
S. M.T. Mohammed
Department of Horticulture, Faculty of Agriculture, Tishreen University, Latakia, Syria
M. Humidan
Department of Horticulture, Faculty of Agriculture, Tishreen University, Latakia, Syria
M. Boras
Department of Horticulture, Faculty of Agriculture, Tishreen University, Latakia, Syria
O. A. Abdalla
Department of Plant Protection, College of Food and Agricultural Sciences, King Saud University, Riyadh, Saudi Arabia
International Journal of Agricultural Research
Year: 2009 | Volume: 4 | Issue: 11 | Page No.: 362-369







ABSTRACT
The objective of this study was to determine the impact of interaction between rootstocks and scions of watermelon and tomato on the chemical contents of their leaves and roots. The rootstock of local Syrian tomato produced the highest amount of total lipids, total fatty acids percentage and total unsaponated percentage of total lipids in root (0.69, 92.39, 3.34%) and leaf (0.73, 91.54, 4.02%) compared to Beaufort rootstock and He-man rootstock when grafted with Cecilia scion. With regard to watermelon grafts Samara on C. pepo excelled the two other watermelon grafts namely, Samara on Tetsukauto and Samara on Lagenaria siceria in its overall root contents of total fatty acids percentage of total lipids and total unsaponated percentage of total lipids. Samara on C. pepo excelled the other two watermelon grafts with regards to its significant leaves (0.1326, 18.73%) and root contents (0.1214, 15.33%) of phospholipids percentage and total unsaponoted phospholipids, respectively. Grafting increased indole-3-acetic acid (IAA) in seedlings of both tomato and watermelon. Cecilia on Beaufort (32.43 nm g-1) and Cecilia on Syria (36.71 nm g-1) had significantly greater contents of Indole-3-acetic acid in their roots whereas, only Samara on C. pepo watermelon grafts (98.27 nm g-1) had significant greater contents of IAA in its roots. Clear effects of grafting watermelon and tomato on their lipids and IAA contents were concluded in this study.
PDF Abstract XML References Citation
How to cite this article
S. M.T. Mohammed, M. Humidan, M. Boras and O. A. Abdalla, 2009. The Role of Grafting Tomato and Watermelon on Different Rootstocks on Their
Chemical Contents. International Journal of Agricultural Research, 4: 362-369.
DOI: 10.3923/ijar.2009.362.369
URL: https://scialert.net/abstract/?doi=ijar.2009.362.369
DOI: 10.3923/ijar.2009.362.369
URL: https://scialert.net/abstract/?doi=ijar.2009.362.369
INTRODUCTION
The production of grafted plants first began in Japan and Korea in late 1920, with watermelon grafted on gourd rootstock. There were many grafting methods for different types of fruit-bearing vegetables, which have been used in greenhouses as well as in the open-field agriculture (Khah et al., 2006; Pogonyi et al., 2005; Augustin et al., 2002; Mohammed et al., 2009). In addition to the many advantages indicated for grafting in the previous references, enhancement of nutrient uptake is one of the preferable advantage of this technique (Takacs et al., 2005). Good combination between rootstocks and scions results in increase of fruit yield and quality (Romano and Paratore, 2001).
Chemical analysis of leaves and roots of tomato and watermelon for lipids and hormones plays an important role for determination of the quality of some scions on different rootstocks. Many scientists found an increase in the concentration of fatty acids and some changes in lipids during the growth periods of different plants (De La Roche et al., 1972; Gerloff et al., 1967; Smith, 1968; Willemot, 1975). Roberts et al. (2005) stated that using grafting technique led to improve the vegetative and root growth of watermelon.
The objective of this study was to investigate the effect of different rootstock/scion combinations of tomato and watermelon on the chemical contents of leaves and roots of each of these two plant species.
