
P. Senguttuvel
Centre for Plant Breeding and Genetics, TNAU, Coimbatore, 641 003, India
M. Raveendran
Centre for Plant Molecular Biology, TNAU, Coimbatore, 641 003, India
C. Vijayalakshmi
Department of Plant Physiology, TNAU, Coimbatore, 641 003, India
K. Thiyagarajan
Centre for Plant Breeding and Genetics, TNAU, Coimbatore, 641 003, India
J.R. Kannan Bapu
Centre for Plant Breeding and Genetics, TNAU, Coimbatore, 641 003, India
B. C. Viraktamath
Directorate of Rice Research, Rajendranagar, Hyderabad, 500 030, India
International Journal of Agricultural Research
Year: 2010 | Volume: 5 | Issue: 9 | Page No.: 708-719







ABSTRACT
In the present study, genotyping through microsatellite markers for salinity tolerance in a set of twenty five genetically divergent genotypes and phenotyping for Na+/K+ ratio grown under Yoshida solution with 60 and 120 mM NaCl and the mechanism behind the association was detected. The polymorphic SSR markers already reported for major saltol QTLs was utilized in these studies revealed highly reproducible. The cluster and principal component analysis allowed a clear grouping of 25 genotypes grouped into 8 distinct clusters of resistant and susceptible genotypes with high and low level of Na+/K+ ratio. There was a highly significant correlation (R2 = 88.75; p = 5%) between the SSR markers and physiological trait based on ion analysis. The association of SSR markers viz., RM 23, 493 and 8053 for the trait linked to Na+/K+ ratio can be regarded as the most reliable markers for marker-assisted selection to identify salinity tolerance in rice. The study revealed that the selection of genetically diverse and resistant genotypes based on association of Na+/K+ ratio with molecular markers is reliable. These markers can also be used to screen large set of germplasm collection to identify and discriminate more salt tolerant rice genotypes from susceptible based on sequence homology with already identified salt tolerant rice genotypes, which can be utilized as donors in the breeding programme for generating salt tolerant varieties.
PDF Abstract XML References Citation
Received: March 20, 2010;
Accepted: June 04, 2010;
Published: June 26, 2010
How to cite this article
P. Senguttuvel, M. Raveendran, C. Vijayalakshmi, K. Thiyagarajan, J.R. Kannan Bapu and B. C. Viraktamath, 2010. Molecular Mechanism of Salt Tolerance for Genetic Diversity Analysed in Association with Na+/K+ Ratio through SSR Markers in Rice (Oryza sativa L.). International Journal of Agricultural Research, 5: 708-719.
DOI: 10.3923/ijar.2010.708.719
URL: https://scialert.net/abstract/?doi=ijar.2010.708.719
DOI: 10.3923/ijar.2010.708.719
URL: https://scialert.net/abstract/?doi=ijar.2010.708.719
INTRODUCTION
Rice is an important stable food crop and two-third of world population depends on it, however due to adverse biotic, abiotic and soil factors, the productivity is declining and unable to meet out for growing population. Among abiotic stress, salinity is foremost and second most widespread problem causing reduction in growth and productivity (Gregario et al., 1997; Ashraf and O’Leary, 1996; Munns et al., 2006). So, the possible ways are reclamation of soil and breeding new varieties suitable for saline soils, however reclamation needs more financial needs, laborious man power and not always practically feasible. The other possible strategy is breeding to enhance salinity tolerance, but it has been slow due to limited knowledge about the genetics of salt tolerance, inadequate screening techniques, low selection efficiency and poor GxE interactions.
Generally, salinity tolerance is a polygenic trait. Screening rice germplasms to locate salt-tolerant genes for use in improving the currently grown varieties is of continuous importance to plant biotechnologists (Flowers, 2004). One major approach in plant breeding is to maximize the genetic diversity between parental genotypes for hybridization. There are rice varieties which yield high under normal condition; they fail to perform in salt affected soils. So, genetic diversity screening for trait of interest is an essential part of the commencement of a breeding program. Babu et al. (2006) reported that genetic improvement mainly depends on the amount of genetic variability present in the population. Hence, estimation of genetic diversity for salt tolerance parameters among genotypes is important for planning the future crossing programme.
