
Jean Aghofack-Nguemezi
Department of Plant Biology, Faculty of Sciences, University of Dschang, P.O. Box 67, Dschang, Cameroon
Valere Tatchago
Institute of Agricultural Research for Development, P.O. Box 44, Dschang, Cameroon
International Journal of Agricultural Research
Year: 2010 | Volume: 5 | Issue: 10 | Page No.: 821-831







ABSTRACT
The study was undertaken to determine the effects of calcium and magnesium nutrients on the development of plants and subsequent postharvest conservation of tomato fruits. Tomato plants were treated by applications of N/P/K (9.5/8/10) and fertilizers containing Ca2+ and/or Mg2+. Control plants received only N/P/K. Two fertilizer combinations (N/P/K + foliar spray of Manvert Magnesium and N/P/K + calcium nitrate at 800 kg ha-1 + foliar spray of Manvert Magnesium) induced a significant delay in the flowering of plants. Combinations of soil applications of N/P/K and calcium nitrate at 200 kg ha-1 with foliar sprays of Manvert Calcium and/or Manvert Magnesium led to significant increases in the content of Ca2+ in mature-green fruits and subsequently to the delay of their ripening and the prolongation of the conservation period. Combinations of N/P/K + calcium nitrate at 200 kg ha-1 + Manvert Magnesium and N/P/K + calcium nitrate at 400 kg ha-1 + Manvert Calcium + Manvert Magnesium led to significant increases in the Mg2+ content in fruits. Fruits produced by plants that received these fertilizer combinations also showed a prolongation of the duration of ripening period and that of the conservation. The longest shelf-life was obtained after simultaneous applications on soil of N/P/K and calcium nitrate at 200 kg ha-1 and foliar sprays of Manvert Calcium and Manvert Magnesium. These results indicated that calcium and magnesium could be considered as key elements of fertilizers with regard to the delay of ripening of mature-green tomato fruits and to the prolongation of the shelf-life of the red-ripe ones.
PDF Abstract XML References Citation
Received: March 25, 2010;
Accepted: June 08, 2010;
Published: June 26, 2010
How to cite this article
Jean Aghofack-Nguemezi and Valere Tatchago, 2010. Effects of Fertilizers Containing Calcium and/or Magnesium on the Growth, Development of Plants and the Quality of Tomato Fruits in the Western Highlands of Cameroon. International Journal of Agricultural Research, 5: 821-831.
DOI: 10.3923/ijar.2010.821.831
URL: https://scialert.net/abstract/?doi=ijar.2010.821.831
DOI: 10.3923/ijar.2010.821.831
URL: https://scialert.net/abstract/?doi=ijar.2010.821.831
INTRODUCTION
In the course of the past decades, coffee was the main source of income for most of farmers in the western highlands of Cameroon. But now, these farmers have to face the problems of the drastic price reduction and the decrease in the production of these cash crops due to the exhaustion of cultivated soils (Kuete, 2008; Gillermou and Kamga, 2004). It therefore becomes urgent to search for alternative income generating products. In this context, seasonal cultures such as vegetables and fruits are gaining more and more interest.
Indeed, fruits like tomatoes can be cultivated as well during the raining season on lands as during the dry season on marshes. Thus, farmers can harvest tomato fruits twice a year (Uwizeyimana, 2009).
The tomato fruit is one of most widely grown fruits for consumption with more than 122 million tons being produced worldwide in 2005 (FAOSTAT, 2005). It plays an important role in human nutrition due to its content in flavonoids, carotenoids and vitamins (Abushita et al., 2000; Scalbert and Williamson, 2000; Rodriguez-Amaya, 1999). β-Carotene found in tomatoes is the most potent dietary precursor of vitamin A, the deficiency of which leads to blindness and premature death (Mayne, 1996). Carotenoids and flavonoids, when taken regularly and in considerable quantities, provide health benefit by decreasing the risk of disorders (e.g., certain cancers and cardiovascular diseases) and the incidence of age-related degeneration (Santagelo et al., 2007; Giovannucci, 1999; Weisburger, 1998; Seddon et al., 1994).