MATERIALS AND METHODS
Tomato (Lycopersicon esculentum Mill.) var. Cecilia F1 was grafted on three rootstocks, Beaufort, He-man and local Syrian tomato using tube grafting method. Watermelon (Citrullus lanatus [thomb.]) var. Samara F1 was grafted on Tetsukabuto squash, Lagenaria siceraria Standl. and local Syrian squash Cucurbita pepo L. by tongue approach grafting method. Twenty grafts of each of tomato and watermelon were prepared. Seeds of three rootstocks of tomato plants were sown 10 days before the planting of the scion, Cecilia F1. The three rootstocks used in this study were Beaufort (Lycopersicon esculentum x Lycopersicon hirsutum) from De Ruiter seeds, He-man (L. esculentum x L. hirsutum ) from Syngenta seeds B.V. and local Syrian seeds from Syrian farmers. Cecilia F1 seeds were obtained from GSN French Company. All scion seeds were sown on 02/05/2007 in 96 holes trays filled with peat moss substrate inside a fiberglass greenhouse in ASTRA Farms in Tabuk, Kingdom of Saudi Arabia. Grafting was done 18 days after planting of the scion using tube grafting technique. The grafted plants were maintained under transparent plastic cover at 28°C and around 90% humidity. Excellent union of scion with each of the three rootstocks was established 9 days after grafting.
Seeds for the scion, Samara F1 from SAKATA Vegetables Europe French Company, were sown 10 days before seeding of the watermelon rootstocks on 22/04/2007. The three rootstocks used were Tetsukabuto (C. maxima x C. moschata) from Takii Japanese Company, bottle gourd Lagenaria ciceraria and local Syrian squash Cucurbita pepo. Tongue approach grafting method was used.
Five homogeneous grafts were statistically analyzed for each of tomato and watermelon. Following recording of measurements, lipids were analyzed using Sokslet apparatus according to AOAC technique (1995). The percentage of total saturated and unsaturated fatty acids were determined in roots and leaves of both tomato and watermelon seedlings using lead-Salt-ether method. Saponification was performed for oil samples and then percentage of unsaponified substances and total fatty acids were calculated by weight according to Farag et al. (1986). Being an important natural auxin which has several different effects such as induction of cell elongation and cell division, indole-3-acetic acid (IAA) was extracted by 80% methanol according to Zaffase et al. (2002).
Descriptive statistics for the studied characteristics were expressed as percentage. Pooled data were analyzed using the statistical analysis SPSS 13. One-way ANOVA was used to determine differences between treatments. Duncan's multiple range test and LSD. (p = 0.05) were utilized to distinguish between means (Steel and Torrie, 1980).
RESULTS AND DISCUSSION
Significantly, greater amounts of total lipids, total fatty acids percentage of total lipids and total unsaponated substances percentage of total lipids were found in roots of the tomato grafts, Cecilia on Beaufort and Cecilia on Syria (Table 1, Fig. 1). No such significant increase in any of these three substances was found in the roots of Cecilia on He-man. This increase in the level of lipids percentage is attributed to grafting, since, the lipids percentage in both Cecilia hybrid (0.53%) and in Beaufort without grafting (0.51%) was lower than the lipids percentage in grafts Cecilia on Beaufort (0.63%).
In watermelon, roots of grafts Samara on Lageneria siceria and Samara on C. pepo showed significantly greater increase in total fatty acids percentage of total lipids (Table 1, Fig. 2), however, root of graft Samara on Testukabuto and Samara on C. pepo showed significantly greater increase in total unsaponated substances percentage of total lipids.
