
Manal M. Hefny
Department of Agronomy, Faculty of Agriculture, Suez Canal University, Ismailia, 41522, Egypt
A.A. Aly
Department of Agronomy, Faculty of Agriculture, Suez Canal University, Ismailia, 41522, Egypt
International Journal of Agricultural Research
Year: 2008 | Volume: 3 | Issue: 1 | Page No.: 27-39







ABSTRACT
A field experiment was carried out to study the grain yield and Nitrogen Use Efficiency (NUE) components of 16 inbred lines of exotic yellow maize and their crosses. The experiment was cropped at two nitrogen fertilizers; low (70 kg f-1) and high (140 kg f-1), split-plot design was used. The results indicated that, all measured traits were affected significantly by N levels, genotypes and the interaction, except days to 50% tasseling and silking of inbred lines which were not affected by N levels and the N x genotype interaction. N deficiency caused delay in flowering time for male and female inflorescence, accelerated leaf senescence, reduced total dry matter production, N-uptake by plants, grain yield components and grain protein percentage. On the other hand, nitrogen use efficiency for dry matter and grain production and nitrogen harvest index were increased under limited soil N. Inbred lines showed severe reduction for the above variables as compared to crosses. The inbred lines 4, 9, 13 and 15 were distinguished for their superiority in grain yield, nitrogen harvest index, harvest index, nitrogen use efficiency for grain, N-uptake and protein percentage. Three lines, 13, 15 and 16 were the earliest in flowering and represented the highest stay green percentage. While, the inbred lines 1, 8 and 14 were the most N-inefficient for grain production and the lowest for grain yield. In relation to crosses, high nitrogen harvest index, harvest index and nitrogen use efficiency for grain were shown by the crosses (4x1), (8x7), (9x10), (9x12) and (13x15). The crosses (4x1), (9x10), (13x15) and (13x16) surpassed the check and recorded the highest grain yield. The highest stay green percentage was revealed by the single cross Pioneer 3062 followed by the crosses (4x1), (13x15) and (13x16). It is recommended to use the inbred lines; 4, 9, 13 and 15, as a N-efficient source for further studies, whereas using the crosses; (4x1), (8x7), (9x10), (9x12) and (13x15) as N-efficient hybrids for under N limited cultivation. Phenotypic correlation coefficients were higher at low N compared with high application rate. High grain yield was significantly associated with delayed leaf senescence, nitrogen harvest index, harvest index, nitrogen use efficiency for grain and yield plant-1. High nitrogen use efficiency for grain production correlated positively with high yield, yield plant-1, NHI and HI. N-uptake was found to be a function of growth rate at both levels of N fertilizer.
PDF Abstract XML References Citation
How to cite this article
Manal M. Hefny and A.A. Aly, 2008. Yielding Ability and Nitrogen Use Efficiency in Maize Inbred Lines and Their Crosses. International Journal of Agricultural Research, 3: 27-39.
DOI: 10.3923/ijar.2008.27.39
URL: https://scialert.net/abstract/?doi=ijar.2008.27.39
DOI: 10.3923/ijar.2008.27.39
URL: https://scialert.net/abstract/?doi=ijar.2008.27.39
INTRODUCTION
Maize is one of the three main cereal staples, which has to meet an increasing demand for food and feed in the developing world (Cassman et al., 2002). There are important agronomic, economic and ecological reasons for which the primary models of cereal production appear to change for more sustainable agriculture and/or more optimal input technology (El Bassam et al., 1990). Such reasons make search for more efficient cultivars adapted to less favorable nutrition an important breeding task. D`Andrea et al. (2006) stated that, today, the largest investments in maize breeding are made by the private sectors, where the whole selection process (i.e., from early inbred line development to commercial hybrids) takes place in the absence of N restriction. Consequently, the industry does not find it profitable to develop genotypes for areas or markets with low economic return, such as low soil fertility. This may lead to loss of some adaptive traits to these environments.
Growth is a physiological trait associated with grain yield increases in maize plants, moreover is a function of environmental factors (temperature and solar radiation) and mineral nutrition, along with genotype and production practices (Maman et al., 1999).