Integration of breeding with recent marker assisted selection technology; it is now feasible to analyze simply inherited and quantitative traits at early seedling stage, which fasten the breeding program. Molecular markers can now be used to tag quantitative trait loci and to evaluate their contribution to the phenotype by selecting favorable alleles at these loci using Marker-Assisted Selection (MAS) to accelerate genetic improvement. DNA based molecular markers have been used extensively to assess the genetic diversity in crops. Among the marker technology, microsatellite markers have been effectively used to identify genetic variation among rice cultivars (Garland et al., 1999). They can be easily amplified by PCR reaction using DNA nucleotide primers, the unique sequences flanking the repeated motifs. QTL analysis of salt tolerance in rice based on segregating generations has been conducted by several researchers (Flowers et al., 2000; Koyama et al., 2001; Bonilla et al., 2002; Lin et al., 2004; Takehisa et al., 2004; Ren et al., 2005; Leeet al., 2007). However, there are limitations in using this method, Non-availability of mapping populations and substantial time needed to develop such populations are the major limitations in the identification of molecular markers for specific traits. Very large segregating populations are needed to achieve a high resolution map. In linkage-based analyses, only few genotypes are used as parents of mapping populations. Hence, the molecular markers from linkage-based analyses may be specific (polymorphic) to only one particular pedigree. Linkage-based analyses in the past has allowed the identification of genes/QTLs at distances as large as 10-30 cM, which are hardly suitable either for marker-assisted breeding or for identification/cloning of functional genes. Also, the map has a relatively low resolution resulting in the absence of tight linkage between the marker and the trait. A high resolution is necessary for Marker-Assisted Selection (MAS) and other purposes such as cloning a gene close to the marker.
Molecular studies have revealed that salt tolerance is controlled by interactions between several independently regulated but temporally and spatially coordinated processes (Kawasaki et al., 2001; Ozturk et al., 2002; Sekiet al., 2002). Zhang et al. (1995) mapped a major gene for salt tolerance on chromosome 7. Prasad et al. (2000a, b) identified seven QTLs for seedling traits associated with salt stress and mapped these to five different chromosomes. Bonilla et al. (2002) reported QTLs in chromosome 1 using RFLP and SSR markers and identified QTLs for Na+, K+ and Na+/ K+ uptake of phenotypic variation. Koyama et al. (2001) reported the chromosomal location of ion transport and selectivity traits that are compatible with agronomic needs and showed that QTL for Na+ and K+ transport are likely to act through the control of root development and structure and the regulation of membrane localized transport compartmentalization. Lin et al. (2004) have identified a major QTL for high shoot K+ content under salinity stress in the same region of chromosome 1. This study focused on a subset of genotypes to analyze the association of genetic diversity with Na+/K+ ratio utilizing microsatellite markers.
MATERIALS AND METHODS
Plant Materials
Twenty five rice germplasm with diversified genetic background were utilized.
Measurement of Ion Analysis
The seedlings were developed as by growing on sand culture and transplanted on two leaf stage by single seedling per hole on the thermocole and floated on the yoshida nutrient solution. The salinity treatments were imposed 21 onwards using two salinity levels of 60 and 120 mM NaCl.
Sodium and potassium content was assessed in triple acid extract (9:2:1 parts of HNO3:H2SO4:HCLO4) extract using Flame Photometer (Model 21 Systronics) and expressed as mg g-1 on weight basis (Jackson, 1964).