| Table 1: | Root content of total lipids, total fatty acids of total lipids and total unsap percentage of total lipids in tomato and watermelon seedling |
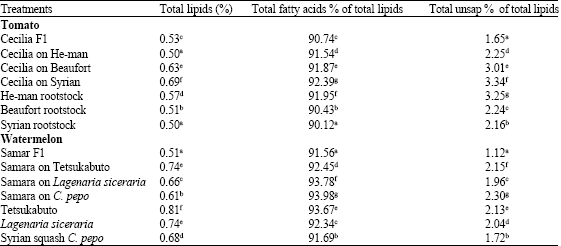 | |
| Means in the same column followed by the same letter(s) are not significantly different (p≤0.05) according to Duncan's multiple range test | |
 | |
| Fig. 1: | Grafting of Cecilia F1 tomato on three different rootstocks (Syrian tomato , He-man and Beaufort) |
 | |
| Fig. 2: | Grafting of Samara F1 watermelon on three different rootstocks (Tetsukabuto, Lagenaria siceraria and C. pepo) |
| Table 2: | Leaves content of total lipids percentage, total fatty acids percentage of total unsap. percentage of total lipids in tomato and watermelon seedlings |
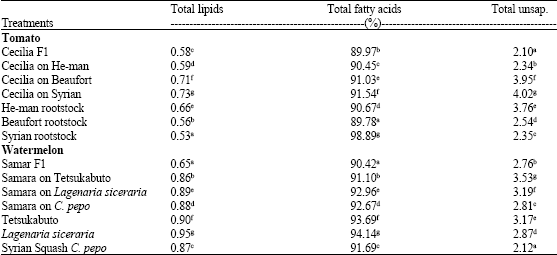 | |
| Means in the same column followed by the same letter(s) are not significantly different (p≤0.05) according to Duncan's multiple range test | |
These results agreed with Horvath et al. (1980) and Bulder et al. (1991). However, Ruiz et al. (2005) stated that grafting gives lowest lipid peroxidation (which it most often affects polyunsaturated fatty acids) in tobacco plants and Li et al. (2008) reported that upon exposure to chilling stress, the malondialdehyde (MDA) content, which reflects the degree of lipid peroxidation, increased markedly in the own-rooted seedling roots and leaves and kept stable in the grafted cucumber seedlings.
Leaves of all three tomatoes and watermelon grafts revealed significant increase in total lipids, total fatty acids percentage and total unsaponated substances compared to the control (Table 2).
Significant increase in phospholipids and total unsaponated percentage of phospholipids was detected in leaves and roots of all tomato and watermelon grafts (Table 3, 4).
The highest phospholipids percentage and total unsaponated percentage of phospholipids was found in tomato plants grafted on local Syrian tomato (0.104 and 22.27%, respectively) as shown in Table 4.
| Table 3: | Total phospholipids % and unsap. of phospholipids in leaves of tomato and watermelon seedlings |
 | |
| Means in the same column followed by the same letter(s) are not significantly different (p≤0.05) according to Duncan's multiple range test | |
| Table 4: | Total phospholipids percentage and unsap. of phospholipids in roots of tomato and watermelon seedlings |
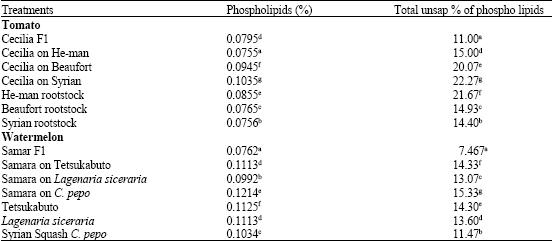 | |
| Means in the same column followed by the same letter(s) are not significantly different (p≤0.05) according to Duncan's multiple range test | |
Phospholipids are considered very vital in cellular structure especially in cellular membranes. Phospholipids and total unsaponated percentage of phospholipids significantly increased in the root of all watermelon grafts (Table 4) however, Samara on C. pepo showed the highest level of increase in both phospholipids percentage and total unsaponated percentage of phospholipids (0.121 and 15.33%, respectively).
Significant increase of total unsaturated fatty acids % of total lipids was detected in leaves of the tomato grafted Cecilia on Beaufort and Cecilia on Syria whereas, significant increase of total saturated fatty acids % of total lipids was detected in the leaves of Cecilia on He-man compared with the other two grafts (Table 5). However, significant decrease of the later was observed in the leaves of all watermelon grafts compared with the control (Table 5).
