
A.H. Hanafy Ahmed
Plant Physiology Section, Plant Botany Department, Faculty of Agriculture, Cairo University, Egypt
E.M. Harb
Plant Physiology Section, Plant Botany Department, Faculty of Agriculture, Cairo University, Egypt
M.A. Higazy
Plant Physiology Section, Plant Botany Department, Faculty of Agriculture, Cairo University, Egypt
Sh.H. Morgan
Plant Physiology Section, Plant Botany Department, Faculty of Agriculture, Cairo University, Egypt
International Journal of Agricultural Research
Year: 2008 | Volume: 3 | Issue: 1 | Page No.: 1-26







ABSTRACT
Two pot experiments were carried out under different soil salinity levels (0, 2000, 4000 and 6000 ppm), as well as field experiment under saline soil condition, to investigate the effect of spraying silicon (0, 250 and 1000 ppm SiO2) and/or boron (0 and 25 ppm B) sprayed twice (40 and 70 days after sowing) under pots experiment, as well as spraying silicon (0, 250, 500 and 1000 ppm SiO2) and/or boron (0, 25 and 50 ppm B) sprayed three successive times (at 35, 60 and 85 days after sowing) under field experiment on growth, yield and chemical composition of wheat (Triticum aestivum L.) var. Seets 1. Generally, under pots experiment, both silicon levels either alone or combined with boron significantly increased shoots height and leaf area as well as grains yield/plant and weight of 1000. However, under field experiment only the lowest level of silicon significantly increased all the studied growth characters, while all levels of silicon significantly increased number of spikes and grains as well as grains yield when compared with control non-sprayed plants, however, the lowest level of silicon had the superiority effect. Both silicon and boron applications correct to some extent the negative effects of salinity either on growth, yield, nutrients uptake, free polyamines and endogenous plant hormones (gibberellic acid and cytokinins) while decreased abscisic acid. However, no constant trend was obtained for indole-3-acetic acid concentration.
PDF Abstract XML References Citation
How to cite this article
A.H. Hanafy Ahmed, E.M. Harb, M.A. Higazy and Sh.H. Morgan, 2008. Effect of Silicon and Boron Foliar Applications on Wheat Plants Grown under Saline Soil Conditions. International Journal of Agricultural Research, 3: 1-26.
DOI: 10.3923/ijar.2008.1.26
URL: https://scialert.net/abstract/?doi=ijar.2008.1.26
DOI: 10.3923/ijar.2008.1.26
URL: https://scialert.net/abstract/?doi=ijar.2008.1.26
INTRODUCTION
Soil salinity is a major problem for agriculture throughout the world. The problem of salinity is of special importance in Egypt for both the old cultivated area as well as for the newly reclaimed lands. The major constraints for plant growth and productivity are ion toxicity with excessive uptake of mainly Cl- and Na+ as well as nutrients imbalance caused by disturbed uptake or distribution of essential mineral nutrients (Hu and Schmidhalter, 2005).
Silicon was reported to reduce the hazard effects of various abiotic and biotic stresses including salt stress, metal toxicity, drought stress, radiation damage, various pests and diseases caused by both fungi and bacteria, nutrients imbalance, high temperature and freezing (Ma, 2004). Usually, plants grown on saline soil suffer from several nutritional disturbances and nutrient deficiencies and consequently plant growth inhibition. In this respect, Hanafy Ahmed et al. (2002b) working on wheat plants grown under different salinity levels (0, 3000 and 6000 ppm) noted that, growth characters of wheat plants had pronounced increased by adding silicon under non-saline or saline soil conditions.
Moreover, it`s well known that under salinity conditions boron (B) concentration was decreased in plants (Holloway and Alston, 1992; Wimmer et al., 2001). Boron is an important micro nutrient known to exhibit several interactions with other plant nutrients.
Thus, the aim of the present study was to investigate the effect of Si and B foliar applications as well as their combination on growth, yield and chemical composition of wheat, aiming to improve wheat yield under non-saline or saline soil conditions.
MATERIALS AND METHODS
Pot experiments were carried out in the wire house of the Plant Physiology Section, Faculty of Agriculture, Cairo University, Giza, Egypt, during the two successive seasons, 2001-2002 and 2002-2003. Plastic pots (30 cm in diameter) were filled with 10 kg clay loam soil obtained from the Farm of Faculty of Agriculture, Giza. Field experiment was carried out in Smosta, Baneswaf governorate during season 2003-2004. Mechanical analysis for pot experiments were determined according to the methods reported by Richards (1954). Chemical analyses of soils for pot and field experiments were determined according to the methods reported by Jackson (1973) (Table 1).