DNA Extraction
Leaf samples were collected from 30 days old plants of rice genotypes, grown at Centre for Plant Breeding and Genetics (CPBG) TNAU, Coimbatore and the duration of study was from 2006 to 2008. Around 5-8 young tender leaves from each genotype were collected in an ice box and the collected leaf samples were used to extract DNA following the CTAB method (Dellaporta et al., 1983). Prior to extraction, the pestle and mortar, spatula and scissors were sterilized by autoclaving. Two grams of leaves were cut into bits with the help of sterile scissors and transferred to mortar. The leaf tissues were frozen in liquid nitrogen and ground into fine powder. The fine powder was allowed to thaw in the presence of 15 mL of pre-heated CTAB extraction buffer containing β mercaptoethanol, in polypropylene centrifuge tubes and incubated for 45 min. at 65° C with occasional mixing. The tubes were removed from the water bath and allowed to cool at room temperature. Equal volume of chloroform and isoamyl alcohol mixture (24:1) was added and mixed by inversion for 15 min. It was centrifuged at 4000 rpm for 20 min in room temperature. The clear aqueous phase was transferred to a new sterile tube. One third volume of ice-cold isopropanol was added and mixed gently by inversion until DNA was precipitated out and incubate it at 20°C for 6 h. Then, it was centrifuged at 4000 rpm for 20 min in room temperature to pellet the DNA and the supernatant was discarded. The DNA pellet was washed with 70% alcohol. The DNA was air dried after washing with 70% alcohol. Depending upon the pellet size the DNA was dissolved in 200-500 μL of TE buffer (pH 8.0). To eliminate the contaminated RNA from DNA one-tenth volume of RNase (10 mg mL-1) was added to DNA sample and incubated at 37°C for 30 min. Equal volume of chloroform: isoamyl alcohol mixture (500 μL) was added and mixed thoroughly by repeated inversions. The mixture was centrifuged at 12000 rpm for 10 min at 4°C and the aqueous phase was transferred to another micro centrifuge tube without disturbing the inner phase. Two volume of absolute alcohol and 1/10 volume of 3 M sodium acetate were added and incubated at -20°C overnight. Then, it was centrifuged to pellet the DNA and the supernatant was discarded. The pellet was washed twice with 70% ethanol. The alcohol was discarded and DNA was air dried completely. Depending upon the size of the pellet, DNA was dissolved in 250-500 μL of TE (pH 8.0) and stored at 4°C.
Quantification of DNA
DNA was checked for its purity and intactness and then quantified. The crude genomic DNA was run on 0.8 per cent agarose gel stained with ethidium bromide following a standard method (Sambrook et al., 1989) and was visualized in a gel documentation system (Alpha ImagerTM1200, Alpha Innotech Corp., CA, USA). DNA was quantifying by using Spectrophotometer. One microliter of crude DNA was diluted to 1 mL deionized water. The absorbance for all accession was measured at 260 nm. DNA has maximal absorbance at 260 nm. An optical density (OD) of 1.0 corresponds to 40 μg mL-1 for double stranded DNA. Based on the quantification data, DNA dilutions were made in TE buffer to a final concentration of 50 ng μL-1 and stored in -20°C for further use.
SSR-PCR Analysis
DNA from 25 genotypes was amplified using a set of 5 SSR primer pairs.
The SSR primers which are located near to the salt QTLs in the previous linkage maps were selected in the present study. Those SSR primers which are amplified in the present set of germplasm were used for diversity analysis after optimizing the PCR conditions. Finally we selected primers that amplified properly. Amplification reactions were done in 15 μL volume containing 120-150 ng of genomic DNA, 0.45 μM forward and reverse of primers, 0.2 mM of each dNTPs, 50 mM KCl, 10 mM Tris HCl, (pH8.3), 2.5 mM MgCl2, 0.01% gelatin and 0.35 units of Taq polymerase. Amplification was performed in PTC Thermal Cycler 100 (MJ Research Inc., Sanfrancisco, USA) programmed for an initial denaturation of 94°C for 12 min, followed by 36 cycles of 30 sec denaturation at 94°C, 30 sec annealing at 52°C and extension of 45 sec at 68°C. Final extension was done to a period of 5 min at 68°C and then product was stored at 4°C.
SSR Analysis
Statistical Analysis of SSR Data
All the genotypes were scored for the presence or absence of the SSR bands and the data were entered into a binary matrix as discrete variables, 1 for presence and 0 for absence of the character and this data matrix was subjected to further analysis.
The genetic associations between varieties were evaluated by calculating the Jaccard’s similarity coefficient for pair wise comparisons based on the proportions of shared bands produced by the SSR primers (Jaccard, 1908). Similarity matrix was generated using the NTYSYS-pc 2.20 L software (Rholf, 2005). The resultant similarity matrix was employed to construct dendrogram using sequential agglomerative hierarchical nesting (SAHN) based on unweighted pair group method with arithmetic mean (UPGMA) to infer genetic relationship and phylogeny (Mathew et al., 2000). In addition, Principal Component Analysis (PCA) based clustering was done using software MINITAB version 15.0, in order to visualize the difference between the individuals graphically in 2D image.