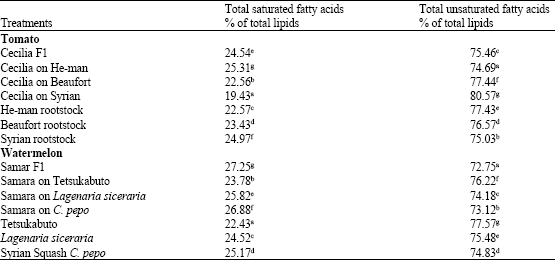
Total Unsaturated fatty acids of total lipids significantly increased in the roots of the tomato grafts Cecilia on Beafourt and Cecilia on Syria (77.44 and 80.57%, respectively), whereas, total saturated fatty acids % of total lipids significantly increased in the roots of Cecilia on He-man only 25.31% (Table 6). Significant increase of total unsaturated fatty acids % of total lipids occurred in the roots of Cecilia on Beaufort and Cecilia on local Syrian tomato but not in roots of Cecilia on He-man, however, the reverse is true in the case of total saturated fatty acids % of total lipids.
| Table 5: | Total saturated and unsaturated fatty acids % in leaves of tomato and watermelon plants |
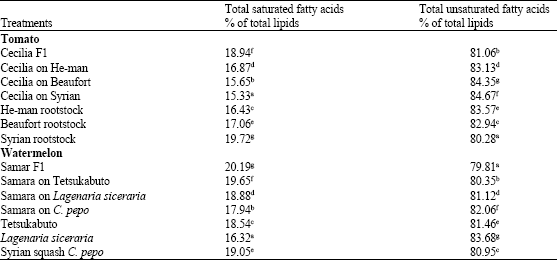 | |
| Means in the same column followed by the same letter(s) are not significantly different (p≤0.05) according to Duncan's multiple range test | |
| Table 6: | Total saturated and unsaturated fatty acids % in roots of tomato and watermelon plants |
 | |
| Means in the same column followed by the same letter(s) are not significantly different (p≤0.05) according to Duncan's multiple range test | |
These results agreed with Markhart-III et al. (1980), Horvath et al. (1980) and Bulder et al. (1991). Although, it seems clearly that scion/rootstock relationship had a role in total saturated and unsaturated fatty acids % of total lipids contents, its affect on the later is considered more important since, it is reported to be involved in cold resistance.
Significantly greater increase of indole-3-acetic acid occurred in the roots of the tomato grafts Cecilia on Beaufort and Cecilia on local Syrian tomato (32.43 and 36.71%, respectively). Although, the hormone amount in these two rootstocks was less than in the scion (Cecilia F1) and high in the grafts, this may be attributed to the interaction between the rootstock and the scion of these grafts. The IAA hormone is the only natural product that promotes or inhibits the growth of roots, buds and stems depending on its concentration, where, the roots are sensitive to its high concentration and in all three watermelon grafts compared to their control treatments (Table 7). Graft Samara on C. pepo revealed a remarkably higher value of IAA content compared to the other two grafts.
| Table 7: | The roots content of Indo-3-acetic acid (IAA) in seedlings of tomato and watermelon |
 | |
| Means in the same column followed by the same letter are not significantly different (P=0.05) according to Duncan's multiple range test | |
CONCLUSION
From the previous results, it can be concluded that chemical evaluation is considered one of the important factors in evaluating success of the grafting process in tomato and watermelon plants. Thus, this will probably make rootstocks selection for vegetables grafting much easier in the future. To improve the application of grafting, farmers and agricultural specialists should always be careful in selecting suitable rootstocks.
REFERENCES
- Augustin, B., V. Graf and N. Laun, 2002. Temperature influencing efficiency of grafted tomato cultivars against root-knot nematode (Meloidogyne arenaria) and corky root (Pyrenochaeta lycopersici). Zeitschrift Fur Pflanzenkrankheiten Und Pflanzenschutz, 109: 371-383.