Although, new cultivars are already known, most of the developing countries are still facing enormous postharvest losses in quality (appearance, firmness, flavor and nutritional value) and quantity of tomato fruits mainly due to the inaccessibility of genetically ameliorated seeds and inadequate fruit conservation methods (Kader, 1986, 2005). Generally, the productivity of plants and quality of fruits can be ameliorated either through genetic engineering or cultivation practices. In this regard, applications of fertilizers containing calcium and/or magnesium concomitantly with standard N/P/K-fertilizers could positively influence the growth and development of plants as well as the ripening and ageing of fruits produced. Calcium participate in the regulation of many processes related to plant growth and development including biomass partitioning and fruit yield (Hao and Papadopoulos, 2004), signal transduction (Hepler, 2005; Navarro-Avino and Bennett, 2005; Trewavas, 2000; Malho, 2000; Raz and Flur, 1992), induction of early auxin-responsive genes (Singha et al., 2006), resistance to pathogens (Ho and White, 2005; Ehret et al., 2002), stress-tolerance (Jiang et al., 2005; Jiang and Huang, 2001), chlorophyll biosynthesis (Jiang and Huang, 2001), pollination (Dumas and Gaude, 2006), senescence (Hepler and Wayne, 1985; Leshem, 1991) and fruit ripening (Aghofack-Nguemezi and Dassie, 2007; Park et al., 2005; Aghofack-Nguemezi and Yambou, 2005). While intracellular Ca2+ concentration is submicromolar, the concentration of closely related divalent cation Mg2+ is millimolar. Despite the concentration difference that would favor Mg2+, cellular processes display enormous selectivity for Ca2+ (Hepler, 2005). It has recently been reported that although Mn2+ and Mg2+ enhanced the nitrate uptake and growth of barley seedlings under saline conditions, none of them was as effective as Ca2+ (Ward et al., 2005).
Notwithstanding the positive physiological roles of Ca2+ and Mg2+ in plant growth and development the use of conventional fertilizers such as N/P/K mixtures or urea is still, to the best of our knowledge, a common agricultural practice in the western highlands of Cameroon. The present study was thus undertaken to examine the influence of different fertilizers containing Ca2+ and Mg2+ applied either by spraying on aerial plant parts or by application on the soil on the growth, crop yield and some parameters related to the quality, ripening and senescence of tomatoes.
MATERIALS AND METHODS
Description of the Experimental Site
The experiment was conducted at the research farm of the Institute of Agricultural Research for Development in Dschang from September 2006 to March 2007. Dschang is
| Table 1: | Means of the daily duration of sunshine, of monthly pluviometry and daily temperature during the experimental period as recorded by the meteorological station of the Institute of Agricultural Research for the development in Dschang |
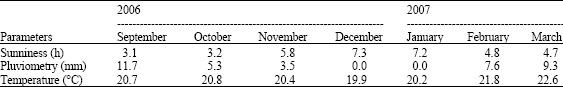 | |
| Table 2: | Compositions of different fertilizer combinations |
 | |
located in the western highlands of Cameroon at 5.26°N latitude, 10.26°E longitude and 1400 m altitude. The experimental period coincided with least rainfall and highest sunniness in the locality (Table 1).
Experimental Design and Plant Materials
Seeds of Solanum lycopersicum Mill. var. Rio Grande were sowed in a nursery on a soil enriched with urea and hen droppings at the doses of 300 and 26.1 g m-2, respectively. Seedlings in the nursery were treated with the fungicides Banko PlusTM (chlorothalomil + carbendazime) and BeauchampTM (metalaxyl + mancozebe). Forty eight days after sowing, seedlings were pricked out in a soil previously enriched with 3 t ha-1 hen droppings according to a randomized complete bloc design with seventeen treatments (Table 2) and four replications. Urea, super phosphate and potassium chloride were applied on the soil respectively 11, 14 and 15 days after planting out of seedlings at doses of 95, 80 and 100 kg ha-1, respectively. Control tomato plants received only N/P/K (9.5/8/10) whereas other plants received N/P/K, calcium nitrate and/or liquid fertilizers containing Ca2+ or Mg2+. Liquid fertilizers were of the trademark Manvert (BIOVERT, Spain). The ManvertTM solutions (3 mL L-1) were sprayed weekly on the aerial parts of tomato plants from 24 days after pricking out onwards till the harvest of the first mature fruits. Tomato plants were in case of need watered; they were treated with the insecticide CalllidimTM (dimethoate) and the fungicides BankoTM and BeauchampTM. A manual weeding was done when necessary.