Marker and Morphological Trait Association
Single marker analysis was performed by one way ANOVA to identify SSR markers associated with resistance to salinity. A significant F test (p<0.01) indicated association of marker locus with phenotypic trait.
Multiple regression analysis was worked out to detect the marker-trait association. The physiological traits served as dependent variables and the SSR marker data as an independent variable. Multiple regression analysis was performed using a software MINITAB version 15.0 for the subset of markers that had significant correlation with the trait on an individual basis.
RESULTS AND DISCUSSION
Genetic Diversity Analysis Using Molecular Markers
In the present study, 25 rice genotypes including check varieties were evaluated for association of genetic diversity with salinity tolerance using Simple Sequence Repeats (SSR) markers.
Effect of Salinity on Ion Concentration in Plant Tissues (mg g-1)
The ratio between the shoot sodium concentration and shoot potassium concentration was also calculated for all the rice genotypes in order to classify them into salt tolerant, moderately tolerant and salt susceptible lines (Table 1), the Na+/ K+ ratio values found to be lower for salt tolerant lines and higher for salt susceptible lines. The genotypes, FL 478, Pokkali (salt tolerant) and IR 29 (salt susceptible) were used as checks. The range for Na+/ K+ ratio varies from 0.67 (IR 72593-B-19-2-3-1) to 0.98 (IR 29).
SSR Analysis
To assess the quality, all the genomic DNA samples were run on 0.8% agarose gel and the gel was stained with ethidium bromide and the quantity of DNA present in each sample was determined by reading the absorbance at 260 nm in a Hoefer Dyna Quant 200 fluorometer. The quantity of DNA in different samples varied from 100 - 600 ng μL-1.
| Table 1: | Na+/K+ content (mg g-1) in shoots of rice genotypes after 7 days of salinization |
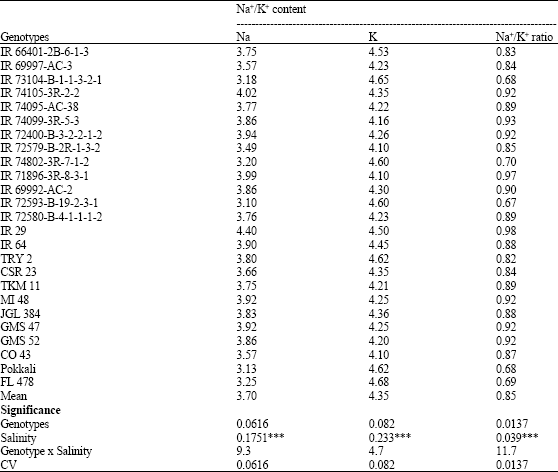 | |
| ***Significant at p<0.001% | |
This variation was due to difference in the quantity and quality of leaf sample taken for DNA extraction. After quantification, all the samples were diluted to 25 ng μL-1, using sterile distilled water and used for PCR reactions.
The 5 SSR primer pairs used in the present study produced scorable, unambiguous markers. The details of markers amplified by the 5 SSR primer pairs among 25 rice genotypes are given in Table 2, a total of 5 loci were amplified and found polymorphic. Polymorphism percentage was 100%. The number of alleles produced by different primers was 10 with an average 2 alleles per primer. All the primers were proved to be highly polymorphic (100.0%). The SSR products size ranged from 136 to 209 bp. The SSR marker profiles of 25 rice genotypes generated by the primer RM 9, 23, 212, 493 and 8053 are given in Fig. 1 and 2, respectively.
Cluster Analysis
The similarity index values were obtained for each pair wise comparison among the 25 rice genotypes. The similarity coefficients based on 5 SSR loci ranged from 0.1 to 1.00. Among the 25 rice genotypes the highest similarity index (1.000) was observed between IR 72400 and 66401. Similarly between GMS 47, 52 and JGL 384. The genotype IR 74802 and 72593 had higher similarity index. The lowest similarity index (0.10) was observed between IR 73104 with 72580, 29, 64, GMS 47 and 52 and CO 43.
