
P. Thobunluepop
Department of Agricultural Technology, Faculty of Technology, Mahasarakham University, Talard Sub-District, Muang District, Maha Sarakham 44000, Thailand
W. Chitbanchong
Plant Variety Protection Research Group, Plant Variety Protection Division, Department of Agriculture, Ministry of Agriculture and Cooperatives, Royal Thai Government, Bangkok, 10900, Thailand
E. Pawelzik
Department of Agronomy, Faculty of Agriculture, Chiang Mai University, 50000, Thailand
S. Vearasilp
Institute of Agricultural Chemistry, Section of Quality of Plant Products, Georg-August University of Goettingen, Carl-Sprengel-Weg, D-37075, Goettingen, Germany
International Journal of Agricultural Research
Year: 2009 | Volume: 4 | Issue: 5 | Page No.: 169-184







ABSTRACT
The effect of pre-sowing seed treatments in direct-seeding rice production system on the germination, seedling vigor of rice cv. KDML 105 was evaluated through investigating the biochemical changes during storage following seed coating techniques. The seeds were coated by traditional fungicide (captan; CA), biological fungicide polymers [chitosan-lignosulphonate polymer (CL) and eugenol incoporated into chitosan-lignosulphonate polymer (E+CL)] and un-coated seeds as control (CO). CA significantly affected the rice seed storability and the associated biochemical deterioration. After 12 months storage, seed moisture content and seed water activity increased that affected the germination rate and spread, seedling vigor; seedling dry weight, shoot and root length, seedling growth rate and susceptible to stress conditions. The loss of viability is associated with disturbances of the cell membranes, the loss of enzymes; α-amylase, ascorbate peroxidase APX and superoxide dismutase SOD activity, sugars and lipid content accompanied by increased free fatty acid FFA and activated lipoxygenase enzyme LOX. CL and E+CL performed the best of seed vigor, because they could maintain the antioxidative scavenging enzymes are APX and SOD and a high antioxidant activity. In addition, α-amylase activity and sugar content increased which was positive correlated with seed germination and vigor. These improvements were attributed to maintain the nutritive reserve and dehydrogenase activity in seeds. Moreover, the biological seed treatments stimulated the embryo growth and so speeding up the seedling emergence.
PDF Abstract XML References Citation
How to cite this article
P. Thobunluepop, W. Chitbanchong, E. Pawelzik and S. Vearasilp, 2009. Physiological and Biochemical Evaluation of Rice Seed Storability with Different Seed Coating Techniques. International Journal of Agricultural Research, 4: 169-184.
DOI: 10.3923/ijar.2009.169.184
URL: https://scialert.net/abstract/?doi=ijar.2009.169.184
DOI: 10.3923/ijar.2009.169.184
URL: https://scialert.net/abstract/?doi=ijar.2009.169.184
INTRODUCTION
Rice occupies a conspicuous position in the agro-based economy of Thailand. Besides meeting the dietary requirements of the people, it has emerged as a major export commodity contributing about 9.4 million-tons or 5.8 million US dollars in 2008, increasing for 11.3% from 2007. In Thailand, it was grown on an area 9.25 million h with the average yield of 3,431 kg ha-1 (Rice Department of Thailand, 2009).
Direct-seeding rice production system could be an attractive alternative of the traditional rice culture but poor results of seed germination, seedling establishment and high fungal infection are account the main constrains to the adoption of this production system (Du and Tuong, 2002). More than 90% of the rice grown in the world are propagated through seeds and all of them are attacked by devastating seed born pathogens. The rice crop (Oryza sativa) is known to be attacked by many pathogenic fungi, e.g., Bipolaris sp. (brown spot), Alternaria sp. (stackburn), Fusarium sp. (bakanae), Rhizoctonia sp. (sheath blight), Nigrospora sp. (kernel smut), Curvularia sp. (blast). Important storage fungi are Aspergillus flavus and Aspergillus niger. Fungi contamination is a major cause leading to seed deterioration and finally to the degradation of rice grain qualities (Hewett, 1987). Selective fungicides are now available for the effective of fungal control. Nowadays, fungicide seed treatments are the most used traditional application to protect the seeds and young-seedlings from many seed-and soil-borne pathogens. However, the uses and expectations of chemical seed treatment have raised the concern due to the impact on environment, which cause either direct or indirect impacts on human or other living organisms. It can also negatively affect the food chain, the ecosystem at all and may lead to fungicide resistance problems (Ester et al., 2003). Moreover, the fungal pathogen normally attached to the seed, so it is difficult to find chemical substances that will destroy the fungus without harming the seeds. Furthermore, the toxicity of fungicide has been ascribed to produce phytotoxic compounds induced seeds deterioration (Han, 2000). The use of medical plant extracts in plant pathogenic fungi control is a key application in chemical-free agricultural system. The use of biodegradable, low-toxicity, natural products for biological control of fungi is the today trend (Burt and Reinders, 2003). In order to minimize undesirable side effects, many academic interests have focused on natural bio-fungicides as alternatives for toxic synthetic fungicides. Several antifungal compounds of plant origin are known to control the seed borne infection (El-Ghaouth, 1997).
Phenolic compounds, as eugenol and eugenol acetate, are the main components of clove (Eugenia caryophillis) (Velluti et al., 2004). They were proved to have bactericide, virocide (Tullio et al., 2006) and fungicide effects and act against many seed borne fungi (Thobunleupop et al., 2008). The use of coated seeds with chemicals or alternative bioactive substances may reduce significantly the percentage of plant damage by pest and the level of pesticide application Moreover, this application have been used to reduce the seedling emergence time, produced synchronized emergence, improved emergence rate and increased seedling stand production in many field crops like wheat (Basra et al., 2005) and rice (Farooq et al., 2004). Seed quality has pronounced effects of progeny survival and fitness, especially under storage conditions and stressful conditions in general (Agrawal, 2002). The morphological and chemical attributes of these seed components are critical for successful seedling establishment under various storage or stress conditions. Moreover, seedlings development depends on seed reserves in the early stages of development. Thus, the present study aimed to investigate the effects of various seed coating substances on rice seed (cv. KDML 105) quality, seedling performances and chemical and biochemical changes during storage.