Determination of the Number Days after Pricking out Required for the Flowering of 50% of Plants and of the Percentage of Abortion of Floral Buds and Flower
The time in days passed between the pricking out of seedlings and the flowering of 50% of plants in each plot was recorded.
Floral buds were counted from 38 days after pricking out onwards. Seven days before the harvest of the first mature fruits, the total numbers of floral buds, flowers and fruits in each plot were recorded. The percentage of floral bud and flower abortion was calculated according to Tarchoun and Dridi (Tarchoun and Didri, 2005).
Determination of the Diameter and Height
The diameter and height of plants were measured at the sixth week after pricking out. The height was measured from the first branching upwards. The diameter of the stem was measured at the level of collar. At harvest, the diameter of fruits was also measured both laterally and longitudinally. The measurement of the diameter was done using a vernier caliper.
Determination of Water Content in Fruits and Estimation of Crop Yield
Fresh tomato fruits were weighed and dried in an oven successively at 65 and 105°C for 5 day and 24 h, respectively. They were then weighed for the determination of dry matter weight according to Chapman (1976). The crop yield was obtained by calculating the total fresh weight of tomato fruits harvested in all the plots that received the same treatment.
Determination of Carotenoid Content
The 0.5 g of the peel of mature green tomato fruits was ground and extracted with a mixture of benzene/acetone/water (15/75/10, v/v). The extract was centrifugated at 400 rpm for 30 min. The absorbance of the extract was then measured at 477 nm. The carotenoid content in peel extract of tomato fruits was calculated according to Armenta et al. (2006).
Determination of Calcium and Magnesium Contents
Fresh tomato fruits were dried in an oven at 105°C for 24 h. One gram of the dried samples was then calcined at 405°C for 24 h. Ten milliliter of 1 N nitric acid solution were added to the ash and the mixture was heated till the evaporation of half of the volume. The residue was completed to 50 mL with distilled water. This solution was further threefold diluted before use. Ca2+ and Mg2+ contents were determined by the complexometric method as described by Pauwels et al. (1992).
Determination of the Duration of the Ripening Period and of the Shelf-Life
Tomato fruits at mature green stage were harvested and stored at 25°C. The number of days elapsed between the mature green stage and the red-ripe stage for 100% of tomato fruits in each lot was recorded. The shelf-life was considered as the length of time between the red-ripe stage and the trickling of 100% of fruits.
Statistical Analysis
This analysis was done with the GraphPad InStat software. Group comparisons were made using One-way Analysis of Variance (ANOVA) to see if variations among the means were significantly greater than expected by chance. The Student-Newman-Keuls Test was used to compare means differences, whereby a p value of <0.05 was considered as statistically significant. The Linear Regression Test was used to determine the correlation coefficients between some relevant parameters.
RESULTS AND DISCUSSION
Growth and Development
No significant effect of fertilizers containing calcium and/or magnesium on stem diameter and height of tomato plants, on the percentage of abortion of floral buds and the percentage of abortion of flowers could be observed (Table 3). Treatments of tomato plants with T6 (N/P/K application on the soil and spray of Manvert Magnesium solution on aerial plant parts (SMMS)) or T15 (simultaneous application of N/P/K and 800 kg ha-1 calcium nitrate on the soil and SMMS) induced significant increases in the number of days after planting out required by 50% of tomato plants to flower (Table 3). Obviously, calcium and magnesium ions could regulate the flowering duration in tomato plants. This finding corroborated with previous results obtained from Pharbitis nil. It has thus been reported that Ca2+ was involved in the photoperiodic flower induction process of this species and that its endogenous level may be a limiting factor (Friedman et al., 1989). Furthermore, based on results of the analysis of ion distribution in shoot apex of Pharbitis nil, Kobayashi et al. (2006) suggested for the first time that Mg2+ plays an important role in flower induction.