| Table 2: | List of SSR primer pairs showed polymorphism |
 | |
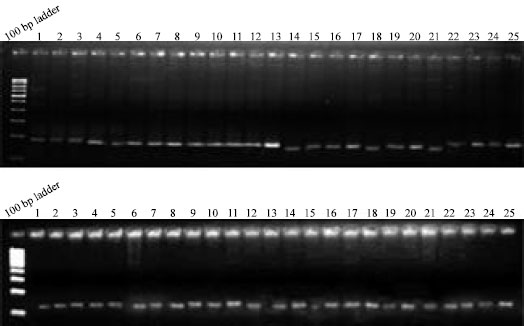 | |
| Fig. 1: | SSR marker profile of rice genotypes generated by the primer RM 9 and 23 |
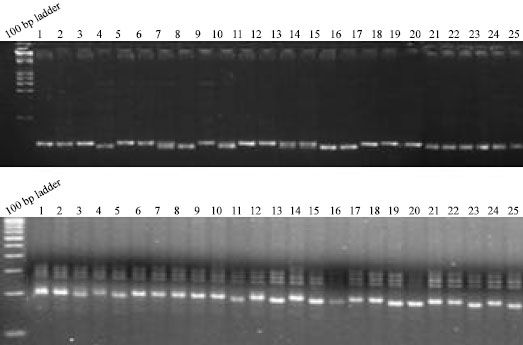 | |
| Fig. 2: | SSR marker profile of rice genotypes generated by the primer RM 493 and 8053 |
Clusters based on Dendrogram
The SSR marker data were subjected to cluster analysis using NTSYS program for 25 genotypes, to estimate similarity indices and genetic relatedness among the genotypes. The Similarity Index (SI) values were computed as a ratio of number of similar bands to the total number of bands in pair wise comparison of the accessions. The SI values were obtained for each pair wise comparison of SSR following the UPGMA method using NTSYS-pc 2.20L software program (Rholf, 2005). Option SAHN was performed which resulted in the dendrogram. To estimate the genetic relatedness among the 25 rice genotypes, similarity analysis was done and dendrogram showing the genetic relatedness among the 25 rice genotypes was constructed (Fig. 3).
The 25 rice genotypes were grouped into eight major clusters (Table 3), cluster I was the largest with twelve genotypes, the cluster VII comprised of 4 genotypes with all tolerant viz., IR 73104, 74802, 72593 and FL 478, the cluster II, V and VI comprised of two genotypes each, the cluster III, IV and VIII comprised of one genotype (Pokkali). The 5 SSR marker profiles were generated among the 25 rice genotypes and finally the amplified products were analyzed. The similarity matrix was computed using SSR markers based on Jaccard’s coefficient following the UPGMA method using SAHN programme of NTSYS-pc. This matrix was used for cluster and principal component analysis. The Jaccard’s similarity coefficient for the SSR data set varied from 0.10 to 1.000. The SSR marker profile resulted in eight clusters. The cluster I and II was flocked with salt susceptible accessions and the cluster VII and VIII with salt tolerant genotypes IR 72593, IR74802 and IR73104 along with salt tolerant FL 478 and Pokkali.
Principal Component Analysis
PCA showed the distribution and grouping of rice genotypes. The two dimensional ordination of the 25 rice genotypes confirmed the clustering pattern obtained by the cluster analysis. PCA analysis was more informative to analyze the variation between salt tolerant genotypes and susceptible genotypes. Inspite of it, a few more clusters were also seen, which deviated from both the salt tolerant and susceptible clusters (Fig. 4).
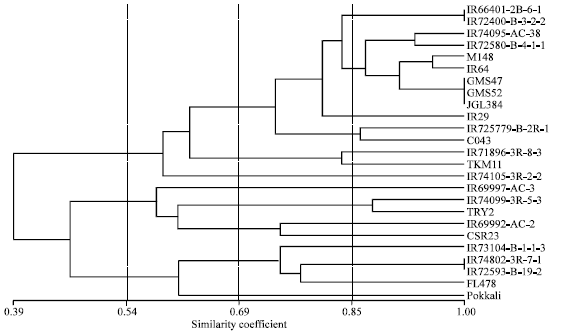 | |
| Fig. 3: | Dendrogram constructed using SSR marker data based on Jaccard’s similarity index values |
| Table 3: | Cluster composition of rice genotypes for SSR marker |
 | |
The Association of SSR Markers with Physiological Traits Single Factor ANOVA Analysis and Multiple Regressions Analysis
The five polymorphic loci, which were significantly associated with Na+/K+ ratio QTL by multiple regression. Among the five polymorphic loci, only three loci (RM 23, RM 493 and RM 8053) were found to be significantly associated with Na+/K+ ratio QTL by Single marker analysis and multiple regressions, the markers that were found to be associated with Na+/K+ ratio QTL in multiple regression analysis are presented in Table 4.
