
A.M. Roslan
Department of Bioprocess Technology, Faculty of Biotechnology and Biomolecular Sciences, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
M.A. Hassan
Department of Bioprocess Technology, Faculty of Biotechnology and Biomolecular Sciences, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
S. Abd-Aziz
Department of Bioprocess Technology, Faculty of Biotechnology and Biomolecular Sciences, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
P.L. Yee
Department of Bioprocess Technology, Faculty of Biotechnology and Biomolecular Sciences, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
International Journal of Agricultural Research
Year: 2009 | Volume: 4 | Issue: 5 | Page No.: 185-192







ABSTRACT
In this study, Palm Oil Mill Effluent (POME) was selected as supplement in rice straw fermentation to replace commercial inducer due to its high concentration of simple carbohydrate and nitrogenous compounds. The cellulase activity of the enzyme produced were then determined individually and combined (enzyme cocktail) to observe increment in cellulase activity. Individually, filter paperase (FPAse), carboxymethyl cellulase (CMCase) and β-glucosidase activities increase for all 3 fungi used except FPAse for Aspergillus sp. (NEW). Highest FPAse and CMCase activities increment observed was from Phanerochaete chrysosporium which were 72 and 236% increment, respectively, while highest β-glucosidase activity was from Aspergillus terreus (AT) which was 298% increment. Meanwhile in cocktail cellulase, synergistic effect was observed especially for FPAse activity whereby highest FPAse activity observed was by cocktail of all the three species of fungi which show 393% increment. Though CMCase increment was observed in this mode, it was not as high as FPAse where the highest CMCase activity observed was cocktail by the three fungal species which increased by 41%. However, in β-glucosidase activity, only cocktail of NEW and AT showed increment from their average activity which was by 6%. It seems that β-glucosidase activity plays major role in determining the total cellulase activity. In the nutshell, supplementation of POME can improve cellulase activity.
PDF Abstract XML References Citation
How to cite this article
A.M. Roslan, M.A. Hassan, S. Abd-Aziz and P.L. Yee, 2009. Effect of Palm Oil Mill Effluent Supplementation on Cellulase Production from Rice Straw by Local Fungal Isolates. International Journal of Agricultural Research, 4: 185-192.
DOI: 10.3923/ijar.2009.185.192
URL: https://scialert.net/abstract/?doi=ijar.2009.185.192
DOI: 10.3923/ijar.2009.185.192
URL: https://scialert.net/abstract/?doi=ijar.2009.185.192
INTRODUCTION
The world’s fossil fuel supply is nearly depleted. As the fuel price becomes unstable, it is a must to find alternatives for future energy. With the greenhouse gases as a barrier to a clean energy, the best solution is the biofuel. One type of biofuel which is getting huge interest in research is bioethanol. Bioethanol is an octane enhancer (He et al., 2003) and also functions as a carbon-neutral fuel to mitigate global warming (Balat and Balat, 2009). In order to produce a relatively cheap bioethanol, the raw material must be easily obtained and at low cost. In this case, biomass is the most suitable candidate since it is abundant and not fully exploited.
There are huge amount of biomass to be exploited in Malaysia, which were produced during and after processing of the plantation materials. Examples of these biomass are Empty Fruit Bunches (EFB), rice straw, sago hampas, Palm Oil Mill Effluent (POME), POME sludge and sago effluent. With regards to each characteristic and properties, these wastes have potential to be converted into higher value products. For example, rice straw were used for compost (Roca-Perez et al., 2009), EFB were used as mulching and compost (Baharuddin et al., 2009) and POME were used as substrate for biogas production (Sulaiman et al., 2009). Additionally, biomass including rice straw, have potential to be converted into bioethanol (Cardona and Sanchez, 2007).
Rice straw can be converted into bioethanol through 3 stages of processing which is, pretreatment, enzymatic saccharification and fermentation (Moiser et al., 2005; Inoue et al., 2008; Balat et al., 2008). Pretreatment is the first stage to simplify the structure unit of the biomass by physical, thermal or chemical mean. Enzymatic stage includes hydrolysis of the pretreated biomass into sugars which finally can be fermented into ethanol at the third stage. However, the main barrier of this process is high cost of commercial cellulase enzyme being used at the enzymatic hydrolysis stage (Fang et al., 2009). It was suggested to produce the enzyme in-situ and use it directly for the saccharafication process.