MATERIALS AND METHODS
The experiment was conducted at section of Seed Science and Technology, Department of Agronomy, Faculty of Agriculture, Chiang Mai University (CMU), Department of Agricultural Technology, Faculty of Technology, Maha Sarakham University (MSU), Thailand and Institute of Agricultural Chemistry, Georg-August University of Goettingen, Germany during the year 2007-2008. Dry graded rice seeds (Oryza sativa L. cv. KDML 105) from one seed lot were supplied by Bureau of Seed Multiplication of Thailand. The split-plot design with four replications was applied. The main plot was seed treatment; captan (CA), only chitosan lignosulphonate polymer (CL), eugenol incorporated into chitosan lignosulphonate polymer (E+CL) and control seeds (CO). The sub-plot was the storage duration (12 months). The seeds of each treatment were randomly stored in plastic bag sealed in an incubation chamber (KPB6395FL, Termaks, S/N 2-858 Germany), at controlled temperature of 30±2°C and relative humidity at 40±5%. Seeds were sampled immediately and then collected each month for the determination of seed qualities and the seedling performances.
Seed Treatments
The following seed treatments were employed for seed coating application:
Traditional Captan Treatment Preparation (CA)
Captan fungicide (1, 2, 3 and 6-tetrahydro-N-(trichloromethyl thio) phthalimide was applied as slurry dust in a solution of polyethylene glycol (PEG 8000 at-2 MPa) (4 g of captan per 1 kg of the seeds). After that, seeds were dried to 10±2% of Moisture Content (MC) at 35°C to obtain a similar MC as the control (untreated) seeds.
Chitosan-lignosulphonate Coating Polymer Preparation (CL)
Three percentage of chitosan-lignosulphonate coating polymer was prepared by adding 3 g chitosan (Fluka, Germany) into 100 mL of 1% v/v acetic acid combined with 1% w/v sodium lignosulphonate (Fluka, Germany) in distilled water. Then, the chitosan-lignosulphonate polymer was sprayed and mixed well into 500 g seeds. The seed MC of 10±2% was obtained after drieing at 35°C.
Eugenol Incorporated into Chitosan-lignosulphonate Coating Polymer Preparation (E+CL)
One percentage of eugenol incorporated into chitosan-lignosulphonate coating polymer was prepared by adding 0.5 mL eugenol solution (Fluka, Germany) into 50 mL of 3% chitosan-lignosulphonate coating polymer binder. Then, it was sprayed onto mixed-well 500 g seeds samples. The seeds were dried at 35°C to obtain MC of 10±2%.
Seed Physical Properties Assessment
The seeds Moisture Content (MC) was determined by hot air oven method and calculated on a wet mass basis. Testo 650 apparatus was used for analyzing the seed water activity (Aw) and seed viability was measured as percentage of normal seedlings by standard germination test (Germ) in rolled-towel paper at 28°C for 14 days, which was carried out according to ISTA (2006). Dehydrogenase activity of seeds was analyzed by tetrazolium test (TZ test) according to ISTA (2006) and Germination Index (GI) as high speed of germination is an indication of vigorous seed lots. The number of germinating seeds was counted every day from the first day (14 days). The cumulative index was calculated by the following formula:
GI = (n1/1 + n2/2 + n3/3 + … + nx/x) |
where, n1…nx are the number of germinated seed on day 1 to day x
The time to reach 50% germination (T50) of final germination was calculated according to the following formulae, of some modified by Farooq et al. (2005);
where, N is the final number of germination and ni, nj is the cumulative number of seeds germinated by adjacent counts at time ti and tj when ni<N/2<nj. Seedling vigor data were recorded according to ISTA (2006) as following; Seedlings Dry Weight (SDW) determining by the dry weights (milligram) of seedlings was determined on the 14th day after germination. After that, seedlings were oven dried at 80°C for 24 h.
Seedling Growth Rate (SGR) was calculated from the equation as:
SGR = SDW – (SHW + RTW + RSW) |
where, SDW is dry weight of seed before germination, SHW is dry weight of shoot, RTW is dry weight of root, RSW is dry weight of seed after germination.
The electrical conductivity test (Cond), 100 seeds were weighted and soaked into de-ionized water. Then, they were placed at 25°C for 24 h. Finally, the solution was determined for electrical conductivity by the conductivity meter (Sartorius, model PP-20).
The percentage of field emergence (Field) and uniformity of seedling under field condition were determined by Brix test.
Seed Health Testing
According to ISTA (2006), the Blotter method is the testing method recommended for seed borne fungi detection. The procedure started with the test of a 400 seeds working sample in four replications and 25 seeds per dish were placed on three filter papers (blotters) which were soaked well in sterilized water. The seeds were later incubated at 20-25°C in 12 h light for 14 days. Seed borne fungi infection was recorded under a stereoscopic microscope (Olympia-SZ61). Then, the inhibition percentage of each seed born species was calculated, based on control seed, with the following equation (ISTA, 2006):
Seed Biochemical Properties Assessment
All experiments were done using dehulled grains with 12±0.5% MC. Freeze-dried rice seeds were ground (UDY, Cyclone Sample Mill No.2).