Fruit Quality and Yield
There was no significant alteration in water and total carotenoid contents in tomato fruits produced by plants that received fertilizers containing Ca2+ and/or Mg2+ in comparison to fruits from control plants (Table 4). The calcium content in tomato fruits from plants that received fertilizers containing Ca2+ and/or Mg2+ were generally higher than that found in fruits produced by control plants. However, this increase trend was not overall statistically confirmed. Thus, calcium contents in tomato fruits produced by plants that received T12 (N/P/K + calcium nitrate at 400 kg ha-1 + SMMS), T13 (N/P/K + calcium nitrate at 400 kg ha-1 + spray of Manvert Magnesium solution on aerial plant parts (SMCS) + SMMS) and T15 did not change significantly when compared to fruits produced by control plants. No synergistic effect of soil application of Ca(NO3)2 and foliar spray of Manvert Calcium on Ca2+ content in fruits could be observed. A slight negative correlation (r = -0.21) between the dose of Ca(NO3)2 applied on the soil and the Ca2+ content in fruits was found (Table 4). These results demonstrated that applications of fertilizers containing Ca2+ either on the soil at doses = 400 kg ha-1 or on aerial plant parts led to increases in the Ca2+ concentration in fruits.
| Table 3: | Effects of fertilizers containing calcium and/or magnesium on stem diameter and height of tomato plants, on the number of days after planting out required for the flowering of 50% of plants (D50F), the percentage of abortion of floral buds (PAFB) and the percentage of abortion of flowers (PAF) |
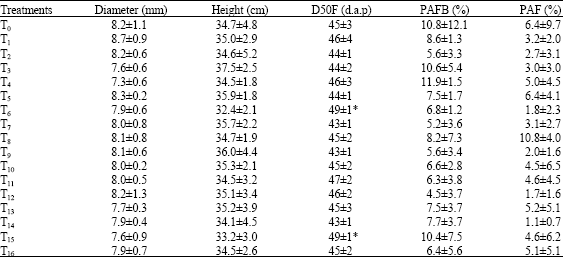 | |
| Values are Mean±SD (n = 4). d.a.p.:days after pricking out, *Values are statistically different from T0 at p<0.05 | |
| Table 4: | Effects of fertilizers containing calcium and/or magnesium on contents of water, carotenoids, calcium and magnesium ions in tomato fruits, and the correlation between the dose of Ca(NO3)2 applied on the soil and the Ca2+ content in fruits |
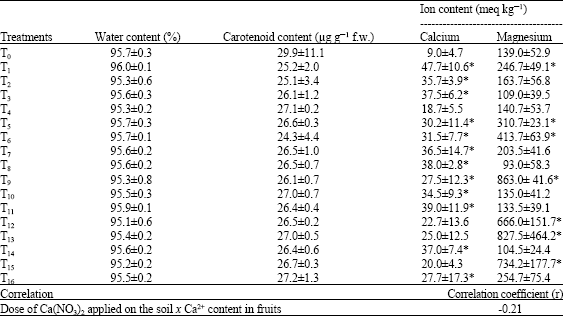 | |
| Values are Mean±SD (n = 4). f.w.: fresh weight, *Values are statistically different from T0 at p<0.05 | |
In a similar study, foliar applications of calcium salts were also shown to increase Ca2+ levels in tomato fruits (Garcia et al., 1995). It is well known that mineral ions absorbed by roots are transported in long distances in the xylem concurrently with water (Opik et al., 2005). Most plants can also absorb mineral nutrient applied to their leaves as sprays and this mode of uptake is most effective when the nutrient solution remains on the leaf as a thin film (Taiz and Zeiger, 2006). As indicated by the negative correlation between the dose of Ca(NO3)2 used and the Ca2+ content in fruits, the former was in inverse ratio to the latter. Thus, tomato plants could no longer absorb calcium ions or transport them in growing fruits when high doses of Ca(NO3)2 were applied on the soil. The mechanisms whereby Ca2+ or NO3- could at high concentration inhibit the absorption of the former ion by roots of tomato plants are not known. Nevertheless, Ca2+ has been shown to be very effective in the enhancement of the uptake of nitrate by barley seedlings under saline conditions. At 800 kg ha-1 of Ca(NO3)2 Ca2+ might thus, to the detriment of its own absorption, enhance the uptake of NO3- by tomato plants. The mechanisms whereby liquid fertilizers containing Mg2+ triggered an increase in Ca2+ content in tomato fruits are not yet known. An increase in the level of Ca2+ in banana fruits after treatment by dipping in MgSO4 solution has also previously been observed (Aghofack-Nguemezi and Dassie, 2007).