Direct Link - Bulder, H.A.M., A.P.M. Den-Nijs, E.J. Speek, P.R. van Hasselt and P.J.C. Kuiper, 1991. The effect of low root temperature on growth and lipid composition of low temperature tolerant rootstock genotypes for cucumber. J. Plant Physiol. 138: 661-666.
Direct Link - De La Roche, I.A., C.J. Andrews, M.K. Pomeroy, P. Weinberger and M. Kates, 1972. Lipid changes in winter wheat seedlings (Triticum aestivum)at temperatures inducing cold hardines. Can. J. Bot., 50: 2401-2409.
CrossRefDirect Link - Gerloff, E.D., T. Richardson and M.A. Stahmann, 1966. Changes in fatty acids of alfalfa roots during cold hardening. Plant Physiol., 41: 1280-1284.
CrossRefDirect Link - Farag, R.S., S.A.S. Hallabo, F.M. Hewedi and A.E. Bosyony, 1986. Chemical evaluation of Rape seed. Fotte- Seifen Anstrichmittel., 88: 391-397.
CrossRefDirect Link - Horvath, I., L. Vigh, A. Belea and T. Farkas, 1980. Hardiness dependent accumulation of phospholipids of wheat cultivars. Physiol. Plant, 49: 117-120.
Direct Link - Khah, E.M., E. Kakava, A. Mavromatis, D. Chachalis and C. Goulas, 2006. Effect of grafting on growth and yield of tomato (Lycopersicon esculentum Mill.) in greenhouse and open-field. J. Applied Hortic., 8: 3-7.
Direct Link - Li, J.Y., H.X. Tian, X.G. Li, J.J. Meng and Q.W. He, 2008. Higher chilling-tolerance of grafted cucumber seedling leaves upon exposure to chilling stress. Agric. Sci. China, 7: 570-576.
CrossRef - Markhart-III, A.H., M.M. Peet, N. Sionit and P.J. Kramer, 1980. Low temperature acclimation of root fatty acid composition, leaf water potential gas exchange and growth of soybean seedlings. Plant Cell Environ., 3: 435-441.
Direct Link - Mohammed, S.M.T., M. Humidan, M. Boras and O.A. Abdalla, 2009. Effect of grafting tomato on different rootstocks on growth and productivity under glasshouse conditions. Asian J. Agric. Res., 3: 47-54.
CrossRefDirect Link - Pogonyi, A., Z. Pek, L. Helyes and A. Lugasi, 2005. Effect of grafting on the tomato's yield, quality and main fruit components in spring forcing. Acta Aliment., 34: 453-462.
Direct Link - Ruiz J.M., J.J. Rios, M.A. Rosales, R.M. Rivero and, L. Romero, 2005. Grafting between tobacco plants to enhance salinity tolerance. J. Plant Physiol., 163: 1229-1237.
Direct Link - Smith, D., 1968. Varietal chemical differences associated with the freezing resistance in forage plants. Cryobiology, 5: 148-159.
Direct Link - Steel, R.G.D. and J.H. Torrie, 1980. Principles and Procedures of Statistics: A Biometrical Approach. 2nd Edn., McGraw Hill Book Co., New York, USA., ISBN-13: 9780070609266, Pages: 633.
Direct Link - Takacs, T., I. Biro, A. Anton and H. Chaoxing, 2005. The effect of AMF inoculation on growth ant nutrient uptake of tomato. Cereal Res. Commun., 1: 125-128.
Direct Link - Willemot, C., 1975. Stimulation of phospholipids biosynthesis during frost hardening of winter wheat. Plant Physiol., 55: 356-359.
Direct Link - Zaffase, G.R., L.E.P. Peres, F.A. Tcacenco and G.B. Kerbauy, 2002. Indol-3-acetic acid metabolism in normal and dwarf micropropagated banana plants (Musa spp. AAA). Braz. J. Plant Phisiol., 14: 211-217.
Direct Link