Commercial cellulase productions usually utilize high performance or genetically modified fungi such as Aspergillus sp., Trichoderma sp. and Acremonium sp. etc. High activity cellulase production depends on the fungi, substrate, substrate morphology and inducer. Inducers such as thiocellobiose (Rho et al., 1982), cellobiose, CMC (Busto et al., 1995) and lactose (Morikawa et al., 1995) showed drastic increment of cellulase activity. In addition, Wu et al. (2009) reported it is possible to use POME for biotechnological means due to its high concentration of carbohydrate, protein, nitrogenous compounds, lipids and minerals in POME. Therefore, the objective of this study was to determine the effect of POME supplementation on cellulase production from rice straw by using local fungal species. The performance of cellulase activity in individual enzyme and cocktail enzyme was also demonstrated in this study.
MATERIALS AND METHODS
Lab Scale Fermentation
This research was conducted at lab scale in Institute of Bioscience and Biomass Technology Centre, Universiti Putra Malaysia, Malaysia, from January 2008 until December 2008.
Strain Maintenance and Spore Counting
Three species of fungi were used in this study namely Phanerochaete chrysosporium (PC), Aspergillus sp. (NEW) and Aspergillus terreus (AT). PC was obtained from Dr. Zahangir Alam, IIUM, Malaysia, while NEW and AT were obtained from the researcher’s laboratory collection. The individual spores of all the three strains were stored at -20°C in 30% glycerol and grown on Potato Dextrose Agar (PDA) for 7 to 9 days. Spore suspensions were prepared by washing the agar surface with distilled water. Spore counts were determined with a hemocytometer and the final spore inoculums were adjusted to give a concentration of 1x106 spores mL-1.
Pretreatment of Rice Straw and POME
Rice Straw (RS) was obtained from a rice field in Sekinchan, Selangor, Malaysia. It was sundried prior grinding it using hammer mill. It was pretreated using 0.5% NaOH at 121°C for 20 min with ratio of substrate to NaOH solution of 1:10. Rice straw was washed and rinsed using distilled water until the pH become stable at nearly neutral pH. It was dried at 60°C overnight and kept at room temperature prior to use. POME was obtained from Seri Ulu Langat Palm Oil Mill in Dengkil, Selangor, Malaysia. It was kept at 4°C until further used.
| Table 1: | List of individual and cocktail enzymes |
 | |
Fermentation Condition
Fermentation was done in 250 mL shake flasks with cotton stopper. For each flask, 2 g of dried rice straw was added and autoclaved prior to the addition of 60% (6 mL g-1 substrate) moisture which was modified from Mandel’s medium (Mandels et al., 1974). One liter of modified Mandel’s medium at pH 4.8 contained 1.4 g (NH4)2SO4, 2.0 g KH2PO4, 0.3 g CaCl2, 0.3 g MgSO4.7H2O, 1 mL of trace element and 50 mL of POME (no POME added for control media). Each flask was inoculated with 1 mL of 1x106 spore mL-1 of respective spore suspension and incubated at 30°C for 5 days.
Enzyme Collection and Preparation of Cocktail Enzyme
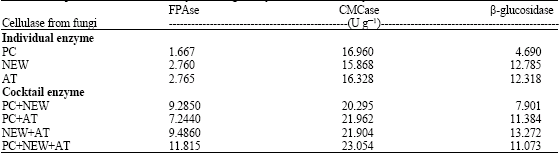
The enzyme produced by individual organism was extracted from the biomass by adding 40 mL of 50 μM acetate buffer into each flask (20 mL g-1 substrate) and immersed for 30 min. All enzymes were then centrifuged at 3000 rpm for 15 min. The supernatant were filtered through filter paper and kept at 4°C prior use, or kept at -20°C for storing purpose. Enzyme cocktail were prepared by mixing the individual enzyme in all possible combinations with same ratio (Table 1) prior subjected to cellulase assay.