In most of cases where tomato plants received fertilizers containing Mg2+, magnesium ion contents in fruits produced by these plants were significantly higher than those observed in the extracts of fruits from control plants (Table 5). In this context, when compared to the control, higher contents in Mg2+ ions were found in fruits produced by tomato plants that received T9 (N/P/K + calcium nitrate at 200 kg ha-1 + SMMS), T12 (N/P/K + calcium nitrate at 400 kg ha-1 + SMMS), T13 (N/P/K + calcium nitrate at 400 kg ha-1 + SMCS + SMMS) or T15 (N/P/K + calcium nitrate at 800 kg ha-1 + SMMS). However, the content of Mg2+ also significantly increased in tomato fruits produced by plants that received only fertilizers containing Ca2+, namely T1 (N/P/K + calcium nitrate at 200 kg ha-1) or T5 (N/P/K + SMCS).
| Table 5: | Effects of fertilizers containing calcium and/or magnesium on the yield, diameter, duration of the ripening period (DRP) and that of shelf-life (DSL) of tomato fruits, and correlations between ion contents and DRP or DSL |
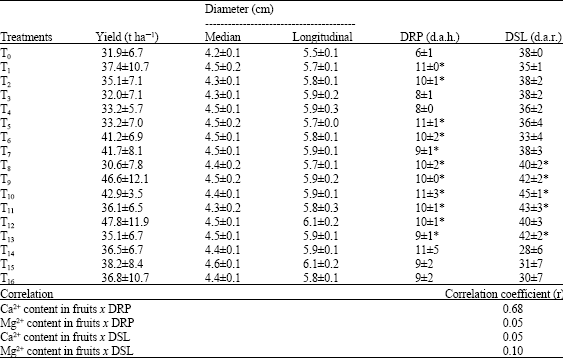 | |
| Values are Mean±SD (n = 4), d.a.h.: Days after harvesting, d.a.r.: Days after ripeness, *Values are statistically different from T0 at p<0.05 | |
These results indicated that as it generally the case for most of mineral nutrients (Taiz and Zeiger, 2006; Opik et al., 2005), Mg2+ could be absorbed through the leaves of plants and then transported into developing tomato fruits. The mechanisms whereby fertilizers containing Ca2+ triggered an increase in Mg2+ content in tomato fruits are not yet known.
No significant difference could be observed between the total yield and the diameter of tomato fruits produced by plants which received fertilizers containing Ca2+ and/or Mg2+ and that of fruits produced by control plants (Table 5). In contradiction to these results, Hao and Papadopoulos (2003) reported that Ca2+ at 300 mg L-1 and Mg2+ at 20 mg L-1 nutrient solutions led, respectively to increase and decrease in tomato fruit yield. In a ferrallitic sandy loam soil, tomato fruit yield was significantly increased by liming with CaO at a pH of 5.2 while at a pH of 5.7 and above liming had little effects; under those conditions applied Mg2+ had no effect on fruit yield (Asiegbu and Uzo, 1983). Suplementary calcium enhanced fruit yield in strawberry cultivars grown at high NaCl salinity (Kaya et al., 2002).