Determination of Total Protein Content
The protein content was analyzed according to Kjeldahl method. First, 4 mL of sulfuric acid and the catalyst (K2SO4: CuSO4.5H2O: Se; 10:1:0.1) was added to 0.25 g of ground rice seed. The sample was digested at 370°C. After complete the digestion, the solution was distilled using Kjeldahl Protein Analyzer, into a receiving flask containing boric acid indicator. The distilled sample was titrated by 0.1 N sulphuric acid. The protein content of the sample was obtained from the multiplying the nitrogen determined by 6.25.
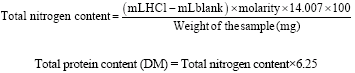 |
Determination of Total Lipid and Free Fatty Acid (FFA) Content
According to Lam and Proctor (2000), the gravimetric method was used to determine the total lipid content. Lipid content was extracted by vortexing 10 g of rice sample with 4 mL of isopropanol for 5 min. Then, added 5 mL of isopropanol into the sample and vortexed for 5 min. After that the extract was centrifuged at 2500 rpm for 10 min. The weight of extracted lipids was determined after evaporating the solvent on an electric hot plate at 40°C. According to Walde and Nastruzzi (1991), FFA of the sample was determined by preparing an assay solution, which contained 0.375 mL of solution A (0.1 M tris/HCL; pH 9.0), 0.125 mL of solution B (2 mM phenol red in 0.1 M Tris/HCl; pH 9.0) and 0.5 mL of solution C (50 mM Bis (2-ethylhexyl) sodium sulfosuccinate in isooctane). Then, 30 μL of isopropanol extract was mixed with 1 mL of assay solution in a 1 cm wide cuvette and was shaked for one minute before measuring absorbance at 560 nm. FFA of each extract was obtained from a calibration curve. The calibration curve was prepared by dissolving oleic acid in isopropanol to produce oleic acid solution of 0.001 to 0.02% (w/w).
Determination of Sugar Content
Sugar content was determined by iodine titration of excess copper of Luff-Schoorl method (Alexander et al., 1985). Sample solution preparation: 5 g of ground sample was accurately weighed to nearest 1 mg and transferred to a 250 mL volumetric flask, which contained 200 mL distilled water. Five milliliters of carrez solution I, (21.795 g of zinc acetate dehydrate (Zn (CH3COO) 2.2H2O) and 3 mL of glacial acetic acid were dissolved and made up to 100 mL with distilled water) and 5 mL of Carrez solution II, (10.76 g of potassium hexacyanoferrate II trihydrate (K4 [Fe (CN) 6].3H2O) in water and make up to 100 mL with water), were mixed after each addition and made up to 250 mL with water and mix well. Luff-Schoorl reagent preparation: citric acid solution (50 g of citric acid dihydrate (C6H8O7.2H2O) in 50 mL of water) was added to sodium carbonate solution (143.78 g of anhydrous sodium carbonate in 300 mL of warm water and allow to cool) in a one-liter volumetric flask with gentle swirling. Then, copper II sulphate pentahydrate solution (25 g of CuSO4.5H2O in 100 mL of water) was added and made up to 1000 mL with water. The solution was allowed to stand overnight and then filtered. Twenty-five milliliters of Luff-Schoorl reagent and 25 mL of sample solution were transferred into 250 conical flasks and extracted at 80°C in ultrasonic bath (Bandelin, Germany) for 10 min. After that, the sample was immediately cooled for 5 min. Then, 10 mL of 30% (w/v) potassium iodide solution were added and immediately followed by 25 mL of 3 M sulphuric acid adding. Titration was done with 0.71 M sodium thiosulphate solution until the solution was almost colorless, then added a few milliliters of starch indicator (5 g of soluble starch; Sigma, St. Louis, MO, USA.) slurred in 30 mL of water, boil for 3 min, allowed the mixture to cool and added water to make the 1 L solution). The titration was continued until the blue color disappears.
Finally, sugar content was calculated from the standard table by using the weight of glucose or the weight of invert sugar in percentage corresponding to the difference between the two-titration readings, expressed in milliliter of 0.71 M sodium thiosulphate. Express the results in terms of invert sugar or D-glucose as percentage of the dry matter.
Determination of Total Antioxidant Activity
Total antioxidant activity was determined by the diphenylpicrylhydrazyl (DPPH) radical scavenging effect according to Kim et al. (2002). To obtain the concentration of crude methanolic extract, one gram of ground rice seed sample was dissolved in 50 mL methanol (Merk, Germany) and the ultrasonic solvent extraction was carried out in an ultrasonic bath (Bandelin, Germany) for 1 h. The sample solution was then filtered. The filtrate was evaporated with rotary evaporator (Buchi, Switzerland) at 40°C until dried. Crude extract was then weighed and calculated back to obtain the concentration in mg (crude) per mL (methanol). To determine the antioxidant activity, three milliliters of methanol solution of each treatment at various concentrations (1.5-45 mg mL-1) were added to 1 mL of 1x10-4 M DPPH (Fluka, Germany) in methanol. The reaction mixture was shaken vigorously. After leaving the mixture at the room temperature for 30 min, the optical density was measured at 520 nm using a UV-VIS spectrophotometer (SPECCORD 40). The radical scavenging activity of each sample was expressed by the ratio of lowering of the absorption of DPPH (%) and the absorption (ASB) (100%) of DPPH solution in the absence of test sample (control). The mean value was obtained from four replication experiments. The percent inhibition was calculated by the following equation:
where, ASB is absorption values
The antioxidant activity of each sample was expressed in terms of fifty percent of effective concentration (IC50) value, which required inhibition of DPPH radical formation by 50%. The IC50 value was calculated by plotting the inhibition percentage of each tested concentrations in the logarithmic graph. A logarithmic equation was used to calculate IC50 value.