Sample Analysis
Cellulose, hemicelluloses and lignin content in rice straw were determined by performing the standard methods (AOAC, 1997). POME characteristics were analyzed using standard method (APHA, 1998). Characteristics analyzed including Chemical Oxygen Demand (COD), biological oxygen demand, total solid, volatile solid, nitrogen content and etc. Cellulase assay of the extracted enzymes and cocktail enzymes were performed using Wood and Bhat method (Wood and Bhat, 1988) where FPAse, CMCase and β-glucosidase activities were determined individually. All experiments were done in duplicate and the results obtained were the mean of two data.
SEM Micrograph
Sample was observed under SEM for physical changes after pretreatment. Scanning Electron Microscope (SEM) viewing was conducted using SEM model JOEL CN6400. Sample was prepared by drying it and mounted on stub. It was then coated with gold prior viewing under SEM.
RESULTS
Effect of Pretreatment of Rice Straw
Total fibers of rice straw prior and after pretreatment are shown in Table 2. Pretreatment using 0.5% NaOH at 121°C for 20 min removed 14.11% of lignin from total fibers thus increased cellulose percentage in the fiber by 32.29%. SEM pictures (Fig. 1) show properties of treated and untreated rice straw. It can be observed that lignin and hemicelluloses were effectively removed from the rice straw while cellulose structure was maintained after NaOH pretreatment.
| Table 2: | Comparison of lignocellulosic content in raw and pretreated rice straw |
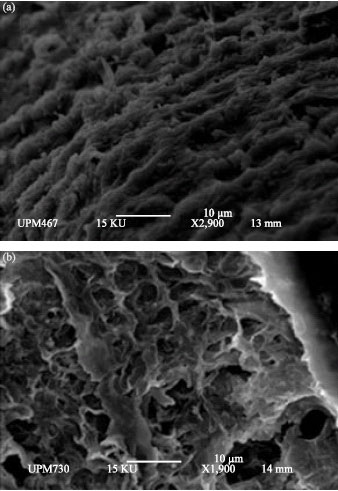 | |
| Fig. 1: | SEM pictures of rice straw (a) before pretreatment and (b) after pretreatment using 0.5% NaOH at 121°C for 20 min |
POME Characteristics
Properties of POME used in this study are shown in Table 3. Main constituent and its concentration are the total solid (TS, 40 000-50 000 mg L-1), total suspended solid (TSS, 18 980 mg L-1), oil and grease (1 000-2 000 mg L-1), total Kjeldahl Nitrogen (TKN, 400-500 mg L-1), volatile suspended solid (VSS, 15 000-20 000 mg L-1), total carbohydrate (92.50%), ammonium nitrogen (120.96 mg L-1), ash content (31 386 mg L-1) and lignin (1 345 mg L-1). The pH of the POME was recorded to be around 4.0 to 5.0.
| Table 3: | Major POME constituents from Seri Ulu Langat Palm Oil Mill, Dengkil Selangor used in this study |
 | |
| Table 4: | Comparison of cellulase activity in control media and POME-added media in rice straw using three different fungal species |
 | |
| Table 5: | Comparison of cellulase activity from single enzyme and cocktail enzyme added with cellulase |
 | |
Effect of POME Addition on Enzyme Activity
The results of cellulase performance for both individual and cocktail enzyme are shown in Table 4 and 5, respectively. It was observed that the overall enzyme activity increased in POME-Mandels medium except for FPAse of NEW fungi (Aspergillus sp.) which reduced by 21%. FPAse for PC and AT showed increment by 72 and 19%, respectively. In addition, FPAse activity for PC was almost same if 50 ml L-1 of POME was substitute with 3 g L-1 of peptone in another study (data not shown here). Meanwhile, CMCase and β-glucosidase activities for all fungi increased significantly. In fact, CMCase activity by PC was observed to be the highest increment (236%), while highest β-glucosidase activity increment observed was from AT (298%).
FPAse activity by cocktail of PC and AT increased by 227% over the average activity of both of the fungi. Meanwhile the highest FPAse activity increment was by cocktail of all the three species of fungi which shower 393% improvement. The same trend goes for CMCase except the activity increments was not as significant as FPAse, where the highest CMCase activity observed was cocktail by all the fungal species used in the study which increased by 41% followed by cocktail of NEW and AT which is 36%. However, in β-glucosidase activity, any cocktail with PC enzyme will not increase the activity. Only cocktail of NEW and AT showed increment from their average activity which was by 6%. In this study, the highest FPAse and CMCase activity were the cocktail of PC, NEW and AT, while for β-glucosidase, the highest activity was from cocktail of NEW and AT.
