
Haiqing Yu
Triticeae Research Institute, Sichuan Agricultural University, Dujiangyan 611830, Sichuan, China
Chunbang Ding
Triticeae Research Institute, Sichuan Agricultural University, Dujiangyan 611830, Sichuan, China
Chun Zhang
Triticeae Research Institute, Sichuan Agricultural University, Dujiangyan 611830, Sichuan, China
Yonghong Zhou
Laboratory of Medical Molecular Biology, Luzhou Medical College, Luzhou 646000, Sichuan, China
International Journal of Agricultural Research
Year: 2009 | Volume: 4 | Issue: 3 | Page No.: 131-136







ABSTRACT
In order to obtain more cytological data, the karyotypes of Pseudoroegneria gracillima and P. kosaninii were investigated. Root tips of P. gracillima and P. kosaninii were pretreated in an ice bath, fixed in a mixture of 95% ethanol: glacial acetic acid and treated in 1 M HCl at 60°C in a water bath. Somatic cells were stained in Schiff at room temperature and the meristematic portions of the root tips were squashed in 45% acetic acid. The results show that: (1) P. gracillima is diploid with two pairs of satellites and P. kosaninii is octoploid with three pairs of satellites. The karyotypes of diploid P. gracillima and octoploid P. kosaninii are first reported, (2) the karyotype formulas of P. gracillima and P. kosaninii are 2n = 2x = 14 = 12 m (2sat)+2sm (2sat) and 2n = 8x = 56 = 42 m (6sat)+12sm+2st, respectively and (3) the karyotype of P. gracillima is 1A type, while P. kosaninii is 2B type. This demonstrated that there are great variations between the karyotypes of P. gracillima and P. kosaninii.
PDF Abstract XML References Citation
How to cite this article
Haiqing Yu, Chunbang Ding, Chun Zhang and Yonghong Zhou, 2009. Karyotype Studies on Pseudoroegneria gracillima and P. kosaninii (Poaceae: Triticeae). International Journal of Agricultural Research, 4: 131-136.
DOI: 10.3923/ijar.2009.131.136
URL: https://scialert.net/abstract/?doi=ijar.2009.131.136
DOI: 10.3923/ijar.2009.131.136
URL: https://scialert.net/abstract/?doi=ijar.2009.131.136
INTRODUCTION
Pseudoroegneria is a genus in Triticeae (Poaceae) with Pseudoroegneria strigosa (M. Bieb.) a. Löve as the type species (Love, 1980). The genus contains a basic St genome, with diploid (2n = 2x = 14, StSt) and auto- and allo-polyploid species. St genome is one of the most important genomic components, present in more than half of the perennial Triticeae species (Love, 1984; Dewey, 1984; Yen and Yang, 1990; Yen et al., 2005a, b). Morphologically, the species in this genus are caespitose, long-anthered and cross-pollinating perennials. They are distributed in the Northern Hemisphere, with its species occurring on open rocky hillsides from the Middle East and Transcaucasia across Central Asia and Northern China to Western North America (Love, 1984). Pseudoroegneria grasses have exceptionally drought tolerant and excellent forage quality, which are precious germplasm resources in crop forage breeding (Dewey, 1984).
Pseudoroegneria gracillima (Nevski) a. Löve and Pseudoroegneria kosaninii (Nabelek) a. Löve are two species of Pseudoroegneria which distributed in Russian Federation and Turkey, respectively. Love (1984) treated them into Pseudoroegneria based on the morphological study and he noted that the taxonomic status of the two species is temporary with lacking of cytological data. Ding et al. (2004) reported the karyotype of P. gracillima, whereas no cytological data about P. kosaninii are reported. Pseudoroegneria kosaninii is inferred to be tetraploid and the relationship of P. gracillima and P. kosaninii is close in the RAPD and RAMP analysis (Ding et al., 2005a, b). Based on genome specific RAPD markers, P. gracillima and P. kosaninii contained at least one St or slightly modified St genome (Ding et al., 2005c). Yu et al. (2008) suggested that P. gracillima and P. kosaninii contain one St genome in the analysis of the ITS data of the species in Pseudoroegneria. Therefore, the chromosome numbers and genomic constitutions of P. gracillima and P. kosaninii are still obscure. To obtain more cytological data, karyotypes of the two species were investigated. The aims of this study are (1) to report the karyotypes of P. gracillima and P. kosaninii and (2) to provide more cytological data for the appropriate taxonomic treatments of the two species.
MATERIALS AND METHODS
The study was conducted in July 2007 at Dujiangyan City, Triticeae Research Institute of Sichuan Agricultural University. The materials used in this study are shown in Table 1. Seeds of P. gracillima and P. kosaninii were kindly provided by American National Plant Germplasm System (Pullman, Washington, USA). The two species are currently growing at Triticeae Research Institute, Sichuan Agricultural University, China (SAUTI) and the mature plants were carefully identified and determined by Chi Yen, Junliang Yang and Yonghong Zhou.
