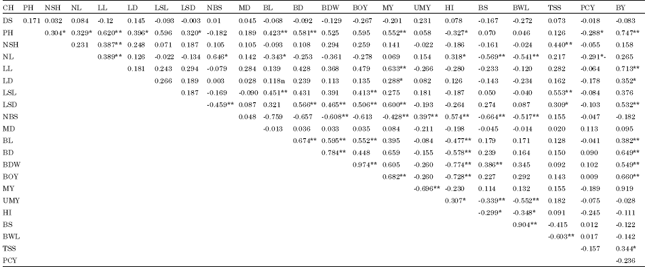 | |
| *and** indicated significant at p = 0.05 and p = 0.01probability level; respectively DS: Date to sprouting; PH: Plant height; NSH: No. of shoots per plant; NL: No. of leaves per plant; LL: Leaf length; LD: Leaf diameter; LSL: Leaf sheath length BL: Bulb length BD: Bulb diameter BOY: Biological yield above ground per plant; BDW: Bulb dry weight per plant; HI: Harvest index per plant ; UMY: Unmarketable yield per plant; Y: Marketable yield per plant; NBS: No. of bulb splits per plant; LSD: Leaf sheath diameter; MD: Days of maturity; BY: Bulb yield per plant ; BS(%): Percentage of sprouting bulbs; BWL(%): Percentage of bulb weight loss; TSS: Total soluble solid and PCY: Pungency | |
| Table 5: | Genetic correlation coefficients (g ) of the 22 quantitative characters of shallot accessions at DZARC (2009/2010) |
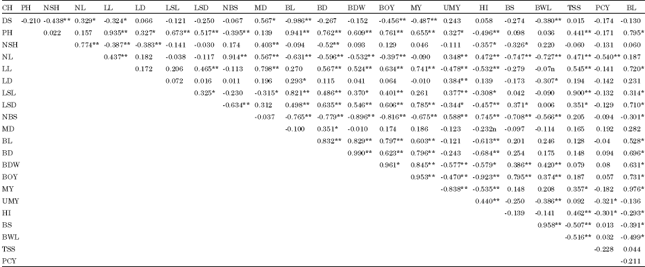 | |
| DS: Date to 50% sprouting PH: Plant height; NSH: No. of shoots per plant; NLL No. of leaves per plant; LL: Leaf length; LD: Leaf diameter; LSL: Leaf sheath length BL: Bulb length BD: Bulb diameter BOY: Biological yield above ground per plant; BDW: Bulb dry weight per plant; HI: Harvest index per plant; UMY: Unmarketable yield per plant; MY: Marketable yield per plant ; NBS: No. of bulb splits per plant; LSD: Leaf sheath diameter; MD: Days of maturity; BY: Bulb yield per plant; BS(%): Percentage of sprouting bulbs; BWL(%): Percentage of bulb weight loss; TSS: Total soluble solid and PCY: Pungency | |
| Table 6: | Estimate of direct effect (bold face and diagonal ) and indirect effects (off diagonal) at genotypic level in 49 shallot accessions tested at DZARC (2009/10) |
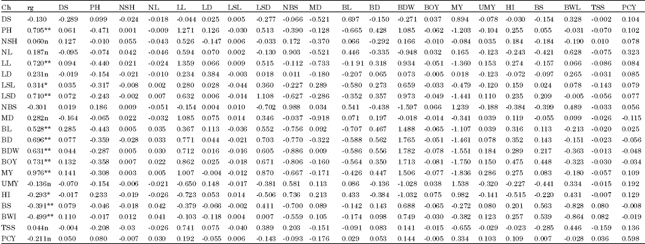 | |
| rg: Genotypic direct effect residual effect = 0.4320, Ch: Character, DS: Date to sprouting, PH: Plant height, NSH: No. of shoots per plant, NL: No. of leaves per plant, LL: Leaf length, LD: Leaf diameter, LSL: Leaf sheath length, BL: Bulb length, BD: Bulb diameter, BOY: Biological yield above ground per plant, BDW: Bulb dry weight per plant, HI: Harvest index per plant, UMY: Unmarketable yield per plant, Y: Marketable yield per plant, NBS: Number of bulb splits per plant, LSD: Leaf sheath diameter, MD: Days of maturity, BY: Bulb yield per plant, BS(%): Percentage of sprouting bulbs, BWL(%): Percentage of bulb weight loss, TSSL: Total soluble solid and PCY: Pungency | |
Path coefficient analysis: Results in Table 6 showed path coefficient analysis of all traits on bulb yield per plant. High magnitude and maximum positive direct effects on bulb yield was exerted by bulb dry weight (1.782) followed by leaf length (1.359), leaf sheath diameter (1.108), number of bulb splits per plant (0.988), pungency (0.598) and percentage of sprouting bulbs(0.563) and bulb diameter (0.562), number of bulb splits per plant, indicating the true relationship between these traits as good contributors to bulb yield in agreement with findings of Hyder et al. (2007) in shallot.