Worldwide, nitrogen, together with phosphorus is one of the macronutrients that are most limiting to maize grain yield (D`Andrea et al., 2006). The results on the accumulation patterns and redistribution of N-nitrate in maize plants showed that although maize can absorb substantial quantities of N following anthesis and mobilization of vegetative N accumulated before anthesis provides the major source of N in the grain (Di Fonzo et al., 1982). Grain maize NUE is defined as the grain yield per unit of nitrogen available from the soil, including nitrogen fertilizer. The genetic variability and genotype x nitrogen fertilization level interactions for NUE reflect differences in responsiveness have been observed in many studies on maize (Bertin and Gallais, 2000). Accordingly, they suggested that the limiting steps in N-assimilation may be different when plants are grown under different levels of N fertilizers. The post-anthesis N-uptake in grain filling can be related to leaf senescence, because it prolongs the capacity of the plant to absorb mineral N (Racjan and Tollenaar, 1999). The high cost and energy-intensive production of nitrogenous fertilizers and the pollution that resulted from their excessive use necessitate the identification of alternative ways to lessen the dependence on high N inputs (Singh and Arora, 2001). These objectives can be met through efficient farming techniques, but also by using plant varieties that have better Nitrogen Use Efficiency (NUE) (Gallais and Hirel, 2004). Lafitte et al. (1995) suggested that a further progress for low N environments may achieved by selecting N efficient genotypes. So the present study is designed with the aims to: (I) identify new sources for nitrogen use efficiency from exotic materials (ii) compare yielding ability and nitrogen use efficiency related-traits in the parents and the resulting crosses and (iii) describe the relationships between yield and nitrogen use efficiency for grain production and the other tested variables.
MATERIALS AND METHODS
Twelve single-crosses were developed in the 2005 cropping season by crossing sixteen yellow maize inbred lines in a hierarchical mating design (NCD I) of Comstock and Robinson (1948). The inbred lines (Table 1) were introduced from National Plant Germplasm System, USA. The inbred lines were classified into two sets, where each set comprise two male inbred lines crossed to three female inbred lines. One check single cross Pioneer 3062 was included for comparison. The 16 inbred lines and their 12 crosses were sown on 1 May 2006 at the experimental farm of Suez Canal University, Faculty of Agriculture, Ismailia, Egypt. The result of physical and chemical characteristics of experiment`s site showed the values of coarse sand, fine sand, silt and clay are found to be 89.9, 5.7, 2.7 and 1.7%, respectively.
Two levels of nitrogen fertilizer; 70 kg feddan-1 (LN) and 140 kg feddan-1 (HN) (1 ha = 2.4 feddan) were added as ammonium nitrate. Nitrogen was added as single application 3 weeks after planting for the LN treatment, whereas for HN treatment two equal doses were added, 3 weeks after planting and at the beginning of silking. The experiment was laid out in a split-plot design with three replications. The N levels allocated to the main plots, whereas genotypes (12 crosses and 16 parents) distributed in sub-plots. Each genotype (cross and inbred line) was planted in one row, 3 m long, 0.50 m apart and 0.25 m within row.
| Table 1: | List of maize inbred lines and their F1 crosses |
 | |
| IL: Inbred Line | |
Data were recorded on days to 50% tasseling and silking as 50% of the plants in the plot presented their anthers and silks. Stay green (indication of leaf senescence) was determined for each plot by visually assessing the degree of green leaves at 3 wk before harvest. Total Dry matter (TDM) (g m-2) was determined by multiplying total aboveground fresh weight at maturity (stem + leaves) and percentage dry matter of a sub-sample after drying at 70°C for 2 days, Grain yield (GY) (g m-2), was determined by harvesting the ears from the sample area, shelled and weighed. Harvest index (HI), was estimated as the proportion of grain weight to the biological yield (above ground biomass including grains).
Yield per plant was determined by harvesting five individual plants, hand-shelled and grain weighing and Kernel weight was taken on 100 grains,For protein percentage determination, plant parts (grains and aboveground biomass) were ground to pass through a 1 mm sieve for N percentage determination using Kjeldahl procedure (AOAC, 1990), then N percentage was multiplied by 6.25 (Oikeh et al., 1998) for protein percentage calculation.
The following parameters were calculated to estimate Nitrogen Use Efficiency (NUE) (Maranville et al., 1980):
Nitrogen uptake (g N m-2) = TDMxN concentration. |
Nitrogen use efficiency for biomass accumulation (NUEb) = TDM (g)/g N uptake in the aboveground biomass, Nitrogen use efficiency for grain yield (NUEg) = g grain/g N uptake in the aboveground biomass.
Nitrogen harvest index (NHI) was estimated according to Koutroubas and Ntanos (2003) = Ng/Nt
| where: |
| Nt | : | Total aboveground N and |
| Ng | : | Grain N. |
The statistical analyses were performed according to Steel and Torrie (1980) using GenStat Software Package, Release 4.24 to estimate the significance effect of N levels, genotypes and their interactions. LSD values were calculated and used to compare treatment means.
RESULTS AND DISCUSSION
Flowering Dynamics, Stay Green, Biomass Production and N-Uptake
The analysis of variance (Table 2 and 3) indicated that days to 50% tasseling and silking showed significant differences among inbred lines. In case of crosses, days to 50% silking responded significantly with respect to applied nitrogen, crosses and the interactions, whereas days to 50% tasseling were influenced significantly by crosses and nitrogen levels. The overall means for the previous traits are presented in Table 4-9. The limited nitrogen fertilizer did increase the days required to reach flowering phase for inbreds and crosses. Under N stress, plants were earlier in days to 50% silking than days to 50% tasseling. This in turn resulted in an increase in Anthesis-Silking Interval (ASI) (5.93 vs 4.82 days for inbreds and crosses, respectively).