DISCUSSION
Effect of Pretreatment of Rice Straw
Muthuvelayudham and Viruthagiri (2006) reported that removal of lignin is crucial for increasing cellulase activity which can be observed whereby the cellulose provided is more digestible. From Table 2 and Fig. 1, it is observed that lignin reduction is related with the rice straw structure. Removal of lignin can significantly simplified the rice straw structure thus providing the fungi more digestible biomass for cellulase production.
POME Characteristics
From Table 3, main constituent to be highlighted are the total solid, total Kjeldahl nitrogen, total carbohydrate and ammonium nitrogen. The supplementation of POME into the fermentation media was due to its high concentration of carbohydrate and nitrogen. Wu et al. (2009) reported that there are a number of studies on producing cellulase directly from POME where simple carbohydrate and high nitrogen content renders the possibility to enhance cellulase production.
Effect of POME Addition on Enzyme Activity
Generally, by comparing Mandels medium and POME-Mandels medium, it is observed that fungi supplied with POME-Mandels medium gave higher cellulase activity. In comparison, Prasetsan et al. (1997) reported that CMCase activity increased by 43% with the addition of 0.6 g L-1 NH4NO3. Therefore, in this study, it was suggested that the main factor for the increase in cellulase activity might be influenced by relatively high concentration of carbohydrate and nitrogen in POME (Table 2). However, it is not concluded which parameter affect the increment and further analysis are required to determine it.
Synergistic effect of cellulase cocktail has been reported for a while (Henrissat et al., 1985). In the enzyme cocktail case, especially the FPAse, most of the enzyme combinations react synergistically, increasing the FPAse activity higher than doubling the average activity of the enzymes themselves. In this study, it is observed that cellulase cocktail improved FPAse and CMCase obviously. However in β-glucosidase case, any cocktail with cellulase from PC did not increase the activity. This is may be due to the β-glucosidase activity of PC which is already low compared to AT and NEW, thus leading to a lower result as compared to cocktail of NEW and AT. It seems that β-glucosidase activity plays a major role in determining the total cellulase activity. It was reported that β-glucosidase is the restricted component in multi-enzyme system (Zhou et al., 2009). Therefore, it is crucial to obtain high activity β-glucosidase for saccharification.
CONCLUSION
POME supplementation in fermentation media indirectly increases the cellulase activity. Nevertheless the supplementation of POME into media might not be applicable to all fungi. Individually, the FPAse activity increased in P. chrysosporium and A. terreus but decrease in Aspergillus sp. Additionally, CMCase and β-glucosidase activities were increased in all the three species of fungi used. Variation of combinations for these enzymes as enzyme cocktail further improves the whole enzyme performance. This was observed in FPAse and CMCase activity of all cocktail. However, in the case of β-glucosidase, it is advisable to produce a significantly high activity β-glucosidase separately since combination with lower activity β-glucosidase reduced to total β-glucosidase activity. Main component in POME which affects the enzyme activity also need to be further investigated.
ACKNOWLEDGMENTS
The authors would like to thank the Ministry of Science Technology and Innovation of Malaysia for financially support throughout this research project and Assoc. Prof. Dr. Zahangir Alam for providing the fungi strain.
REFERENCES
- Baharuddin, A.S., M. Wakisaka, Y. Shirai, S. Abd-Aziz, N.A. Abdul-Rahman and M.A. Hassan, 2009. Co-composting of empty fruit bunches and partially treated palm oil mill effluents in pilot scale. Int. J. Agric. Res., 4: 69-78.
CrossRefDirect Link - Balat, M. and H. Balat, 2009. Recent trends in global production and utilization of bio-ethanol fuel. Applied Energ., 86: 2273-2282.
CrossRefDirect Link - Busto, M.D., N. Ortega and M.P. Mateos, 1995. Induction of β-glucosidase in fungal and soil bacterial culture. Soil Bio. Biochem., 21: 949-954.