Determination of Ascorbate Peroxidase Activity (APX)
Enzyme extraction: 5.0 g of rice powder was weighed into a centrifuge tube. Five milliliters of extraction buffer (0.1 M phosphate buffer, pH 6.0) were added and mix well on shaker for 15 min. The mixture was then centrifuged at 12 000 rpm at 4°C for 30 min. Supernatants were used for enzyme assays. All steps of extraction procedure were carried out at 1-4°C. Soluble protein content of the extracts was determined by the method of Bradford (1976), which used a Bio-Rad protein assay kit with bovine serum albumin (Sigma, St. Louis, MO, USA.) as the calibration standard (150-900 μg mL-1) at 595 nm. Fifty microliters of extracted solution added to 2.5 mL Bio-rad before it was vortexed for 5 min. Then the mixture was shaken for one minute before measuring the absorbance at 595 nm. APX was determined according to Nakano and Asada (1981), with some modifications. The reaction mixture (2.0 mL) contained 0.05 M phosphate buffer (pH 7), 0.1 mM EDTA, 5 mM ascorbate and 800 μL of crude enzyme extract. The reaction was initiated by adding 200 μL H2O2 (2 mM). The decreasing of absorbance from the oxidation of ascorbate at 290 nm was recorded using a spectrophotometer (Hewlett Packard 8453, Germany) at 1,200 sec after the adding H2O2. The activity of APX was expressed as μmole min-1 100 mg-1 protein.
Determination of Superoxide Dismutase Activity (SOD)
The method of Oberley and Spitz (1985), with some modifications was used to determine the activity of SOD. One milliliter of reaction mixture for the determination of SOD activity contained 800 μL of reaction mixture (0.1 mM Xanthine, 0.056 mM nitro blue tetrazolium (NBT), 1.0 mM DETAPAC and 1 U CAT in 0.05 M phosphate buffer (pH 7.8)), 100 μL of enzyme extracted and 100 μL of 0.1 U mL-1 xanthine oxidase. SOD was evaluated by measuring the ability of the enzyme extract to inhibit the photochemical reduction of NBT. The reaction was initiated by illuminating the reaction mixtures at 26-28°C for 30 min and absorbance was read at 590 nm. One unit of SOD was defined as the enzyme activity that inhibited photoreaction of NBT to blue formation by 50%. SOD activity of the extracts was expressed as Δ activity mg-1 protein.
Determination of Lipoxygenase Activity (LOX)
Enzyme extraction: 3.0 g of rice powder from the seeds subjected to various duration of ageing were homogenized with 5 mL of extraction buffer (0.05 M sodium borate buffer, pH = 9.0) and shaken for 30 sec. The mixture was centrifuge at 12000 rpm at 26-28°C for 5 min. For LOX determination, 5 mL of 96% (v/v) Ethanol were added and incubated at 26-28°C for 15 sec, then filtered. Afterwards, 3 mL of 70% (v/v) Ethanol were added to 50 μL of extract solution and absorbance was read at 234 nm using 70% (v/v) Ethanol for blank. LOX activity of the extracts was expressed as Δ activity mg-1 protein (Meshehdani, 1990).
Determination of α-Amylase Activity
One gram of ground rice seeds was mixed with 10 mL of phosphate buffer (pH = 7) and left for 24 h at 4°C. Enzyme activity of the supernatant was measured by Di Nitro Salicylic acid (DNS) method slightly modified by Basra et al. (2005).
Statistical Analysis
The data are presented as Mean±SD. The analysis of variance was performed for data analysis and differentiated with last significant different (LSD) test at p<0.05 using the software SX release 8.0 (Analytical software, Tallahassee, USA).
RESULTS
Seed Physical Properties
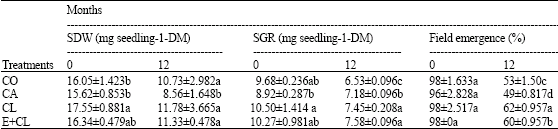
MC at the beginning of storage (0 month) showed the same levels, which was ranged from 11.18-11.50 g 100 g-1 DM in each treatment. However, MC increased significantly after 12 months storage, especially in CA treatment. On the other hand, CL and E+CL was no significantly different from CO (Fig. 1). At the beginning of storage, Aw ranged from 0.628-0.642 in each treatments. During the storage period, Aw was significantly affected by the seed treatment variants. After 12-months storage, CA seeds had significantly increased Aw. However, CL and E+CL treatments were non- significantly different when compared with CO (Fig. 2). Seed coated with various seed coating substances affected significantly (p<0.05) the seed viability and germination percentage of rice seeds. Statistically similarly earlier germination was recorded in seeds subjected to CL and E+CL as well as CO Moreover, they could maintain high seed storability, which was provided high percentage of seed viability after stored for 12 months. However, CA drastically decreased not only both characteristics (Fig. 3, 4) but also affected seed vigor. Uniform and earlier germinated was noted in CL, E+CL as well as CO as is evident from GI and T50 values. However, delayed and scattered germination was recorded from the seeds was coated with CA treatment (Table 1). Additionally, CA disrupted cell membrane stability, its showed highest seed conductivity. Conversely, CL and E+CL coating polymer did not affected on this property, which was not differed seed conductivity when compared with CO (Table 1). High Seedling vigor, as is evident from SDW and SGR values, was observed in CL and E+CL, whereas CA gave lowest of SDW and SGR than rest of both treatments and CO (Table 2).