| Table 2: | Mean squares of 16 yellow maize inbred lines cropped at two levels of nitrogen (70 and 140 kg N f-1) |
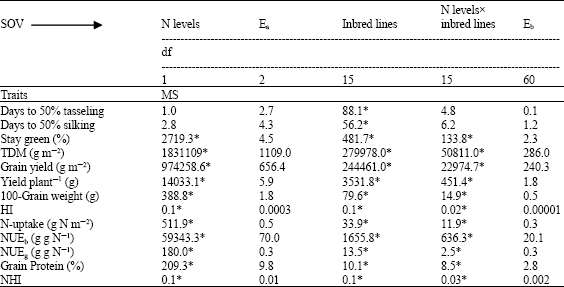 | |
| *, Significant at 0.05 levels of probability, MS: Mean squares | |
| Table 3: | Mean squares of 13 yellow maize single crosses cropped at two levels of nitrogen (70 and 140 kg N f-1) |
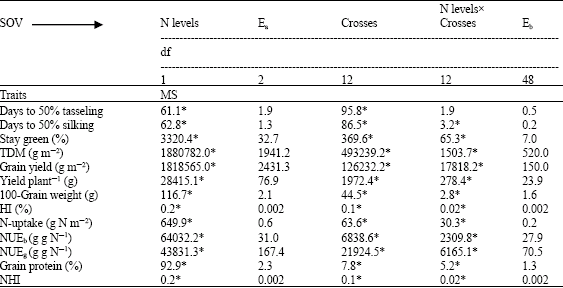 | |
| *: Significant at 0.05 levels of probability, MS: Mean squares | |
| Table 4: | Performance of 16 yellow maize inbred lines cropped at two levels of nitrogen fertilizer (70 and 140 kg N f-1*) for flowering, stay green, total dry matter and N-uptake traits |
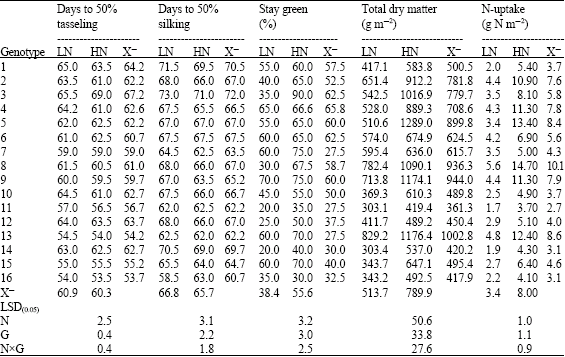 | |
| LN: Low Nitrogen, HN: High Nitrogen and X: Mean | |
| Table 5: | Performance of 16 yellow maize inbred lines cropped at two levels of nitrogen fertilizer (70 and 140 kg N f-1*) for grain yield and its components |
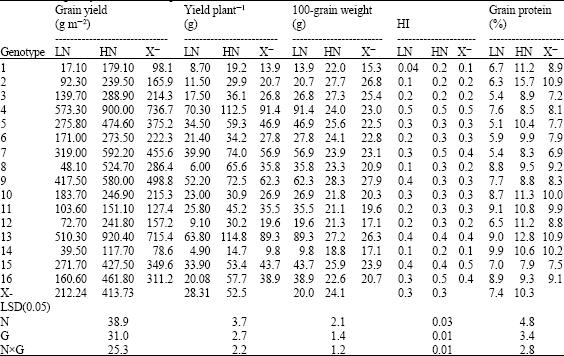 | |
| LN: Low Nitrogen, HN: High Nitrogen and X–: Mean | |
The inbred lines 16 and 13 were among the earliest for anthesis and silking and the inbreds 1 and 3 were the most delayed. The crosses (13x16) and (13x15) followed by the cross (9x11) were the earliest for anthesis and silking. On the other hand the crosses SC Pioneer 3062 and 4x3 were the most delayed. Regarding crosses, a difference of 8 (for anthesis) and 12 days (for silking) was recorded in time to flowering between the early and late groups. For inbreds, there was 11 days difference between the same groups. A set of 12 inbred lines were evaluated under field conditions at two N levels, 0 and 400 kg N ha-1 by D`Andrea et al. (2006), the results recorded an increase in thermal time required to anthesis and silking at reduced level of N. Also, the delay in anthesis was less than in silking which resulted in increasing in ASI but this was not common to all inbreds. The same authors registered 7-10 days differences in flowering time between the early and late flowering groups. Lafitte and Edmeades (1995) confirmed that, increases in ASI is raised when maize plants are subjected to various stresses such as drought or N deficiency.