Seeds were scarified and germinated in Petri dishes at 22°C on filter paper. Root tips were obtained from roots that were 1.0-1.5 cm in length and pretreated in an ice bath for 24 h before fixation in a mixture of 95% ethanol: glacial acetic acid (3:1, v/v) for 24 h. They were then treated for 8-10 min in 1 M HCl at 60°C in a water bath. Somatic cells were stained in Schiff at room temperature (20-25°C) for about 30 min. The meristematic portions of the root tips were squashed in 45% acetic acid. Microphotographs were taken from metaphase cells with a complete chromosome complement by the Olympus BX-51 camera system. Five metaphase cells were analyzed for each species (Li and Chen, 1985). Idiograms were constructed based on the chromosome lengths and relative arm ratios. Index of the karyotypic asymmetry and karyotype type analyses were basically the same as described by Arano (1963) and Stebbins (1971), respectively.
RESULTS
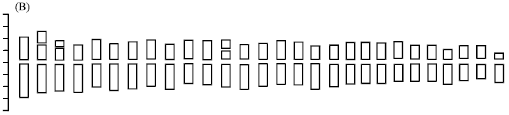
Chromosomal characteristics and chromosomal parameters are shown in Table 2 and 3, respectively. The morphology of somatic chromosomes and karyotypes are shown in Fig. 1A-D. The idiograms are shown in Fig. 2A and B. Results of karyotype analysis of two species studied are presented below.
P. gracillima
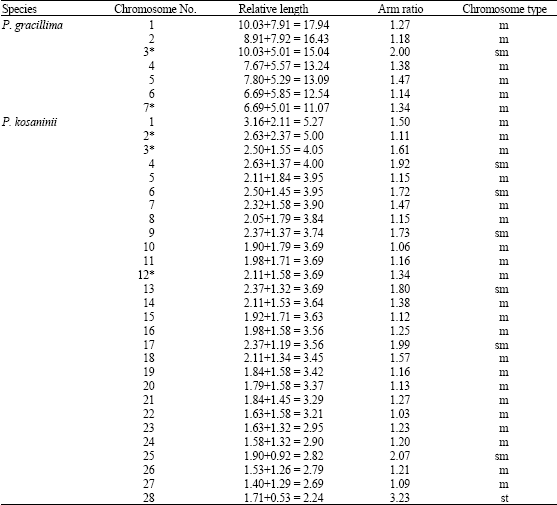
The karyotype formula is 2n = 2x = 14 = 12 m (2sat)+2sm (2sat), which belongs to 1A type. The average arm ratio is 1.40, with a longest chromosome/shortest chromosome ratio of 1.62. Percentage of chromosomes with arm ratio >2 is 0 and index of the karyotypic asymmetry is 58.20. The relative length of chromosomes ranges in size from 11.07 to 17.94. All the chromosomes are metacentric with the exception of chromosome 3, which is submetacentric. One pair of minute satellites and one pair of large satellites are located on the short arms of chromosome 3 and 7, respectively (Fig. 1A, 2A).
| Table 1: | Materials used in the karyotype analysis |
 | |
| Table 2: | Chromosomal parameters of two species in the karyotype analysis |
 | |
| *Satellite chromosome, with the satellite length included in the chromosome length | |
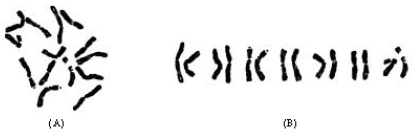 | |
| Fig. 1: | The morphology of somatic chromosomes and karyotypes of two species in Pseudoroegneria (A, B) P. gracillima and (C, D) P. kosaninii. Bar = 10 μm |
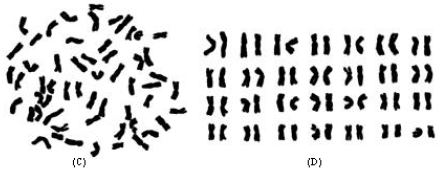
| Table 3: | Chromosomal characteristics of two species in the karyotype analysis |
| AAR: Average arm ratio; Lc: Longest chromosome; Sc: Shortest chromosome; PCA: Percentage of chromosomes with arm ratio >2; As.k (%): Index of the karyotypic asymmetry | |
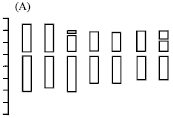 | |
Fig. 2: | Idiograms of two species in Pseudoroegneria (A) P. gracillima and (B) P. kosaninii |
P. kosaninii
The karyotype formula is 2n = 8x = 56 = 42 m (6sat)+12sm+2st, which belongs to 2B type. The average arm ratio is 1.45, with a longest chromosome/shortest chromosome ratio of 2.35. Percentage of chromosomes with arm ratio >2 is 0.07 and index of the karyotypic asymmetry is 57.98. The relative length of chromosomes ranges in size from 2.24 to 5.27. All the chromosomes are metacentric or submetacentric, with the exception of chromosome 28, which is subtelocentric. Three pairs of large satellites are located on the short arms of chromosomes 2, 3 and 12, respectively (Fig. 1B, 2B).