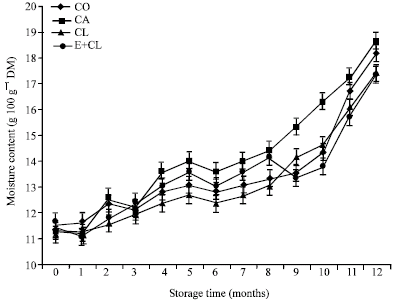 | |
| Fig. 1: | The effect of seed coating substances and storage time on seeds moisture content |
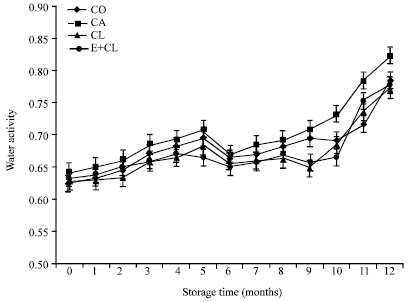 | |
| Fig. 2: | The effect of seed coating substances and storage time on seeds water activity |
| Table 1: | Effect of seed coating substances on seedling vigor are Germination Index (GI), the time to reach 50% germination (T50) and seed conductivity (Cond.) |
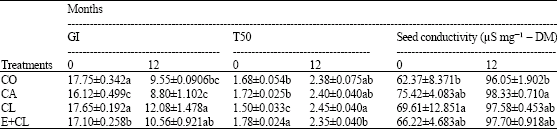 | |
| Data are expressed as Mean±SD. The different letter(s) indicate the statistically significant difference by LSD at 5% level | |
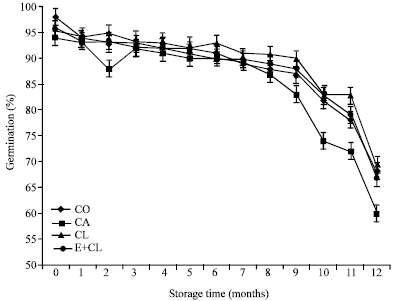 | |
| Fig. 3: | The effect of seed coating substances and storage time on seeds germination |
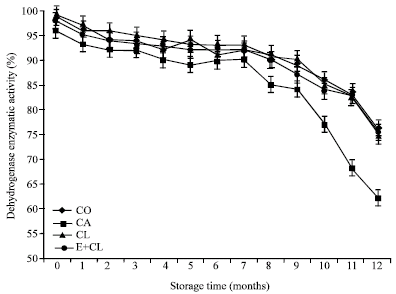 | |
| Fig. 4: | The effect of seed coating substances and storage time on seeds viability |
All the treatments resulted in high level of field emergence at the prior period of storage. However, after store for 12 months, CA reduced significantly field emergence, while CL, E+CL as well as CO could prolonged field emergence property of the seeds. The highest effective of seed infection control was observed in E+CL, which was similarly efficiency with CA treatments, followed by CL coated seeds. On the other hand, CO showed the highest of infected seeds (Fig. 5).
Seed Biochemical Properties
CL and E+CL as well as CO did not affect on total protein content except CA treatment. The changes of total lipid content not found in all the seed coating substances, while CA was provided the trend to reduce the content of total lipid, accompanied by significantly increased FFA. The lowest of total antioxidant activity was found in CA, while CL, E+ CL as well as CO could prolong the high levels (Table 3). The results indicated that seed treatment variants, storage duration influenced glucose, sucrose and lactose content in the seeds.
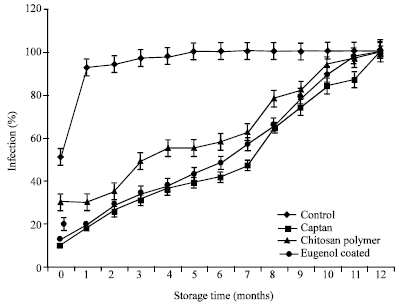 | |
| Fig. 5: | The effect of seed coating substances and storage time on seed infection |
| Table 2: | Effect of seed coating substances on seedling vigor are Seedling Dry Weight (SDW), Seedling Growth Rate (SGR) and field emergence (field) |
 | |
| Data are expressed as Mean±SD. The different letter(s) indicate the statistically significant difference by LSD at 5% level | |
| Table 3: | The effect of seed treatment variants on total protein, lipid, free fatty acid and antioxidant activity content changes during storage for 12 months |
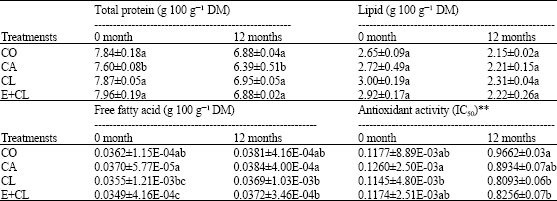 | |
| Data are expressed as Mean±SD. The different letter(s) indicate the statistically significant difference by LSD at 5% level. **IC50 was expressed in terms of the fifty percent of effective concentration (IC50) value, which required inhibiting DPPH radical formation by 50%. This IC50 parameter has the drawback value with the total antioxidant activity | |
However, the seed treatment variants showed no effect on the seed maltose content, but the storage duration was the main impact factors of maltose content. Table 4 at the beginning of storage (0 month), showed glucose, sucrose and lactose content significantly decreased in CA. When compare with the CL and E+CL, it provided significantly different results. After 12-months storage, the content of glucose, sucrose, lactose and total sugar content in CA notable lower than in CO, CL and E+CL, while maltose content did not changed. The highest activity of enzymes; APX, SOD as well as α-amylase, were noted in seed coated with CL and E+CL, followed by CO seeds. Increased activity of LOX were observed in CA treated seeds in both prior of storage and after stored for 12 months. Minimum activity of LOX was remarked in E+CL, CL as well as CO (Table 5). The correlation regression analysis resulted that all biochemical factors had a high correlation value with the seed physical properties. Seed treatment variants increased moisture content and water activity of the seed. It is negatively correlated with antioxidant mechanisms in rice seed, which affected SOD, APX and total antioxidant activity. However, the seeds treatment variants were found to have positive correlations with the LOX activity and free fatty acid content that noted lipid peroxidation. Additionally, seed treatment variants induced hydrolysis of starch, which affected α-amylase and sugar content and associated with the increasing of seed infection (Table 6).