| Table 6: | Performance of 16 yellow maize inbred lines cropped at two levels of nitrogen fertilizer (70 and 140 kg N f -1) for NUE traits |
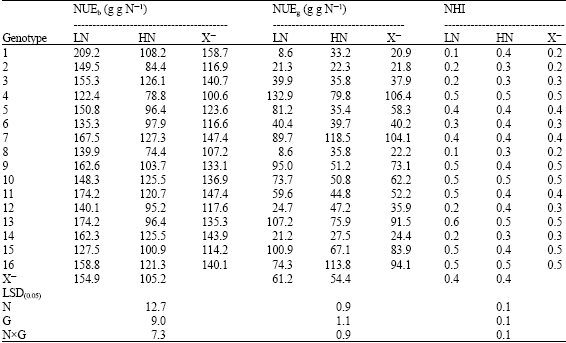 | |
| LN: Low Nitrogen, HN: High Nitrogen and X–: Mean | |
| Table 7: | Performance of 13 yellow maize crosses cropped at two levels of nitrogen fertilizer (70 and 140 kg N f-1) for flowering, stay green, total dry matter and N uptake traits |
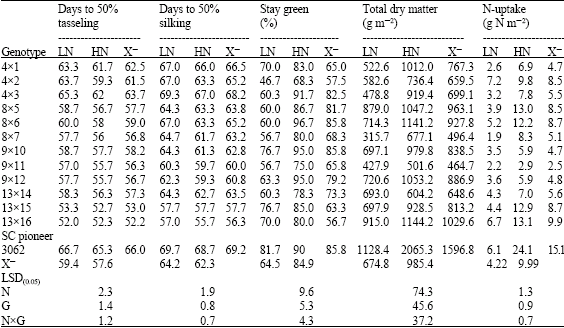 | |
| LN: Low Nitrogen, HN: High Nitrogen and X–: Mean | |
Stay green trait was influenced significantly by applied nitrogen treatments, genotypes (lines and crosses) and their interaction. Similar results were obtained by many authors when maize plants were cultivated under a range of soil N fertilizers. Gungula et al. (2005) found significant differences between N rates (0 to 120 kg N ha-1) and the tested varieties for leaf senescence percentage which an indicator of the effect of soil N on greenness. The reduction in N availability encouraged leaf senescence as shown in Table 4 and 7. Similar results were achieved by different authors; Racjan and Tollenaar (1999) found that leaf longevity was enhanced by an increase in soil N supply. In addition, reduced N availability accelerated post flowering leaf senescence than at high N and maize inbred lines showed differences in their magnitude of response (D`Andrea et al., 2006). Gungula et al. (2005) declared that highest percentage of leaf senescence at the lowest N-level (30 kg N ha-1), while the lowest leaf senescent percentage was recorded at 120 kg N ha-1. In their results, Borrell et al. (2001) established that roots of the stay green sorghum maintain greater capacity to extract N from the soil compared with the non-stay green hybrids during kernel filling. They assessed it as a consequence of the balance between N-demand by the kernel and N-supply during the kernel filling. The inbred lines showed leaf greenness reduction more than their crosses (31.0 and 24.04%, respectively) when grown at N deficiency conditions. The inbreds 4, 5, 6, 7, 9, 13 and 15 and the crosses Pioneer 3062, 9x10, 4x1, 13x16 and 13x15 were characterized by their highest greening percentage at physiological maturity and kernel filling phase at reduced N soil. The previous result can be explained based on Boràas et al. (2003) suggestion that delay in senescing for the previous group of genotypes during kernel filling is linked to the quantity of light received by the leaves and N availability via remobilization to actively growing kernels of maize. In addition, such genotypes maintain their green leaves longer than others and in turn represent differences in photosynthetic capacity (Gungula et al., 2005).
| Table 8: | Performance of 13 yellow maize crosses cropped at two levels of nitrogen fertilizer (70 and 140 kg N f-1) for flowering, stay green, total dry matter and N uptake traits for grain yield and its components |
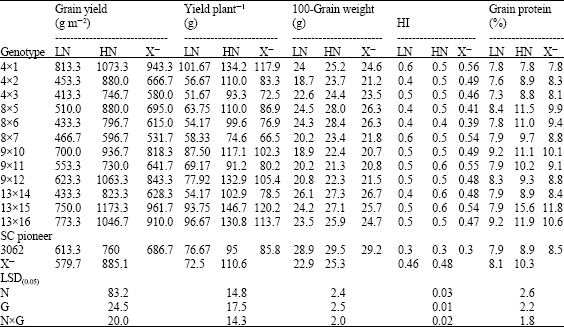 | |
| LN: Low Nitrogen, HN: High Nitrogen and X–: Mean | |
| Table 9: | Performance of 13 yellow maize crosses lines cropped at two levels of nitrogen fertilizer (70 and 140 kg N f-1) for NUE traits |
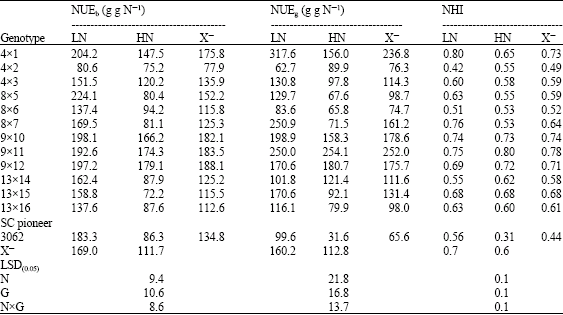 | |
| LN: Low Nitrogen, HN: High Nitrogen and X–: Mean | |
Results in Table 2 and 3 revealed the significant effects of N fertilizer levels, genotypes and the interaction on biomass production. The limited nutrition promoted reduction in biomass production by 34.97 and 31.52% for inbreds and their crosses, respectively. Greater reduction in biomass production and plant growth rate for maize inbreds than for hybrids around silking period was affirmed (Uhart and Andrade, 1995). Their result was related to the reduction in leaf expansion and light interception efficiency as suggested by D`Andrea et al. (2006). Our results are go in line with the idea supported by Tollenaar et al. (2004) in that reduction in biomass production usually observed in inbred lines is more related to the negative effects of inbreeding on leaf expansion and light interception than to photosynthesis levels. The consequence of N stress strengthens this tendency. The inbreds 13, 8, 9 and 2 showed the highest dry matter production at N stress planting, where the production ranged from 26.75-61.42% above the average. The cross Pioneer 3062, showed the highest dry matter production relative to the crosses (13x16), (8x5) and (9x12) at 70 kg N f-1, with registered 6.79-67.22% increase over general mean (Table 4-9).