Ripening and Senescence of Fruits
The number of days elapsed between the mature green stage and the red-ripe stage of fruit ripening was greatly affected by treatments of tomato plants in the farm with fertilizers containing Ca2+ and/or Mg2+ (Table 5). Nearly all these fertilizers combinations induced a prolongation of the duration of the ripening period of tomato fruits. Apart from tomato fruits produced by plants that received T3 (N/P/K + calcium nitrate at 800 kg ha-1), T14 (N/P/K + calcium nitrate at 800 kg ha-1 + SMCS), T15 and T16 (N/P/K + calcium nitrate at 800 kg ha-1 + SMCS + SMMS), fruits produced by plant that received all other fertilizers containing Ca2+ and/or Mg2+ had a longer duration of ripening period than those from control plants. The length of time between the red-ripe stage and the trickling of 100% of tomato fruits (the shelf-life) was mostly significantly prolonged by the applications of T8 (N/P/K + calcium nitrate at 200 kg ha-1 + SMCS), T9 (N/P/K + calcium nitrate at 200 kg ha-1 + SMMS), T10 (N/P/K + calcium nitrate at 200 kg ha-1 + SMCS + SMMS), T11 (N/P/K + calcium nitrate at 400 kg ha-1 + SMCS), T12 (N/P/K + calcium nitrate at 400 kg ha-1 + SMMS) and T13.
The positive correlation between Ca2+ content and the duration of ripening period of tomato fruits was more pronounced (r = 0.68) than that found between the magnesium content and this period (r = 0.05). The positive correlation between the Mg2+ content and the duration of the shelf-life was more marked (r = 0.1) than the correlation found between Ca2+ content and the duration of the shelf-life (r = 0.05) (Table 5).
Obviously, calcium nitrate at doses ≤400 kg ha-1 or foliar spray of Manvert Calcium induced increases in Ca2+ content and subsequently a delay of the ripening process of tomato fruits. When soil application of Ca(NO3)2 was associated to foliar spray of Manvert Calcium and/or Manvert Magnesium, both the ripening of mature green tomato fruits and the trickling of red ripe fruits were subsequently retarded. This means that even if no synergistic effect of soil application of Ca(NO3)2 and foliar spray of Manvert Calcium on Ca2+ content in fruits was observed, the combination of both types of fertilization could lead to an increase in the duration of ripening period as well as a delay of the onset of senescence of red-ripe tomato fruits. It has previously been demonstrated that fruit from tomato plants expressing Arabidopsis Thaliana H+/cation exchangers have more Ca2+ and prolonged shelf life when compared to controls (Park et al., 2005). Postharvest calcium applications on loquat (Akhtar et al., 2010) and peach fruits (Manganarisa et al., 2007) retained their firmness, thereby delaying the ripening process. It is well established that calcium ion can delay the ripening and senescence by stabilizing cell membrane and increasing the rigidification of monolayers. Moreover, the Ca2+-mediated cross-linking may occur as bridging between phospholipids, between phospholipids and corboxyltails of embedded membrane proteins and between phospholipids and cytoskeletal. Magnesium ion affects the electrostatic cross-linking between membranes components to a lesser extend than calcium ion (Leshem, 1991). There are also several calcium-pectate interactions which make the cell wall firmer (Carpita and McCann, 2000). It is not yet known why combination of application on the soil and foliar spray of fertilizers containing Ca2+ (e.g., T8 and T11) induced a retardation of both the ripening and the senescence of tomato fruits while soil application of Ca(NO3)2 (e.g., T1 and T2) or foliar spray of Manvert Calcium (e.g., T5) alone led only to the increase in duration of the ripening period. The fact that combinations of soil application of Ca(NO3)2 with foliar spray of Manvert Magnesium (e.g., T9) also led to increases in both the duration of ripening period and that of shelf-life could be explained by the additional effects of Mg2+. Indeed, due to their divalent character, Mg2+ and Mn2+ can with lesser effectiveness play physiological roles ascribed to Ca2+ (Ward et al., 2005; Leshem, 1991) or quite different roles (Fanasca et al., 2006). The appropriate Mg2+ concentration in the nutrient solution required for the production of winter greenhouse tomato crop was lower than that of Ca2+ (Hao and Papadopoulos, 2004). High proportion of Ca2+ as related to K/Ca/Mg ratio in the nutrient solution was reported to improve fruit yield and reduce the incidence of blossom-end rot whereas high proportion of Mg2+ rather resulted in the increase of total antioxidant activity in tomato fruits (Fanasca et al., 2006). The retarding effect of foliar spray of Manvert Magnesium alone on the ripening process could be related to increases in Mg2+ and Ca2+ contents in tomato fruits.