In the two successive seasons, pots were divided into 3 main groups. The first main group was sprayed with fresh water (control A) and the other two main groups were sprayed with silicon as sodium meta silicate (Na2SiO3.5H2O) at concentration 250 or 1000 ppm SiO2, respectively. Each main group was divided into two sub-groups. The first sub-group was sprayed with boron as boric acid (H3BO3) at concentration 25 ppm B, while the second sub-group was sprayed with water. Each sub-group of pots was divided into 4 sub-sub-groups of salinity levels (0, 2000, 4000 and 6000 ppm). Soil salination was made by adding mixture of sodium chloride, calcium chloride and magnesium sulphate at the ratio of 1:1:0.5 by weight. Each pot received 2.2 g of calcium superphosphate (15.5% P2O5) and 1.1 g potassium sulphate (48% K2O) before sowing and 3.0 g ammonium nitrate (33.5% N) added two weeks after sowing. In each pot, 10 grains of wheat (Triticum aestivum L.) var. Seets 1 were sown on the 30th Nov. 2001 in the first season and on the 29th Nov. 2002 in the second one. The seedlings were thinned to 6 plants per pot two weeks later after sowing. Seeds were weekly irrigated with fresh water keeping it not more than field capacity. Different foliar treatments were applied at 40 and 70 days after sowing.
Field affected with salinity (4400 ppm) was parted into 3 main plots. The first main plot sprayed with water whereas the second and third main plots were sprayed with boric acid at concentrations 25 and 50 ppm B, respectively. Each main plot was splited into 4 sub plots. The first sub plot was sprayed with fresh water (control A) and the other 3 sub plots were sprayed with silicon at the rates of 250, 500 and 1000 ppm SiO2, respectively. Fertilization was carried out according to recommendation of Ministry of Agriculture; calcium superphosphate (15.5% P2O5) and potassium sulphate (48% K2O) were added before planting at the rate of 150 and 50 kg/fed., respectively. Ammonium nitrate (33.5% N) were added before the first irrigation at the rate of 300 kg/fed. Wheat grains were sown on 24th Nov. 2003. Foliar application treatments were applied at three successive times 35, 60 and 85 days after sowing.
In both successive seasons of pots and in field experiment, one plant sample was taken 90 days after sowing. The following measurements were recorded: shoot height (cm), number of tillers/plant, leaf number, leaf area (cm2) and shoot dry weight (g). At harvest (160 days after sowing), yield and yield components were estimated as follows: number of spikes and grains per plant, grains as yield g/plant in pots and ardab/fed in field, straw yield as g/plant in pots and ton/fed in field as well as weight of 1000 grains (g).
| Table 1: | Mechanical and chemical properties of the used soils |
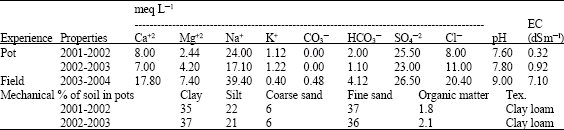 | |
Chemical Analysis
Determinations of N, P, K, Ca, Na and Mg as well as proline were carried out on the ground dry material. Nitrogen concentration was determined by Nessler method according to AOAC (1960). Phosphorus was estimated colorimeterically using the chlorostannous reduced molybdophosphoric blue color method according to Jackson (1973). Potassium and sodium were determined using the flame photometer (CORNING M 410). Calcium and magnesium were determined using the atomic absorption spectrophotometer (GBC, 932 AA). Free proline concentration was measured colorimeterically using ninhydrin reagent (Bates et al., 1973). Total chlorophyll in fresh leaves were extracted with dimethyl formamide and calculated as described by Nornai (1982).
Ethanol extract of shoots were used for the determination of total sugars, total free amino acids and total soluble phenols. Total sugars were determined by using the phenol sulphuric acid reagent (Dubois et al., 1956). Total free amino acids were determined by using ninhydrin reagent (Moore and Stein, 1954). Total soluble phenols were determined by using the Folin-Denis colorimetric method (Swain and Hillis, 1959).
Free polyamines as putrescine (Put) was only determined in second season of pot experiment as well as in field experiment by dansyl chloride reagent and quantified with spectrophotofluroimeter according to Galston (1983).
Extraction of plant hormones was only done in second season of pot experiment, according to Sadeghian (1971). The alkaline fraction was used for the determination of cytokinins according to El-Ghamrawy and Neumann (1977) while the acidic fraction was used for determination of gibberellic acid, abscisic acid and indole-3-acetic acid according to Vogel (1975) using Gas-Liquid Chromatography (GLC).
Data of pot experiments were statistically analyzed by using three factorial completely randomized design, while data of field experiment were statistically analyzed using a split plot design which boron foliar application was the main plot and silicon foliar application was the sub-plot. The means were compared using the least significant difference test (LSD) at 5% level (Snedecor and Cochran, 1980).