Marketable yield per plant had maximum negative direct effect (-1.836) followed by days to maturity (-0.918), percentage of bulb weight loss (-0.864), bulb length (-0.707), pungency (-0.598) and harvest index (-0.515). Marketable yield per plant and bulb length showed the first and fourth (in that order) maximum negative direct effects on bulb yield per plant in spite of their high positive and significant association with bulb yield per plant. The positive indirect influence of marketable yield per plant through days to sprouting, number shoots per plant, number leaves per plant, leaf length, leaf sheath diameter, bulb diameter, bulb dry weight, unmarketable yield per plant, harvest index, percentage of bulb sprouting and pungency and indirect favorable effect of bulb length on bulb yield per plant via leaf length, leaf sheath diameter, bulb diameter and bulb dry weight which was counterbalanced by their unfavorable indirect effects leading to strong positive association (Table 6). Therefore, these traits must be considered if selection is to be made through marketable yield per plant and bulb length in agreement with findings of Rahman and Das (1985) in onion and of Hyder et al. (2007) in shallot. On the other hand, plant height, leaf sheath length and biological yield per plant displayed low and negative direct effects on bulb yield per plant. In addition, biological yield per plant had unfavorable indirect effects on bulb yield per plant through plant height, number bulb splits per plant, bulb length and marketable yield per plant. This unfavorable indirect effect was counterbalanced by the favorable indirect effect of leaf length, leaf sheath diameter, bulb diameter, bulb dry weight, unmarketable yield per plant and harvest index and the indirect effect of plant height via leaf length, leaf sheath diameter, bulb dry weight was outweighed by the unfavorable indirect influence leading to strong positive association, in agreement with the findings of Mohanty (2001) and Mahantesh et al. (2007) in onion. Similarly, leaf sheath length showed highly favorable indirect effects on bulb yield via leaf sheath diameter and bulb dry weight which was highly nullified by bulb length and marketable yield per plant leading to its weak positive correlation with bulb yield.
Bulb dry weight which showed maximum direct effect also exerted considerable positive indirect effect via bulb diameter and negative indirect effect through number of bulb splits per plant, indicating the need for attention while selecting these characters. In addition, favorable indirect effect on bulb yield per plant was obtained for leaf sheath diameter via bulb dry weight. Number of bulb splits in addition to exerting positive direct effect on bulb yield per plant also displayed favorable indirect effect via marketable yield. Percentage of bulb sprouting and bulb diameter having positive direct effects also revealed positive indirect effects on bulb yield through bulb dry weight.
ACKNOWLEDGMENT
The authors wish to thank Ethiopian Institute of Agricultural Research for funding the research.