An attempt has been made to determine the association between the SSR markers and Na+/K+ ratio for salt tolerance as early reported by Bonilla et al. (2002) and Lin et al. (2004) using different rice genotypes. The genetic variability for salt tolerance among 25 rice genotypes was determined using the physiological trait viz., shoot sodium and potassium ratio. Analyzing for salt tolerance especially in shoots is appropriate because most Na+ that is delivered to shoots and remains there (Munns and Tester, 2008) and from this study it is clearly evidence that Na+ accumulated and deposited in shoots, especially in leaves rather than roots. Salt tolerance of a crop depends on Na+ and K+ uptake, ion balance and ion compartmentation (Yeo and Flowers, 1986).
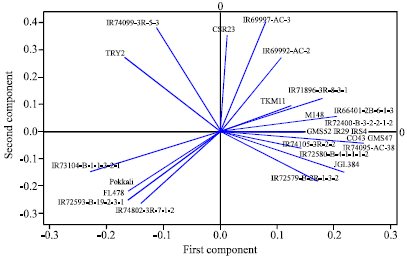 | |
| Fig. 4: | Pattern of relationship among the rice genotypes revealed by PCA analysis based on SSR marker data |
| Table 4: | Analysis of variance involving multiple regressions for physiological trait using SSR markers |
| **Significant at 5 and 1%, x: Regression, y: Residual, df: Degree of freedom | |
Alignment of genotypes in closely associated clusters is related to tolerance level with similar ion concentration. It is evident that genotypes with higher level of tolerance have lower level of Na+/K+ ratio and clustered into same group. Some of the breeding lines derived from IRRI shows tolerance at all level of salt treatments and rest of them were sensitive or moderately tolerant in saline solution. This may be due to the source and genetic backgrounds of the materials were it is derived. It is clear that adaptation of rice to saline soils is different among genotypes with diverse genetic backgrounds (Zeng et al., 2004). The rice genotypes at the extreme clusters can be used in breeding programme to generate high variability. There is clear evidence that genetic diversity was the positively correlated and associated with microsatellite marker for Na+/K+ ratio.
The SSR variation in relation to salt tolerance was found to be more among the rice genotypes. To confirm the clustering pattern, the principal component analysis was followed. PCA was employed to exploit the resolving power of ordination. The two- dimensional ordination of the rice genotypes confirmed the clustering pattern obtained by the cluster analysis. Clear distinction of the salt tolerant and susceptible rice accessions and varieties were evident in two-dimensional ordination. The phenotypic variations for Na+/K+ ratio do exist among the rice accessions used. The salt tolerant genotypes Pokkali, FL 478, IR 72593, IR 74802 and IR 73104 remained healthy without exhibiting salt stress symptoms after being given 120 mM NaCl salt stress and it was clearly evident from the low shoot Na+/K+ ratio. Na+/K+ ratio was low in the salt tolerant accessions and high among the susceptible lines. The mechanism behind the tolerant genotypes under saline condition was associated with Na+ exclusion and increased absorption of K+ to maintain a optimum Na+/K+ tolerance ratio in the shoots. It is considered that damage of leaves was attributed to the accumulation of Na+ from the root to the shoot in high concentration of salt stress (Lin et al., 2004). Vigorous growth, tall stature and tolerance had the same level of Na+ transportation as like the stunted susceptible genotype, but had much lower shoot Na+ concentration. The susceptible genotypes noticed stunted growth, rolling of leaves, white leaf tips, drying of older leaves and ultimately death of plants without further recovery. This was consistent with the results of Koyama et al. (2001). Low Na+/K+ ratios at the seedling stage are associated with tolerance and the possible reason for this can be Na+ exclusion as well as partitioning of Na+ into older leaves (Flowers and Yeo, 1995; Moradi et al., 2003; Golldack et al., 2002; Lee et al., 2003; Ren et al., 2005). Thus Na+/K+ ratio increased tremendously with rise of salt concentration and leaf age, the tolerant and susceptible genotypes responds differently.