CONCLUSION
The most striking features of field application of fertilizers containing Ca2+ and/or Mg2+ were increases in the duration of flowering of plants and in calcium contents in the extracts and the subsequent prolongation of the duration of mature green stage of tomato fruits. Combinations of soil application of Ca(NO3)2 at doses ≤400 kg ha-1 with foliar spray of Manvert Calcium and/or Manvert Magnesium solutions were the most efficient in inducing the delay of ripening and senescence of tomato fruits. Calcium ion seemed to be a key player in the modulation of the ripening process while Mg2+ could rather play an important role in the regulation of the senescence of tomato fruits.
REFERENCES
- Abushita, A.A., H.G. Daood and P.A. Biacs, 2000. Change in carotenoids and antioxidant vitamins in tomato as a function of varietal and technological factors. J. Agric. Food Chem., 48: 2075-2081.
CrossRefDirect Link - Aghofack-Nguemezi, J. and B. Dassie, 2007. Effects of salts and edible oils on calcium and water contents in ripening banana fruits. J. Plant Sci., 2: 302-309.
CrossRefDirect Link - Aghofack-Nguemezi, J. and T.N. Yambou, 2005. Effects of calcium chloride and magnesium sulphate treatments on the shelf-life of climacteric banana and non-climacteric pineapple fruits. Cam. J. Exp. Biol., 1: 34-38.
Direct Link - Akhtar, A., N.A. Abbasi and A. Hussain, 2010. Effect of calcium chloride treatments on quality characteristics of loquat fruit during storage. Pak. J. Bot., 42: 181-188.
Direct Link - Armenta, R.E., A. Burja, H. Radianingtyas and C.J. Barrow, 2006. Critical assessement of various techniques for the extraction of carotenoids and co-enzyme Q10 from the thraustochytrid strain ONC-T18. J. Agric. Food Chem., 54: 9752-9758.
PubMedDirect Link - Asiegbu, J.E. and J.O. Uzo, 1983. Effect of lime and magnesium on tomato (Lycopericon esculentum Mill.) grown in a ferrallitic sandy loam tropical soil. Plant Soil, 74: 53-60.
CrossRef - Dumas, C. and T. Gaude, 2006. Fertilisation in plant: Is calcium a key player? Semin. Cell Dev. Biol., 17: 244-253.
PubMedDirect Link - Ehret, D.L., J.G. Menzies, C. Bogdanoff, R.S. Utkhede and B. Frey, 2002. Foliar applications of fertilizer salt inhibit powdery mildew on tomato. Can. J. Plant Pathol., 24: 437-444.
CrossRefDirect Link - Fanasca, S., G. Colla, G. Maiani, E. Venneria, Y. Rouphael, E. Azzini and E. Saccardo, 2006. Changes in antioxidant content of tomato fruits in response to cultivar and nutrient solution composition. J. Agric. Food Chem., 54: 4319-4325.
CrossRefDirect Link - Friedman, H., E.E. Goldsmidt and A.H. Halevy, 1989. Involvement of calcium in the photoperiodic flower induction process of Pharbitis nil. Plant Physiol., 89: 530-534.
Direct Link - Garcia, J.M., J.M. Ballesteros and M.A. Albi, 1995. Effects of foliar applications of CaCl2 on tomato stored at different temperatures. J. Agric. Food Chem., 43: 9-12.
CrossRefDirect Link - Giovannucci, E., 1999. Tomatoes, tomato-based products, lycopene and cancer: Review of the epidemiologic literature. J. Nat. Cancer Inst., 91: 317-331.
PubMed - Hao, X. and A.P. Papadopoulos, 2004. Effects of calcium and magnesium on plant growth, biomass partitioning and fruit yield of winter greenhouse tomato. Hort. Sci., 39: 512-515.