APPENDIX
| Appendix 1: | Mean performance of 49 shallot Accessions for 22 quantitative characters |
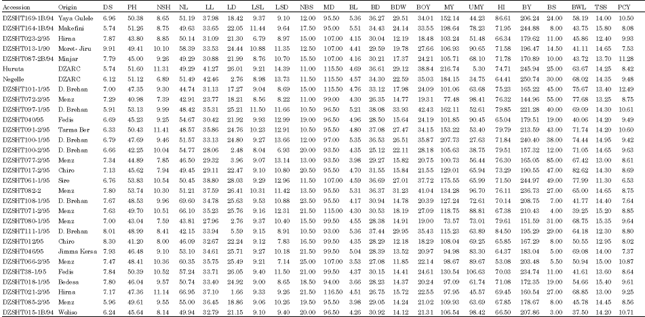 | |
| DS: Date to 50% sprouting PH: Plant height; NSH: No. of shoots per plant; NL: Number of leaves per plant; LL: Leaf length; LD: Leaf diameter; LSL: Leaf sheath length BL: Bulb length BD: Bulb diameter BOY: Biological yield above ground per plant; BDW: Bulb dry weight per plant; HI: Harvest index per plant; UMY: Unmarketable yield per plant; MY: Marketable yield per plant; NBS: Number of bulb splits per plant; LSD: Leaf sheath diameter ; MD: Days of maturity; BY: Bulb yield per plant; BS(%): Percentage of sprouting bulbs; BWL(%): Percentage of bulb weight loss; TSS: Total soluble solid and PCY: Pungency | |
| Appendix 2: | Mean performance of next 19 shallot Accessions for 22 quantitative characters |
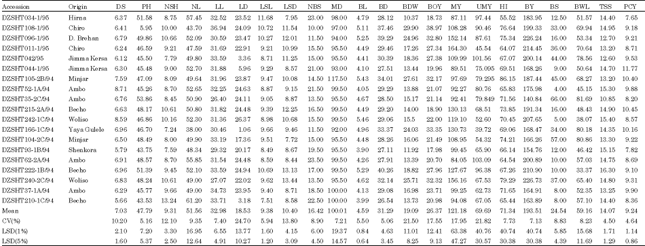 | |
| CV: Coefficient of variation; LSD: Least significant difference; DS: Date to 50% sprout PH: Plant height NSH: No. of shoots per plant; NL: No. of leaves per plant; LL: Leaf length; LD: Leaf diameter; LSL: Leaf sheath length BL: Bulb length BD: Bulb diameter BOY: Biological yield above ground per plant; BDW: Bulb dry weight per plant ;HI: Harvest index per plant; UMY: Unmarketable yield per plant; MY: Marketable yield per plant ; NBS: No. of bulb splits per plant; LSD: Leaf sheath diameter ;MD: Days of maturity; BY: Bulb yield per plant ;BS(%): Percentage of sprouting bulbs; BWL(%): Percentage of bulb weight loss ;TSS: Total soluble solid and PCY: Pungency | |
REFERENCES
- Singh, A., S. Singh and J.D.P. Babu, 2011. Heritability, character association and path analysis studies in early segregating population of field pea (Pisum sativum L. var. arvense). Int. J. Plant Breed. Genet., 5: 86-92.
CrossRefDirect Link - Allard, R.W., 1960. Principles of Plant Breeding. 1st Edn., John Wiley and Sons Inc., New York pp: 372-372.
CrossRefDirect Link - Alsemaan, T., N. Albatal, H. Baydar and K. Almaarri, 2011. Genetic diversity and qualitative variation of Rosa damascene in Syria. Int. J. Agric. Res., 6: 429-436.
CrossRefDirect Link - Baghalian, K., M.R. Naghavi, S.A. Ziai and H.N. Badi, 2006. Post-planting evaluation of morphological characters and allicin content in Iranian garlic (Allium sativum L.) ecotypes. Sci. Hortic., 107: 405-410.
CrossRef - Dar, R.A. and J.P. Sharma, 2011. Genetic variability studies of yield and quality traits in tomato (Solanum lycopersicum L.). Int. J. Plant Breed. Genet., 5: 168-174.
CrossRef - Burton, W.G. and E.H. Devane, 1953. Estimating heritability in tall fescue (Festuca arundinacea) from replicated clonal material. Agron. J., 45: 478-481.