N-uptake was differed significantly by N levels, genotypes and their interaction (Table 2 and 3). Nitrogen accumulation increased from 4.22 to 9.99 and from 3.37 to 7.99 g N m-2 in crosses and their inbred lines respectively with application of 140 kg N feddan-1. Differences in N-uptake due to hybrid effects were twofold higher than that of inbreds. The lines 8, 13, 9, 2, 6 and 4 and the crosses (4x2), (13x16), Pioneer 3062 and (8x6) exhibited the highest N uptake at N limited nitrogen supply. It is clear that total dry matter production at harvest is correlated and affected by N-uptake at both levels of N (Fig. 1 and 2) in hybrids and inbred lines. Singh and Arora (2001), confirmed the same results for wheat genotypes cultivated at 40 and 120 kg N ha-1 and mentioned that rate of dry matter production may control nitrogen uptake and growth is tightly controlled by nitrogen supply. Thus demand factor is an important determinant of uptake rate.. In contrast, inbreds 11 and 14 and the hybrid (8x7) accumulated the least dry matter and had the lowest N uptake.
Grain Yield Determinants
As shown in Table 2 and 3, Nitrogen levels, genotypes and N levelsxgenotypes were significant for grain yield, yield plant-1, 100-grain weight and HI of hybrids and inbred lines. Mean squares due to N levels were highest than those due to genotypes for both groups when estimated for all traits except HI of hybrids. The significant effect of genotypexN level interactions on the measured traits confirms that genotypes perform differently under different N application. The inbred lines group showed severe reduction in grain yield, yield plant-1, 100-grain weight and HI (48.70, 46.06, 16.75 and 21.21%, respectively) when planted under N limited input conditions compare with hybrid group (34.50, 34.50, 9.64 and 4.17%, respectively). Present results support the data achieved by Betrán et al. (2003) in that inbred lines are relatively more sensitive to low soil nutrition compare with hybrids and grain yield for maize inbred lines grown at reduced N recorded 65% reduction of that under high N. Similarly, Gallais and Hirel (2004) recorded 38, 32 and 9% reduction in yield, kernel number plant-1 and kernel weight. Plant grain yield was strongly reduced by N deficiency relative to grain weight, D`Andrea et al. (2006) established that yield plant-1 is severely reduced by low N application and such response is mainly related to variation in kernel number per plant, whereas kernel weight is less susceptible to N stress and grain yield is less responsive to its variation (Gallais and Hirel, 2004; D`Andrea et al., 2006).
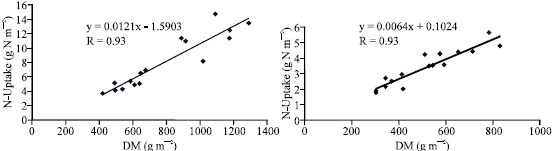 | |
| Fig. 1: | Relation between N-Uptake and DM for 16 maize inbred lines cropped at (a): 70 and (b): 140 kg N feddan-1 |
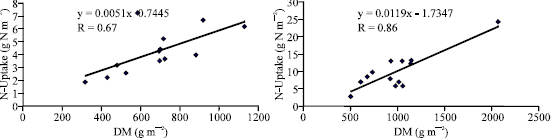 | |
| Fig. 2: | Relation between N-Uptake and DM for 13 maize single crosses cropped at (a): 70 and (b): 140 kg N feddan-1 |
The inbred lines 4, 13, 9, 7, 5 and 15 produced the highest grain yield, yield plant-1, HI and grain weight under low N input and the range in grain yield reduction varied between 28-46%. Although the lines 10 and 11 recorded low yield at stress condition, both were not sensitive to low N (25.60 and 31.44%, respectively) (Table 5). On the other hand, the best hybrids for the previous traits were (4x1), (13x16), (13x15), (9x10) and (9x12), which surpassed the check SC pioneer 3062 (Table 8). HI is the proportion of grain yield and biological yield, accordingly, inbreds with high HI has the capacity to accumulate total amount of aboveground dry matter and translocate it to the growing grains. So the inbred lines with the highest HI are those with high yielding ability. D`Andrea et al. (2006) reported two cases of HI reduction in maize inbred lines, the slight decrease which attribute to the effect of N deficiency on the relationship between plant growth rate change and kernel number per plant. Beside the strong reduction which includes inherent biomass partitioning pattern of the genotype around silking and N availability controlling. It is suggested that there is reduced efficiency for converting dry matter at anthesis to reproductive sinks under N stress (Abbate et al., 1995), this in turn is represented in the effect of N stress on the reduced HI in some maize hybrids.