| Table 4: | The effect of seed treatment variants on glucose, sucrose, lactose, maltose and total sugar content changes during storage for 12 months |
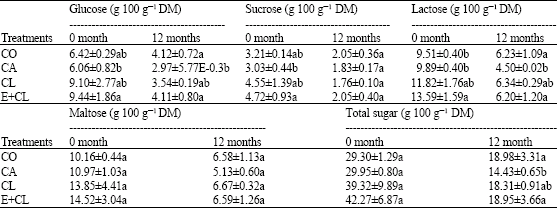 | |
| Data are expressed as Mean±SD. The different letter(s) indicate the statistically significant difference by LSD at 5% level | |
| Table 5: | The effect of seed treatment variants on ascorbate peroxidase (APX), superoxide dismutase (SOD) and lipoxygenase (LOX) and α-amylase enzymes activity changes during storage for 12 months |
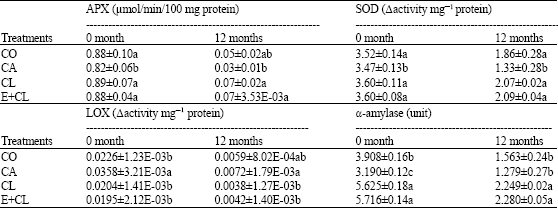 | |
| Data are expressed as Mean±SD. The different letter(s) indicate the statistically significant difference by LSD at 5% level | |
| Table 6: | Pearson correlation coefficients of seed coating substances on physical and biochemical properties of rice seeds** |
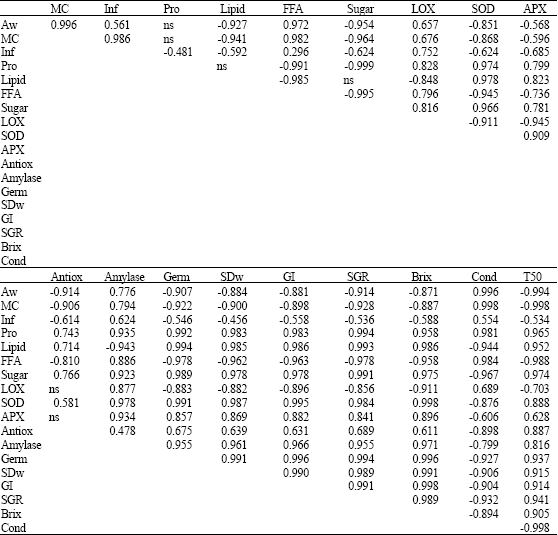 | |
| Number showed significant level at α = 0.05; ns = Not significant different at α = 0.05 level. **Data are involved in all treatments. Aw: Water activity, MC: Seed moisture content, Inf: seed infection, Pro: Protein, Lipid: Lipid, FFA: Total free fatty acid, Sugar: Total sugar, LOX: Lipoxygenase activity, SOD: Superoxide dismutase activity, APX: Ascorbate peroxidase activity, Germ: Seed germination, SDw: Seedling dry weight, GI: Germination index, SGR: Seedling growth rate, Brix: Field emergence, Cond: Conductivity, T50: The time to reach 50% germination, MET: Mean emergence time | |
DISCUSSION
Earlier and uniform germination and seedling vigor was observed in both of CL and E+CL coating substances as indicated by MET, T50, GI, SDw, SGR and Brix test. Lesser GI and T50 indicated the earlier and rapid germination. This findings supported the previous works on canola (Brassica compestris), wheat (Triticum aestivum) (Nayyar et al., 1995) and rice (Oryza sativa) (Lee and Kim, 2000) who reported the improvement of germination percentage and seedling vigor in seeds subjected to seed coating and priming techniques. This study revealed that employing two different seed coating substances; CL and E+CL coating substances, could invigorate rice seeds. The higher, earlier and better-synchronized germination was associated with increased of metabolic activity. These finding strongly supported the idea that enzymatic and metabolic activities are the basis of seed invigoration by seed coating technique, which is indicated by positively correlation between, sugar content, α-amylase activity and seed vigor index (Table 6). The faster emergence rate may be explained by the improvement of seedling establishment in coated seeds could be related to a rapid and regulated production of emergence metabolites. Additionally, the biological seed coating application enhanced the activity of α-amylase or other essential enzymes and increased the levels of soluble sugars in rice seed. Thus, germination metabolites could be provided in required amounts for improving the germination process. However, in the present study, the CA seed treatment caused poor performance of seedling including delaying and uncertainly emerging of the seedling. Farooq et al , (2005) reported that the poor seedling performances are results of the impaired ability of the seeds in utilizing the germination metabolites. Faster and uniform emergence rate after coating with CL and E+CL substances may be explained by an increased rate of provided in required amounts of germination metabolites, which promoted an increased rate of cell division in the meristem cells of shoot and root tips as previously found for wheat (Basra et al., 2002). The enhanced activity of α-amylase during seed coating process may be attributed to hydration during treatment, resulting in increased starch hydrolysis, increased content of total reducing sugars and lower contents of non-reducing sugars. The beneficially of increased starch hydrolysis following hydration treatments was not lost during the redrying process, as shown by the better rate and spread of germination. These findings are also in agreement with Ruan et al. (2002), who reported the increased of á-amylase activity, total sugar content was associated with higher germination of rice seeds.
Seeds were treated with CA resulted lowest of seed germination percentage, rate and decreased seedling vigor as SDW, GI and T50. The lower of seedling dry weight might be a result of more succulent growth. The shorter of seedling and required more germination rate might be due to the toxicity of captan fungicide. In congruence with Kozlowski (1986), seeds injury was reported after the use of captan. Van-Iersel and Bugbee (1996), described that the metabolism of captan is activated as aqueous slurry; it released tetrahydrophthalimide, tetrahydrophthalic acid and three molecules of chloride (Cl¯). The penetration of these compounds into the cell could cause phytotoxic effects induced seed deterioration (Yupsanis et al., 1994). The toxicity results in injury of the cellular organelles and membranes. This finding also provided CA treatment showed the highest of seed conductivity, which is explained seed membrane integrity. Most of the applied fungicides increased conductivity of seeds, which led of membrane degradation (Sung and Chiu, 1995).