CrossRefDirect Link - Darbyshire, B. and R.J. Henry, 1981. The distribution of fructans in onions. J. New Phytol., 81: 29-34.
Direct Link - Deshmukh, S.N., M.S. Basu and P.S. Reddy, 1986. Genetic variability, character association and path coefficients of quantitative traits in Virginia bunch varieties of groundnut. Indian J. Agric. Sci., 56: 816-821.
Direct Link - Dewey, D.R. and K.H. Lu, 1959. A correlation and path-coefficient analysis of components of crested wheatgrass seed production. Agron. J., 51: 515-518.
CrossRefDirect Link - Dowker, B.D., R.C. Hardwick, R.C. Fennell and D.J. Andrews, 1976. Genotypic and environmental correlations between leaf growth and bulb size in onions. Ann. Applied Biol., 82: 341-348.
CrossRef - Falconer, D.S., 1989. Introduction to Quantitative Genetics. 3rd Edn., Longman Scientific and Technical, New York, USA Pages: 438.
Direct Link - Figliuolo, G., V. Candido, G. Logozzo, V. Miccolis and P.L.S. Zeuli, 2001. Genetic evaluation of cultivated garlic germplasm ( Allium sativum L. and A. ampeloprasum L.). Euphytica, 121: 325-334.
CrossRefDirect Link - Galvan, G.A., W.A. Wietsma, S. Putrasemedja, A.H. Permadi and C. Kik, 1997. Screening for resistance to anthracnose (Colletotrichum gloeosporioides Penz.) in Allium cepa and its wild relatives. Euphytica, 95: 173-178.
CrossRef - Gichimu, B.M. and C.O. Omondi, 2010. Morphological characterization of five newly developed lines of Arabica coffee as compared to commercial cultivars in Kenya. Int. J. Plant Breed. Genet., 4: 238-246.
CrossRefDirect Link - Jalata, Z., A. Ayana and H. Zeleke, 2011. Variability, heritability and genetic advance for some yield and yield related traits in Ethiopian barley (Hordeum vulgare L.) landraces and crosses. Int. J. Plant Breed. Genet., 5: 44-52.
CrossRefDirect Link - Johnson, C.E. and T.P. Hernandez, 1980. Heritability studies of early and total yield in tomatoes. Horti. Sci., 15: 280-285.
Direct Link - Johnson, H.W., H.F. Robinson and R.E. Comstock, 1955. Genotypic and phenotypic correlations in soybeans and their implications in selection. Agron. J., 47: 477-483.
CrossRefDirect Link - Lee, W.S., Y.C. Kim and B.C. Lee, 1977. Varietal characters and genetic correlations in different ecotypes of garlic. Korean J. Breed., 9: 149-157.
Direct Link - Mohanty, B.K., 2001. Genetic variability, inter-relationship and path analysis in onion. J. Trop. Agric., 39: 17-20.
Direct Link - Mulungu, L.S., S.O.W.M. S. Reuben, Nchimbimsolla, R.N. Misangu, L.B. Mbilinyi and M.M. Macha, 1998. Performance of nine exotic and local onion (Allium cepa L.) genotypes grown under a dry season tropical condition at Morogoro, Tanzania: 1. Yield and its components. S. Afr. J. Sci., 94: 451-454.
Direct Link - Nikhila, K.R., V.B. Sureshkumar, K.V. Mohanan and A. Santaram, 2008. Association of agronomic characters in robusta coffee (Coffea canephora Pierre ex Froehner). Int. J. Plant Breed. Genet., 2: 47-50.
CrossRefDirect Link - Parthiban, K.T., R. Kirubashankkar, M. Paramathma, V. Subbulakshmi and P. Thiyagarajan et al., 2011. Genetic association studies among growth attributes of jatropha hybrid genetic resources. Int. J. Plant Breed. Genet., 5: 159-167.
CrossRef - Umamaheswari, R. and K.V. Mohanan, 2011. A study of the association of agronomic characters in Vanilla planifolia andrews. Int. J. Plant Breed. Genet., 5: 53-58.
CrossRefDirect Link