Nitrogen Use Efficiency, Grain Protein Percentage and Nitrogen Harvest Index
There were statistically significant differences (Table 2 and 3) between N levels, genotypes and their interaction for NUEb, NUEg, grain protein percentage and the proportion of total plant N in the grains at maturity (NHI). Under soil limited fertility, protein percentage reduced in grains by 28.5 and 21.21% for inbred lines and crosses, respectively. The results showed that, N stress caused 12.19% reduction and 6.25% increase in NHI for inbred lines and hybrids, respectively.
Regarding NUEb and NUEg, N stress increased NUE for plant biomass and grain yield by 47.29 and 12% vs 51.30 and 42.02%, for inbred lines and crosses, respectively. It is apparent that NUEb for inbred lines and their crosses was nearly close, but a wide difference exists among both groups for NUEg. The ranges in NUE for dry matter and grain yield for inbred lines were too low compared with their crosses. This may be due to (Table 4-9) the effect of inbreeding depression on these traits, added to that, inbred lines represented more efficiency for biomass production than grain yield production. Only five inbred lines 4, 13, 15 and 9 and 7 had the highest NUEg and NHI. However for NUEb seven inbred lines (1, 2, 3, 6, 8, 12 and 14) showed the highest NUEb values accompanied with low NHI (Table 6). As already confirmed by Gallais and Hirel (2004), maize genotypes exhibiting low agronomic performance at low N-input, i.e., those having low NUE, are those reacting more to nitrogen application. Therefore, genotypexnitrogen appears to be essentially due to variation in the adaptation of the plant to low N-input.
As indicated in Table 9, the crosses: (4x1), (9x10), (8x7) (9x12), (13x15) and (9x11) combined high NHI and NUEg, thus such crosses are characterized by efficient partitioning of dry matter to grains and producing grains per unit of plant nitrogen at lower nitrogen level. The crosses (8x5), (4x1), (9x10), (9x12), (9x11) were superior and efficient in producing high dry matter per unit of applied N compared to Pioneer 3062. The hybrids, (8x6) and (4x2) were characterized by their N-inefficiency because of their poor NUEg and NHI. The previous results are supported by studies carried out by many researchers. Webb et al., (1998) related the increase in N accumulation of shoots and reduction in NHI in all wheat genotypes with N fertilization to the less efficient use of N. In contrast, Oikeh et al. (2007) recorded 20% reduction in NHI for maize cultivars grown at 0 compared to 120 kg N ha-1. They also recorded a large NHI value of 0.63 for N-use efficient maize cultivars. Koutroubas and Ntanos (2003) attributed the high NUEg for Indica compared to Japonica cultivars of rice to their high N translocation from the vegetative tissues to grains during the post-anthesis period and thus increase NHI. They also added that, increasing in NHI and HI favored high NUEg. Singh and Arora (2001) confirmed a decline in NUE for grain and dry matter production in wheat cultivars as the N rate was increased from 40 to 120 kg N ha-1.
Relationship Between Grain Yield, NUEg and the Other Measured Variables
The phenotypic correlation coefficients (rp) measured between grain yield and NUEg and the other tested traits for inbred lines and crosses at both levels of N fertilizer. At 70 kg N, grain yield of inbred lines correlated negatively with number of days to 50% tasseling and silking. Protein percentage although correlated negatively with grain yield, the value was small (Table 10). On the other hand, grain yield recorded positive and significant values with yield plant-1, stay green, NUEb, NHI, HI and grain weight. The results are in accordance with those established by Betrán et al. (2003) in that negative phenotypic correlation exist between maize grain yield and male and female flowering dates in both inbreds and their hybrids across a range of stress and non stress environments. They affirmed that such earlier genotypes could escape the intense stress at flowering. The same authors recorded negative and significant phenotypic correlation values between senescence and maize grain yield for hybrids (-0.38) and inbreds (-0.48). Grain yield recorded weak correlation with N-uptake. Under optimal conditions of N fertilization, there were significant relationship between yield and yield plant-1, N-uptake, NUEg and HI. Regarding NUEg, a significant and positive relation was recorded with NHI, HI, grain yield and stay green, the values were higher at N stress compared with conditions of available N.