CrossRefDirect Link - Hao, X. and A.P. Papadopoulos, 2003. Effects of calcium and magnesium on growth, fruit yield and quality in fall greenhouse tomato crop grown on rockwool. Can. J. Plant Sci., 83: 903-912.
CrossRefDirect Link - Hepler, P.K., 2005. Calcium: A central regulator of plant growth and development. Plant Cell, 17: 2142-2155.
Direct Link - Hepler, P.K. and R.O. Wayne, 1985. Calcium and plant development. Ann. Rev. Plant Physiol., 36: 397-439.
CrossRef - Ho, L.C. and P.J. White, 2005. A cellular hypothesis for the induction of blosom-end rot in the tomato fruits. Ann. Bot., 95: 571-581.
CrossRefDirect Link - Jiang, T., X. Zhan, Y. Hu, L. Zhou and L. Zong, 2005. Roles of calcium in stress-tolerance of plants and its ecological significance. Ying Yong Sheng Tai Xue Bao., 16: 971-976.
PubMedDirect Link - Jiang, Y. and B. Huang, 2001. Effects of calcium and antioxidant activities and water relations associated with heat tolerance in two cool-season grasses. J. Exp. Bot., 52: 341-349.
Direct Link - Kader, A.A., 2004. Increasing food availability by reducing postharvest losses of fresh produce. ISHS Acta Hort., 682: 2169-2176.
Direct Link - Kader, A.A., 1986. Effects of postharvest handling procedures on tomato quality. Acta Hort., 190: 209-221.
Direct Link - Kaya, C., H. Kirnak, D. Higgs and K. Saltali, 2002. Supplementary calcium enhances plant growth and fruit yield in strawberry cultivars grown at high (NaCl) salinity. Sci. Hortic., 93: 65-74.
CrossRef - Kobayashi, N.I., K. Tanoi and T.M. Nakanishi, 2006. Magnesium localization in shoot apices during flower induction in Pharbitis nil. Can. J. Bot., 84: 1908-1916.
Direct Link - Mayne, S.T., 1996. Beta-carotene, carotenoids and disease prevention in humans. FASEB. J., 10: 690-701.
Direct Link - Manganarisa, G.A., M. Vasilakakisa, G. Diamantisida and I. Mignanib, 2007. The effect of postharvest calcium application on tissue calcium concentration, quality attributes, incidence of flesh browning and cell wall physicochemical aspects of peach fruits. Food Chem., 100: 1385-1392.
CrossRef - Park, S., N.H. Cheng, J.K. Pittman, K.S. Yoo, J. Park, R.H. Schmidt and K.D. Hirschi, 2005. . Increase calcium levels and prolonged shelf life in tomatoes expressing Arabidopsis H+/Ca2+ transporters. Plant Physiol., 139: 1194-1196.
CrossRefDirect Link - Rodriguez-Amaya, D.B., 1999. Latin American food sources of carotenoids. Arch. Latinoam. Nutr., 49: 74S-84S.
PubMedDirect Link - Santagelo, C., R. Vari, B. Scazzochio, R. di Benedetto, C. Filesi and R. Masella, 2007. Polyphenols, intracellular signaling and inflammation. Ann. Ist. Super Sanita, 43: 394-405.
PubMedDirect Link - Scalbert, A. and G. Williamson, 2000. Dietary intake and bioavailability of polyphenols. J. Nutr., 130: 2073S-2085S.
PubMedDirect Link - Seddon, J.M., U.A. Ajami, R.D. Sperato, R. Hiller and N. Blair et al., 1994. Dietary carotenoids, vitamin A, vitamin C and vitamin E and advance age-related macular degeneration. J. Am. Med. Ass., 272: 1413-1420.
PubMedDirect Link - Singha, B., A. Chugh, J.P. Khurana and P. Khurana, 2006. An early auxin-responsive Aux/IAA gene from wheat (Triticum aestivum) is induced by epibrasssinolide and differentially regulated by light and calcium. J. Exp. Bot., 57: 4059-4070.
CrossRefDirect Link - Weisburger, J.H., 1998. Evaluation of the evidence on the role of tomato products in disease prevention. Proc. Soc. Exp. Biol. Med., 218: 140-143.
Direct Link