RESULTS AND DISCUSSION
Growth Characters and Yield Components
Concerning the effect of salinity (Table 2, 3, 5 and 6) with increasing levels there was a significant and gradual decreases in all of the studied growth characters and yield components in the two successive seasons of pot experiment. These results were in agree with that reported by Hu and Schmidhalter (1997), Hanafy Ahmed et al. (2002b) and Iqbal (2005) working on wheat. However, with soil salinity level 2000 ppm a reverse trend was recorded on leaf number and shoot dry weight in both seasons as well as on number of grains/plant in the first season of pot experiment.
The reduction in plant growth and yield due to salinity might be attributed to the inhibiting effects of salinity on many metabolic processes including protein, nucleic acid and polyamine synthesis (Mittle and Dubey, 1991; Reggiani et al., 1994), activity of mitochondria and chloroplasts (Udovenko and Tsibkovaskayai, 1983; Singh and Dubey, 1995), decreasing transpiration, stomatal conductance and photosynthesis (Sharma, 1995), restricts the absorption of water by plant roots and water use efficiency (Mansour, 1994), the toxic effects of certain ions present in soil solution (Maas and Nieman, 1978) and/or imbalance in phytohormone levels through its effect on either the biosynthesis or the destruction of the plant hormones (Nesiem and Ghallab, 1999).
| Table 2: | Shoot height (cm), leaf area (cm2) and leaf number of wheat plants sprayed by different levels of silicon and boron grown under different levels of soil salinity in pot experiment during the two successive seasons 2001-2002 and 2002-2003 |
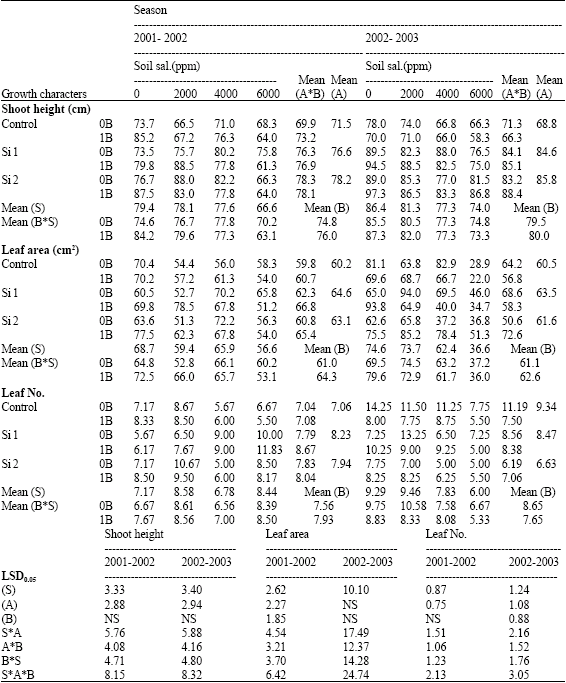 | |
| S: Soil salinity levels, A: Silicon treatment (Si 1 = 250 ppm SiO2 and Si 2 = 1000 ppm SiO2) and B: Boron treatment (0B = Tap water and 1B = 25 ppm boron), NS: Non Significant | |
The reduction in grain yield/plant at soil salinity levels 2000, 4000 and 6000 ppm reached 5.6, 30.3 and 42.9%, respectively when compared with control non-stressed plants in the first season. Meanwhile, the corresponding values in the second season were 17.7, 31.4 and 53.5%, respectively. In this respect, Hanafy Ahmed et al. (2002b) found in wheat a reduction on the most growth characters and yield components [number and/or weight of grains, weight of spikes as well as grain filling (weight of 1000 grain)] due to salinity.
| Table 3: | Number of tillers and shoot dry weight (g) of wheat plants sprayed by different levels of silicon and boron grown under different levels of soil salinity in pot experiment during two successive seasons (2001-2002 and 2002-2003) |
 | |
| S: Soil salinity levels, A: Silicon treatment (Si 1 = 250 ppm SiO2 and Si 2 = 1000 ppm SiO2), B: Boron treatment (0B = Tap water and 1B = 25 ppm boron) NS: Non Significant | |
Concerning the effect of silicon, in pots experiments, data presented in Table 2 and 3 revealed that, both levels of silicon either alone or combined with boron under both non-saline and saline soil conditions significantly increased shoots height and leaf area. However, no constant trend could be detected on leaf number, number of tillers and shoot dry weight, where they significantly increased in the first season at the lower level of silicon, while a reverse trend was recorded in the second season. These findings are in contrast with those previously stated by Hanafy Ahmed et al. (2002b) on wheat plants. They reported that, the highest rate of silicon application (1000 ppm SiO2) resulted in pronounced increase on all of the studied growth characters and all yield components of wheat, especially at the highest level of soil salinity (6000 ppm), but not at the lower rate of silicon (500 ppm SiO2).