The polymorphic SSR markers showed a clear cut differentiation of tolerance based on the Na+ and K+ uptake ratio. Gregorio et al. (2002) have mapped a major QTL designated saltol on chromosome 1 (flanked by SSR markers RM 23 and RM 140) using a population generated from a cross between salt sensitive IR 29 and salt tolerant Pokkali. This QTL accounted for more than 70% of the variation in salt uptake (Bonilla et al., 2002) and is now being mapped to within 1 cM distance using a large set of near isogenic lines (NILs). Pokkali was the source of positive alleles for this major QTL (Bonilla et al., 2002), which accounted for high K+ and low Na+ adsorptions and low Na+ to K+ ratio under salinity stress (Gregorio et al., 2002). Lin et al. (2004) reported that the existence of several QTLs for survival days of the seedlings, root potassium concentration, root sodium concentration, shoot sodium concentration and shoot potassium concentration under salt stress.
The association analysis was performed for the physiological trait Na+/K+ ratio with SSR markers following single marker analysis and multiple regressions. The regression analysis was capable of identifying those markers, which showed a strong association with QTLs. The markers used in the present study were selected from Na+/K+ ratio QTL region on chromosome 1 (Bonilla et al., 2002; Lin et al., 2004). Five markers were selected from Na+/K+ ratio QTL, which generated 5 polymorphic loci. Na+/K+ ratio was regressed with each of the 5 polymorphic loci. The single factor ANOVA and multiple regression analysis revealed that three loci (RM 23, RM 493 and RM 8053) were significantly associated with Na+/K+ ratio QTL.
CONCLUSION
Hybridization between these diverse lines with salt tolerance from the genetic clusters into elite cultivars will maximize tolerance and also enhance variation to create new breeding population. Although, salt tolerance is not controlled by single gene and vary with growth stage and by utilization of tightly linked molecular markers for Na+/K+ ratio may facilitate better differentiation of tolerance at early growth stage. The associated SSR markers can be regarded as the most reliable markers for marker-assisted selection to improve salinity tolerance in rice. These markers can also be used to screen large germplasm collection of rice to identify more salt tolerant rice genotypes, which can be utilized as donors in the breeding programme for generating salt tolerant varieties.
REFERENCES
- Ashraf, M. and J.W. O'Leary, 1996. Responses of newly developed salt tolerant genotype of spring wheat to salt stress: Yield components and ion distribution. Agron. Crop Sci., 176: 91-101.
Direct Link - Bonilla, P.S., J. Dvorak, D. Mackill, K. Deal and G. Gregorio, 2002. RFLP and SSLP mapping of salinity tolerance genes in chromosome 1 of rice (Oryza saliva L.) using recombinant inbred lines. Philippines Agric. Sci., 85: 64-76.
Direct Link - Dellaporta, S.L., J. Wood and J.B. Hicks, 1983. A plant DNA minipreparation: Version II. Plant Mol. Biol. Rep., 1: 19-21.
CrossRefDirect Link - Flowers, T.J., M.L. Koyama, S.A. Flowers, C. Sudhakar, K.P. Singh and A.R. Yeo, 2000. QTL: Their place in engineering tolerance of rice to salinity. J. Exp. Bot., 51: 99-106.
Direct Link - Flowers, T.J. and A.R. Yeo, 1995. Breeding for salinity resistance in crop plants: Where next? Aust. J. Plant Physiol., 22: 875-884.
CrossRefDirect Link - Garland, S.H., L. Lewin, M. Abedinia, R. Henry and A. Blakeney, 1999. The use of microsatellite polymorphisms for the identification of Australian breeding lines of rice (Oryza sativa L.). Euphytica, 108: 53-63.
CrossRefDirect Link - Golldack, D., H. Su, F. Quigley, U.R. Kamasani and C. Munoz-Garay et al., 2002. Characterization of a HKT-type transporter in rice as a general alkali cation transporter. Plant J., 31: 529-542.
PubMed - Gregorio, G.B., D. Senadhira, R.D. Mendoza, N.L. Manigbas, J.P. Roxas and C.Q. Guerta, 2002. Progress in breeding for salinity tolerance and associated abiotic stresses in rice. Field Crop Res., 76: 91-101.