DISCUSSION
Based on the cytological data, Dewey (1984) pointed out that some Pseudoroegneria species may have diploidy and tetraploidy in different populations, such as the diploid (2n = 2x = 14, StSt) P. strigosa and tetraploid (2n = 2x = 28, StStStSt) P. strigosa. The previous analysis indicated that P. gracillima is tetraploid with one pair of satellites (Ding et al., 2004). Pseudoroegneria gracillima and P. kosaninii is closely related and P. kosaninii is inferred to be tetraploid based on molecular data (Ding et al., 2005a, b). However, the ploidy of P. kosaninii is lack of cytological evidence. In the present study, the karyotypes of diploid P. gracillima and octoploid P. kosaninii were first reported. Combined with the earlier study, P. gracillima have diploidy and tetraploidy in different populations. The results added new cytological data of Pseudoroegneria species.
Stebbins and Pun (1953) indicated that different versions of the St genome exist in diploid Pseudoroegneria species, which suggested the differentiations of the St genome. Hsiao et al., (1986) reported that five diploid Pseudoroegneria species have two pairs of satellites. One pair of large satellites are on the short arms of chromosomes 5, the other pair of small satellites are on the short arms of chromosomes 1, 2, or 3. In this study, diploid P. gracillima have one pair of small satellites on the short arms of chromosomes 3 and one pair of large satellites on the short arms of chromosomes 7, which is different to the reported five Pseudoroegneria species. Satellites on different chromosomes displayed the variations in karyotype formulas among diploid Pseudoroegneria species. It is valid that different versions of the St genome have different karyotype formulas. The karyotype of diploid P. gracillima is 1A type and most of the chromosomes are metacentric or submetacentric and the similar results were obtained in diploid Pseudoroegneria species and tetraploid P. gracillima (Hsiao et al., 1986; Ding et al., 2004).
The karyotype analyses indicated that P. kosaninii is octoploid species, which is a higher ploidy than that of other Pseudoroegneria species (Dewey, 1984; Hsiao et al., 1986). Also, there are great variations between the karyotypes of P. kosaninii and other Pseudoroegneria species, especially in sizes and positions of the satellites (Hsiao et al., 1986; Liu and Wang, 1993; Ding et al., 2004). Therefore, more evidence needs to be obtained to make clear the taxonomic status and phylogenetic relationships of P. kosaninii in Pseudoroegneria.
ACKNOWLEDGMENTS
The authors are thankful to the National Natural Science Foundation of China (No. 30670150, 30470135). We particularly thank American National Plant Germplasm System for providing seeds.
REFERENCES
- Ding, C.B., Y.H. Zhou, Y.L. Zheng, R.W. Yang and X.H. Wei, 2004. Karyotypes of six species and two subspecies in Pseudoroegneria and three species in Roegneria. Acta Phytotax. Sin., 42: 162-169.
Direct Link - Ding, C.B., Y.H. Zhou, R.W. Yang, L. Zhang and Y.L. Zheng, 2005. Relationships among Pseudoroegneria species based on randon amplified polymorphic DNA (RAPD). Acta Pratacul. Sin., 14: 38-43.
Direct Link - Ding, C.B., Y.H. Zhou, R.W. Yang, L. Zhang and Y.L. Zheng, 2005. Relationships among Pseudoroegneria species based on RAMP. Acta Bot. Yunnan., 27: 163-170.
Direct Link - Ding, C.B., Y.H. Zhou, R.W. Yang, L. Zhang and Y.L. Zheng, 2005. The genome constitutions of some Pseudoroegneria species assessed by genome-specific RAPD markers. Guihaia, 25: 249-253.
Direct Link - Hsiao, C., R.R.C. Wang and D.R. Dewey, 1986. Karyotype analysis and genome relationships of 22 diploid species in the tribe Triticeae. Can. J. Genet. Cytol., 28: 109-120.
Direct Link - Liu, Z.W. and R.R.C. Wang, 1993. Genome analysis of Elytrigia caespitosa, Lophopyrum nodosum, Pseudoroegneria geniculata ssp. scythica and Thinopyrum intermedium (Triticeae: Gramineae). Genome, 36: 102-111.
CrossRefPubMedDirect Link - Stebbins, G.L., 1971. Chromosomal Evolution in Higher Plants. 1st Edn., Edward Arnold Publisher Ltd., London, UK., Pages: 216.
Direct Link - Yen, C., J.L. Yang and B.R. Baum, 2005. Douglasdeweya: A new genus, with a new species and a new combination (Triticeae: Poaceae). Can. J. Bot., 83: 413-419.
Direct Link - Yen, C., J.L. Yang and Y. Yen, 2005. Hitoshi Kihara, Askell Love and the modern genetic concept of the genera in the tribe Triticeae (Poaceae). Acta Phytotax. Sin., 43: 82-93.
Direct Link - Yu, H.Q., X. Fan, C. Zhang, C.B. Ding, X.L. Wang and Y.H. Zhou, 2008. Phylogenetic relationships of species in Pseudoroegneria (Poaceae: Triticeae) and related genera inferred from nuclear rDNA (internal transcribed spacer) sequences. Biologia, 63: 498-505.
CrossRefDirect Link