CrossRefDirect Link - Cardona, C.A. and O.J. Sanchez, 2007. Fuel ethanol production: Process design trends and integration opportunities. Bioresour. Technol., 98: 2415-2457.
CrossRefDirect Link - Fang, X., S. Yano, H. Inoue and S. Sawayama, 2009. Strain Improvement of Acremonium cellulolyticus for cellulase production by mutation. J. Biosci. Bioeng., 107: 256-261.
CrossRefDirect Link - He, B.Q., J.X. Wang, J.M. Hao, X.G. Yan and J.H. Xiao, 2003. A study on emission characteristics of an EFI engine with ethanol blended gasoline fuels. Atmos. Environ., 37: 949-957.
Direct Link - Henrissat, B., H. Driguez, C. Viet and M. Schulein, 1985. Synergism of cellulases from Trichoderma reesei in the degradation of cellulose. Bio. Technol., 3: 722-726.
CrossRefDirect Link - Inoue, H., S. Yano, T. Endo, T. Sakaki and S. Sawayama, 2008. Combining hot-compressed water and ball milling pretreatments to improve the efficiency of the enzymatic hydrolysis of Eucalyptus. Biotechnol. Biofuel., 1: 2-2.
CrossRefDirect Link - Mandels, M., L. Hontz and J. Nystrom, 1974. Enzymatic hydrolysis of waste cellulose. Biotechnol. Bioeng., 16: 1471-1493.
CrossRefDirect Link - Morikawa, Y., T. Ohashi, O. Mantani and H. Okada, 1995. Cellulase induction by lactose in Trichoderma reesei PC-3-7. Applied Microbiol. Biotechnol., 44: 106-111.
CrossRefDirect Link - Mosier, N., C. Wyman, B. Dale, R. Elander, Y.Y. Lee, M. Holtzapple and M. Ladisch, 2005. Features of promising technologies for pretreatment of lignocellulosic biomass. Bioresour. Technol., 96: 673-686.
CrossRefDirect Link - Muthuvelayudham, R. and T. Viruthagiri, 2006. Fermentative production and kinetics of cellulase protein on Trichoderma reesei using sugarcane bagasse and rice straw. Afr. J. Biotechnol., 5: 1873-1881.
Direct Link - Prasertsan, P., A.H. Kittikul, A. Kunghae, J. Maneesri and S. Oi, 1997. Optimization for xylanase and cellulase production from Aspergillus niger ATTC 6275 in palm oil mill wastes and its application. World J. Microbiol. Biotechnol., 13: 555-559.
CrossRef - Rho, D., M. Desrochers, L. Jurasek, H. Driguez and J. Defaye, 1982. Induction of cellulase in Schizophyllum commune: Thiocellobiose as a new inducer. J. Bacteriol., 149: 47-53.
Direct Link - Roca-Perez, L., C. Martinez, P. Marcilla and R. Boluda, 2009. Composting rice straw with sewage sludge and compost effects on the soil-plant system. Chemosphere, 75: 781-787.
CrossRefDirect Link - Sulaiman, A., Z. Busu, M. Tabatabaei, S. Yacob, S. Abd-Aziz, M.A. Hassan, Y. Shirai, 2009. The effect of higher sludge recycling rate on anaerobic treatment of palm oil mill effluent in a semi-commercial closed digester for renewable energy. Am. J. Biochem. Biotechnol., 5: 1-6.
CrossRefDirect Link - Wood, T.M. and K.M. Bhat, 1988. Methods for measuring cellulase activities. Methods Enzymol., 160: 87-112.
CrossRefDirect Link - Wu, T.Y., A.W. Mohammad, J.M. Jahim and N. Anuar, 2009. A holistic approach to managing palm oil mill effluent (POME): Biotechnological advances in the sustainable reuse of POME. Biotechnol. Adv., 27: 40-52.
Direct Link - Zhou, J., Y.H. Wang, J. Chu, L.Z. Luo, Y.P. Zhuang and S.L. Zhang, 2009. Optimization of cellulase mixture for efficient hydrolysis of steam-exploded corn stover by statistically designed experiments. Bioresour. Technol., 100: 819-825.
CrossRefDirect Link - Balat, M., H. Balat and C. Oz, 2008. Progress in bioethanol processing. Progress Energy Combustion Sci., 34: 551-573.
CrossRef