| Table 10: | Phenotypic correlation (rp) between grain yield (g m-2), NUEg (g g N-1) and the other tested variables at low and high nitrogen applications |
 | |
| *,**: Significant at 0.05 and 0.01, respectively, GY: Grain Yield, NUEg: Nitrogen use efficiency for grain production | |
Data for crosses showed positive and significant phenotypic correlation values between yield; greenness, protein percentage and NHI. Whereas, positive and significant relationship were recorded between NUEg and NHI and HI. Grain yield did not associate with N-uptake compared with values recorded for inbred lines. This may give the crosses the advantage of the ability of N translocation from the stover to the grain. The correlation values were higher at stress conditions, since at optimal level of N no significant values were recorded. NUEg showed positive and significant relationships with NHI and HI at both low and high N-input. In similar way, Koutroubas and Ntanos (2003) recorded positive association between NUEg, HI, NHI and explained this relationship on the basis of dry matter and N translocation from the vegetative parts to the grains which raise the HI and NHI in turn.
High grain yield correlated with low protein percentage in inbred lines, but in crosses the relationship was positive and significant. This result shows the advantage of the possibility of selection for increasing grain yield and protein percentage and that some of the current crosses combine both high yield and protein percentage. In addition, the lack of evidence of significant correlation between N uptake and NUE implies that within this pool of inbred lines and crosses it is possible for combining high N uptake efficiency with high internal use efficiency.
The previous results are in accordance with those obtained by different studies on cereal crops. Gungula et al. (2005) recorded a positive association between lower percentage of leaf senescence and high maize grain yield. They attributed such positive relationship to the higher photosynthetic capacity of green leaves and the ability of the plants to capture more solar energy during grain filling. Muurinen et al. (2006) recorded a positive correlation for total plant biomass and grain yield with nitrogen uptake and also observed positive correlation between NUE and HI. Di Fonzo et al. (1982) mentioned that under low level of N fertilizer, maize grain yield is correlated with the NHI, on the other hand.
CONCLUSION
It could be concluded from the previous results that growing maize plants under N stress resulted in the reduction in grain yield and other related traits, but raised the N use efficiently for dry matter and grain yield production. Inbred lines showed recorded mean values for all measured traits compared with their crosses. There is, in some cases, simultaneous decrease in yield and increases in NUEb, since some inbred lines (1, 8 12 and 14) represented low yielding ability although utilized N in efficient way for biomass production. In addition, it is not necessarily the case that a genotype with high nitrogen uptake record high NUE for biomass production (1, 11 and 14 vs 8) or vice versa as confirmed by Singh and Arora (2001). In developing countries the high cost and the little or no access to fertilizer input, makes farmers attempt to use minimal dose, in such cases it is advantage to cropping high N efficient genotypes under limited N input where high N input maize would not give some appreciable returns (Kogbe and Adediran, 2003). In addition, cultivating N-efficient genotypes would reduce the hazards of environmental pollution. Furthermore, some genotypes (inbred lines and crosses) showed differential in N use efficiency for dry matter {1, 11 and 13 and (4x1), (8x5), (9x10), (9x11 and 9x12)} and/or grain production {4, 13, 15 and 9; (4x1), 8x7), (9x11) and (9x10)}, this gives an ample chance to select genotypes to be used either for grain production or as a source for nutritive value for animal feed, as maize stover has high crop residue quality in terms of crude protein and digestibility than sorghum.
The inbred lines and crosses, that showed desirable tendency towards efficient and inefficient use of N, could be further subject to physiological analyses to study the pattern of enzyme activities controlling N-uptake and translocation. Building the relationship between the enzymes activity and responsible proteins in one hand and the intensity of such relationship among the parents and their crosses on the other hand is of important. Also the obtained crosses could be re-evaluated with the local cultivated hybrids at different levels of N in multi-location experiments to compare their yielding ability.
REFERENCES
- Abbate, P.E., F.H. Andrade and J.P. Culot, 1995. The effect of radiation and nitrogen on number of grains in wheat. J. Agric. Sci., 124: 351-360.
Direct Link - Betran, F.J., D. Beck, M. Banziger and G.O. Edmeades, 2003. Genetic analysis of inbred and hybrid grain yield under stress and nonstress environments in tropical maize. Crop Sci., 43: 807-817.
CrossRefDirect Link - Bertin, P. and A. Gallais, 2000. Genetic variation for nitrogen use efficiency in a set of recombinant maize inbred lines. 1. Agro-physiological results. Maydica, 45: 53-66.
Direct Link - Boràas, L., G.A. Maddonni and M.E. Otegui, 2003. Leaf senescence in maize hybrids: Plant population, row spacing and kernel set effects. Field Crops Res., 82: 13-26.