CA treated seeds had higher seed moisture content and water activity, which is promoted lipid hydrolysis and oxidation. Those mechanisms provided the highest amount of FFA and a higher LOX activity than biological coated seeds and seeds without treatment (CO) (Table 5). The experiments suggested that under the stress conditions and during storage, lipid in rice seeds are broken down by LOX into FFA. According to Narvaey-Vasquez et al. (1999), the deterioration of rice seeds is generally accompanied with lipid peroxidation. An interesting result of the present study was that the CA treatment induced lipid peroxidation, which was seen in an inverse relationship between lipid and FFA content, lipid and LOX and a positive correlation between LOX and FFA as well (Table 6). This relationship was also found by Rosahl (1996), who reported that captan induced LOX activity. This mechanism releasing Reactive Oxygen Species (ROS) from membrane phospholipids in aleurone layer of rice seeds in response to chemical stress conditions could induced cell senescence (McCord, 2000). The ROS have the potential to damage cell membranes and is likely to be a primary cause of deterioration of seeds (Sung and Chiu, 1995). Moreover, several comprehensive reviews have identified ROS mediated lipid peroxidation, enzyme inactivation, protein degradation, disruption of cellular membranes and damage to genetic (nucleic acids) integrity as major cause of seed deterioration (McDonald, 1999). The oxidative scavenging enzymes (APX and SOD) as well as total antioxidant capacity were in biological coated seeds more active than in the CA treatment and CO (Table 6). The experiments suggested that those are important factors for scavenging ROS, which induced the seeds tolerance to oxidative stress conditions. These findings are similar to that of Esfandiari et al. (2007). Additionally, the loss of seed viability was associated with the decrease of SOD, APX and antioxidant activity in the seeds as reported by Zhao et al. (2006). Asada (2000) found that higher activities of SOD and APX decreased the level of ROS in the cells and increased the stability of cell membrane, as well as activate the Calvin cycle that could maintain the seed viability as well.
In CO, the seeds had decreased germination percentage and seedling vigor. The decrease of seed qualities might be a consequence of the damage of embryos and meristematic cells by fungi. Moreover, fungi also influenced the depletion of food reserves in the endosperm. The results showed that seed infection had an inverse relationship with biochemical degradation and seed properties (Table 6). According to Chandrashekar and Sandhyarani (1996), fungi utilized sucrose as a nutrient or carbon source, which caused a decrease in seed vigor by inhibiting the nutrient availability in the seed. It appears that a decrease in viability and germination of the seed because of fungi development led to the inhibition of amino acid and protein synthesis and the decrease in sugar and starch contents. The earlier study, Thobunluepop (2008) showed that E+CL acts as antifungal agent that could protected the seeds from fungal infection during storage. This finding was in agreement with Khin et al. (2006). Chitosan polymer has a very effective combination of attractive characteristics, i.e., stimulates plant growth, provides plant protection and can be mixed well with fungicide agents. Therefore, chitosan could be a very useful biocontrol polymer with a good perspective for plant production systems (Bautista-Banos et al., 2003).
CONCLUSION
In conclusion, the results of the present study suggested that seed coated with CL as well as E+CL are very effective tool for seed invigoration for rice seeds. However, the application of CA treatment affected the storability of rice seeds. The possible basis of seed invigoration by these seed coating applications are directly correlated with the germination, seedling vigor, the content of germination metabolites and enzymatic activities.
ACKNOWLEDGMENTS
Authors acknowledge the Nation Research Council of Thailand (NRCT), Royal Thai Government, for research financial supported of the present studies, gratefully thanks to Dr. Suchada Vearasilp, Prof. Dr. Elke Pawelzik, Prof. Dr. Udo ter Meullen, Prof. Dr. Wolfgang Lüke for they are valuable suggestions through out the research project.
REFERENCES
- Agrawal, A.A., 2002. Herbivory and maternal effects: Mechanisms and consequences of transgenerational induced plant resistance. Ecology, 83: 3408-3415.
Direct Link - Asada, K., 2000. The water-water cycle as alternative photon and electron sinks. Phil. Trans. R. Soc. Lond. B, 355: 1419-1431.
CrossRefDirect Link - Zhao, F., S. Guo, H. Zhang and Y. Zhao, 2006. Expression of yeast SOD2 in transgenic rice results in increased salt tolerance. Plant Sci., 170: 216-224.
CrossRefDirect Link - Basra, S.M.A., M.N. Zia, T. Mehmood, I. Afzal and A. Khaliq, 2002. Comparison of different invigoration techniques in wheat (Tiriticum aestivum L.) seeds. Pak. J. Arid Agric., 5: 11-16.
Direct Link - Basra, S.M.A., M. Farooq, R. Tabassam and N. Ahmad, 2005. Physiological and biochemical aspects of pre-sowing seed treatments in fine rice (Oryza sativa L.). Seed Sci. Technol., 33: 623-628.
CrossRefDirect Link - Bautista-Banos, S., M. Hernandez-Lopez, E. Bosquez-Molina and C.L. Wilson, 2003. Effects of chitosan and plant extracts on growth of Colletotrichum gloeosporioides, anthracnose levels and quality of papaya fruit. Crop Protect., 22: 1087-1092.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Yupsanis, T., M. Moustakas, P. Eleftherrious and K. Damalanidou, 1994. Protein phosporylation-dephosphorylation in alfalfa seeds germinating under salt stress. J. Plant Physiol., 143: 234-240.