CrossRefDirect Link - Jaccard, P., 1908. Nouvelles recherches sur la distribution florale. Bull. Soc. Vaud. Sci. Nat., 44: 223-270.
CrossRefDirect Link - Kawasaki, S., C. Borchert, M. Deyholos, H. Wang and S. Brazille et al., 2001. Gene expression profiles during the initial phase of salt stress in rice. Plant Cell., 13: 889-906.
PubMedDirect Link - Koyama, M.L., A. Levesley, R.M.D. Koebner, T.J. Flowers and A.R. Yeo, 2001. Quantitative trait loci for component physiological trait determining salt tolerance in rice. Plant Physiol., 125: 406-422.
Direct Link - Lee, K.S., W.Y. Choi, J.C. Ko, T.S. Kim and G.B. Gregorio, 2003. Salinity tolerance of japonica and indica rice (Oryza sativa L.) at the seedling stage. Planta, 216: 1043-1046.
Direct Link - Lee, S.Y., J.H. Ahn, Y.S. Cha, D.W. Yun and M.C. Lee et al., 2007. Mapping QTLs related to salinity tolerance of rice at the young seedling stage. Plant Breed., 126: 43-46.
Direct Link - Lin, H.X., M.Z. Zhu, M. Yano, J.P. Gao and Z.W. Liang et al., 2004. QTLs for Na+ and K+ uptake of the shoots and roots controlling rice salt tolerance. Theor. Applied Genet., 108: 253-260.
PubMed - Munns, R., R.A. James and A. Lauchli, 2006. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot., 57: 1025-1043.
CrossRefPubMedDirect Link - Munns, R. and M. Tester, 2008. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol., 59: 651-681.
CrossRefDirect Link - Ozturk, Z.N., V. Talame, M. Deyhols, C.B. Michalowski and D.W. Galbraith et al., 2002. Monitoring large-scale changes in transcript abundance in drought and salt-stressed barley. Plant Mol. Biol., 48: 551-573.
Direct Link - Prasad, S.R., P.G. Bagali, S. Hittalmani and H.E. Shaahidhar, 2000. Molecular mapping of quantitative trait loci associated with seedling tolerance to salt stress in rice (Oryza sativa L.). Curr. Sci., 78: 162-164.
Direct Link - Prasad, K.V.S.K., P. Sharmila, P.A. Kumar and P.P. Saradhi, 2000. Transformation of Brassica juncea (L.) Czern with bacterial codA gene enhances its tolerance to salt stress. Mol. Breed., 6: 489-499.
CrossRef - Ren, Z.H., J.P. Gao, L.G. Li, X.L. Cai and W. Huang et al., 2005. A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat. Genet., 37: 1141-1146.
CrossRef - Sambrook, J., E.F. Frotsch and T. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor, Laboratory press, New York, United States, ISBN: 9780879693091 Pages: 1659.
Direct Link - Seki, M., M. Narusaka, J. Ishida, T. Nanji and M. Fujita et al., 2002. Monitoring the expression profiles of 7000 Arabidopsis genes under drought, cold and high-salinity stresses using a full-length cDNA microarray. Plant J., 31: 279-292.
Direct Link - Takehisa, H., T. Shimodate, Y. Fukuta, T. Ueda and M. Yano et al., 2004. Identification of quantitative trait loci for plant growth of rice in paddy field flooded with salt water. Field Crop Res., 85: 85-95.
Direct Link - Yeo, A.R. and T.J. Flowers, 1986. Salinity resistance in rice (Oryza sativa L.) and pyramiding approach to breeding for saline soils. Aust. J. Plant Physiol., 13: 161-173.
CrossRef - Zhang, G.Y., Y. Guo, S.L. Chen and S.Y. Chen, 1995. RFLP tagging of a salt tolerance gene in rice. Plant Sci., 110: 227-234.
Direct Link - Zeng, L., T.R. Kwon, X. Liu, C. Wilson, C.M. Grieve and G.B. Gregorio, 2004. Genetic diversity analyzed by microsatellite markers among rice (Oryza sativa L.) genotypes with different adaptations to saline soils. Plant Sci., 166: 1275-1285.
CrossRefDirect Link