CrossRefDirect Link - Borrell, A.K., G.L. Hammer and E.V. Oosterom, 2001. Stay-green: A consequence of the balance between supply and demand for nitrogen during grain filling? Ann. Applied Biol., 138: 91-95.
CrossRefDirect Link - Cassman, K.G., A. Dobermann and D.T. Walters, 2002. Agroecosystems, nitrogen-use efficiency and nitrogen management. AMBIO: J. Hum. Environ., 31: 132-140.
CrossRefDirect Link - Comstock, R.E. and H.F. Robinson, 1948. The components of Genetic variance in populations of biparental progenies and their use in estimating the average degree of dominance. Biometrics, 4: 254-266.
Direct Link - D'Andrea, K.E., M.E. Otegui, A.G. Cirilo and G. Eyherabide, 2006. Genotypic variability in morphological and physiological traits among maize inbred lines-Nitrogen responses. Crop Sci., 46: 1266-1276.
CrossRefDirect Link - Di Fonzo, N., M. Motto, T. Maggiore, R. Sabatino and F. Salamini, 1982. N-uptake, translocation and relationships among N related traits in maize as affected by genotype. Agronomie, 2: 789-796.
Direct Link - Maranville, J.W., R.B. Clark and W.M. Ross, 1980. Nitrogen efficiency in grain sorghum. J. Plant Nutr, 2: 577-589.
CrossRefDirect Link - Gallais, A. and B. Hirel, 2004. An approach to the genetics of nitrogen use efficiency in maize. J. Exp. Bot., 55: 295-306.
Direct Link - Gungula, D.T., A.O. Togun and J.G. Kling, 2005. The influence of n rates on maize leaf number and senescence in Nigeria. World J. Agric. Sci., 1: 1-5.
Direct Link - Kogbe, J.O.S. and J.A. Adediran, 2003. Influence of nitrogen, phosphorus and potassium application on the yield of maize in the Savanna zone of Nigeria. Afr. J. Biotechnol., 2: 345-349.
Direct Link - Koutroubas, S.D. and D.A. Ntanos, 2003. Genotypic differences for grain yield and nitrogen utilization in Indica and Japonica rice under Mediterranean conditions. Field Crops Res., 83: 251-260.
Direct Link - Lafitte, H.R. and G.O. Edmeades, 1995. Association between traits in tropical maize inbred lines and their hybrids under high and low nitrogen. Maydica, 40: 259-267.
Direct Link - Lafitte, H.R., G.O. Edmeades and S. Taba, 1995. Adaptative strategies identified among tropical maize landraces for nitrogen-limited environments. Field Crops Res., 49: 187-204.
Direct Link - Muurinen, S., G.A. Slafer and P. Peltonen-Sainio, 2006. Breeding effects on nitrogen use efficiency of spring cereals under Northern conditions. Crop Sci., 46: 561-568.
Direct Link - Oikeh, S.O., J.G. Kling and A.E. Okoruwa, 1998. Nitrogen fertilizer management effects on maize grain quality in the west Africa moist savanna. Crop Sci., 38: 1056-1061.
CrossRefDirect Link - Oikeh, S.O., V.O. Chude, G.J. Kling and W.J. Horst, 2007. Comparative productivity of nitrogen use efficient and nitrogen-inefficient maize cultivars and traditional grain sorghum in the moist Savanna of West Africa. Afr. J. Agric. Res., 2: 112-118.
Direct Link - Maman, N., S.C. Mason, T. Galusha and M.D. Clegg, 1999. Hybrid and nitrogen influence on pearl millet production in Nebraska: Yield, Growth and nitrogen uptake and nitrogen use efficiency. Agron. J., 91: 737-743.
Direct Link - Singh, V.P. and A. Arora, 2001. Intraspecific variation in nitrogen uptake and nitrogen utilization efficiency in wheat (Triticum aestivum L.). J. Agron. Crop Sci., 186: 239-244.
Direct Link - Steel, R.G.D. and J.H. Torrie, 1980. Principles and Procedures of Statistics: A Biometrical Approach. 2nd Edn., McGraw Hill Book Co., New York, USA., ISBN-13: 9780070609266, Pages: 633.
Direct Link - Tollenaar, M., A. Ahmadzadeh and E.A. Lee, 2004. Physiological basis of heterosis for grain yield in maize. Crop Sci., 44: 2086-2094.
Direct Link - Uhart, S.A. and F.H. Andrade, 1995. Nitrogen deficiency in maize: I effects on crop growth, development, dry matter partitioning and kernel set. Crop Sci., 35: 1376-1383.
Direct Link - Webb, J., F.M. Seeney and B.R. Sylvester, 1998. The response to fertilizer nitrogen of cereals grown on sandy soils. J. Agric. Sci., 130: 271-286.
Direct Link - Racjan, I. and M. Tollenaar, 1999. Source-sink ratio and leaf senescence in maize. II. Nitrogen metabolism during grain filling. Field Crops Res., 60: 255-265.
Direct Link