Direct Link - Du, L.V. and T.P. Tuong, 2002. Enhancing the Performance of Dry-seeded Rice: Effects of Seed Priming, Seedling Rate and Time of Seedling. In: Direct Seeding: Research Strategies and Opportunities, Pandey, S., M. Mortimer, L. Wade, T.P. Toung, K. Lopes and B. Hardy (Eds.). International Research Institute, Manila, Philippines, pp: 241-256.
- El-Ghaouth, A., 1997. Biologically based alternatives to synthetic fungicides for the control of post harvest diseases. J. Ind. Microbiol. Biotechnol., 19: 160-162.
CrossRefDirect Link - Esfandiari, E.O., M.R. Shakiba, S.A. Mahboob, H. Alyari and M. Toorchi, 2007. Water stress, antioxidant enzyme activity and lipid peroxidation in wheat seedling. Int. J. Food Agric. Environ., 5: 149-153.
Direct Link - Ester, A., H. De-Putter, J.G.P.M. Van-Bilsen, 2003. Film coating the seed of cabbage (Brassica oleracea L. convar. Capitata, L.) and cauliflower (Brassica oleracea L. var. Botryti L.) with imidacloprid and spinosad to control insect pests. Crop Prot., 22: 761-768.
CrossRefDirect Link - Farooq, M., S.M.A. Basra, K. Hafeez and E.A. Warriach, 2004. The influence of high and low temperature treatments on the seed germination and seedling vigor of coarse and fine seed. Int. Rice Res. Notes, 29: 75-77.
Direct Link - Khin, L.N., N. Nitar, S. Chandrkrachang and F.S. Willem, 2006. Chitosan as a growth stimulator in orchid tissue culture. Plant Sci., 170: 1185-1190.
CrossRefDirect Link - Kim, J.K., J.H. Noh, S. Lee, J.S. Choi and H. Suh et al., 2002. The first total synthesis of 2, 3, 6-Tribromo-4, 5- dihydroxybenzyl methyl ether (TDB) and its antioxidant activity. Bull. Korean Chem. Soc., 23: 661-662.
Direct Link - Kozlowski, T.T., 1986. Effects on seedling development of direct contact of Pinus resinosa seeds or young seedlings with Captan. For. Pathol., 16: 87-90.
CrossRefDirect Link - Lam, H.S. and A. Proctor, 2001. Rapid methods for milled rice total lipid and free fatty acid determination. AACC Int., 78: 494-498.
CrossRefDirect Link - Van-Iersel, M.W. and B. Bugbee, 1996. Phytotoxic effects of fungicides on bedding plants. J. Am. Soc. Hort. Sci., 121: 1095-1102.
Direct Link - Walde, P. and C. Nastruzzi, 1991. Application of a new, simple and economic colorimetric method for the determination of non-esterified fatty acids in vegetable oils. Food Chem., 39: 249-256.
Direct Link - McCord, J.M., 2000. The evolution of free radicals and oxidative stress. Am. J. Med., 108: 652-659.
CrossRefDirect Link - McDonald, M.B., 1999. Seed deterioration: Physiology, repair and assessment. Seed Sci. Technol., 27: 177-237.
Direct Link - Meshehdani, T., J. Pokorny, J. Davidek and J. Panek, 1990. The lipoxygenase activity of rapeseed. Nahrung/Food, 34: 727-734.
CrossRefDirect Link - Nakano, Y. and K. Asada, 1981. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in Spinach chloroplasts. Plant Cell Physiol., 22: 867-880.
CrossRefDirect Link - Narvaey-Vasquez, J., J. Florin-Christensen and C.A. Ryan, 1999. Positional specificity of a phospholipase an activity induced by wounding systemic and oligosaccharide elicitors in tomato leaves. Plant Cell, 11: 2249-2260.
Direct Link - Velluti, A., V. Sanchis, J.A. Ramos and S. Marin, 2004. Effect of essential oils of cinnamon, clove, lemon grass, oregano and palmaross on growth of and fumonisin B1 production by Fusarium verticillioides in maize. J. Sci. Food Agric., 84: 1141-1146.
Direct Link - Tullio V., A. Nostro, N. Mandras, P. Dugo and G. Banche et al., 2006. Antifungal activity of essential oils against filamentous fungi determined by broth microdilution and vapor contact methods. J. Applied Microbiol., 102: 1544-1550.
CrossRefDirect Link - Rosahl, S., 1996. Lipoxygenase in plants-their in development and stress response. Z. Naturforsch [C], 51: 123-138.
PubMedDirect Link - Ruan, S., Q. Xue and K. Tylkowska, 2002. The influence of priming on germination of rice (Oryza sativa L.) seeds and seedling emergence and performance in flooded soil. Seed Sci. Technol., 30: 61-67.
Direct Link - Sung, J.M. and C.C. Chiu, 1995. Lipid peroxidation and peroxide-scavenging enzymes of naturally aged soybean seed. Plant Sci., 110: 45-52.
Direct Link - Thobunluepop, P., E. Pawelzik and S. Vearasilp, 2008. Possibility of biological seed coating application on direct-seed rice production: Emphasis on plant productivity and environment awareness. Agric. Sci. J., 39: 449-452.
Direct Link - Farooq, M., S.M.A. Basra, N. Ahmad and K. Hafeez, 2005. Thermal hardening: A new seed vigor enhancement tool in rice. J. Integr. Plant Biol., 47: 187-193.
CrossRefDirect Link - Burt, A.S. and R.D. Reinders, 2003. Antibacterial activity of selected plant essential oils against Escherichia coli O157:H7. Lett. Applied Microbiol., 36: 162-167.
CrossRefPubMedDirect Link