The lowest level of silicon (250 ppm) significantly increased all the studied growth characters when compared with control non-sprayed plants (Table 4). Similar results were recorded by Trivedi et al. (2004).
| Table 4: | Growth characters and yield components of wheat plants sprayed by different levels of silicon and boron grown under saline field (4400 ppm) |
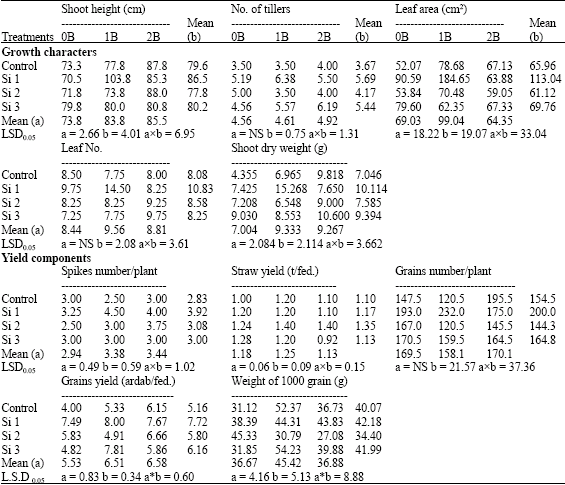 | |
| b: Silicon treatment, Si 1 = 250 ppm SiO2, Si 2 = 500 ppm SiO2, Si 3 = 1000 ppm SiO2, (a) Boron treatment, 0B = Tap water, 1B = 25 ppm boron, 2B = 50 ppm boron, NS: Non Significant | |
It could be suggested that, silicon application might exert their favorable effect to counteract the detrimental effects of salinity when the plants would show obvious stunting. In this connection, evidence has been provided that silicon not only contributes to cell wall rigidity and strengthening but might also increase cell wall elasticity during extension growth (Marschner, 1995). Moreover, the enhancement effect on shoot height of wheat plants supplied with Si might be induced through its role in both cell division and cell expansion by their effect on RNA and DNA synthesis. In this respect, Aleshin (1988) mentioned that nucleic acid preparations from rice tissues contain large amounts of Si. Moreover, silicon increased the thickness of the culm wall and the size of the vascular bundles (Shimoyama, 1958) preventing lodging in rice, thereby enhancing the strength of the stems.
Moreover, Ma and Takahashi (2002) and Ma (2004) reported that, Si could alleviate the effects of abiotic stresses including salt stress, metal toxicity and nutrient imbalance. These beneficial effects are mostly expressed through Si deposition in the leaves, stems and hulls. Therefore, Si is characterized by wide effects associated with the greater Si accumulation in the shoots.
Hanafy Ahmed et al. (2002b) and Trivedi et al. (2004) mentioned that, adding silicon under both non-saline and saline conditions reversing the salt inhibition on growth mainly by reducing the sodium uptake into the shoot. Moreover, Liang et al. (1996) concluded that silicon could increase photosynthesis and decrease the permeability of plasma membranes of leaves of salt stressed barley. They also mentioned that silicon could inhibit the uptake of Na and increase the uptake of K, thus mitigating salt toxicity to the plants and improving the vegetative growth of salt stressed
barley. Moreover, Marschner (1995) noted that, silicon could stimulate growth and yield under both non-saline and saline soil by several indirect actions. These include decreasing mutual shading through improving leaf erectness. Ma and Takahashi (1993) mentioned that the maintenance of erect leaves as a result of silicate application can easy account for 10% increase in the photosynthesis of the canopy and consequently increase in both growth and yield.
In addition, Agrie et al. (1992) working on rice, reported that silicon reduced transpiration and increased water use efficiency in leaves, which in turn reduced the decline in photosynthesis and chlorophyll destruction in older leaves. Also, Liang et al. (1996) pointed out that silicon treatment increased CO2 assimilation of barley leaves.
Concerning the effect of silicon on yield components of pot experiments the two successive seasons, silicon decreased spikes number/plant, straw yield and grains number/plant when compared with control non-sprayed plants (Table 5 and 6). While, grains yield/plant and weight of 1000 grain were significantly increased by both silicon treatments either alone or combined with boron under saline soil conditions in the two successive seasons, with some exceptions, but under non-saline soil the lower level of silicon combined with boron have the superiority effect comparing with control non-sprayed plants under the same saline soil condition. Concerning the effect of silicon on yield components under field conditions, all measured yield components significantly increased by all levels of silicon with some exceptions, as compared with control non-sprayed plants (Table 4). However, the lowest level of silicon had the superiority effect in all yield components, except of straw yield which was higher by the middle level of silicon.
In this respect, Pandey and Yadav (1999) reported that spraying silicon (50 or 100 ppm Si) increased grain yield/plant of wheat. They referred that to an increase in plant water status, chlorophyll content, biological yield and harvest index, coupled with reduced values of water potential, increase in dry matter accumulation, dry matter production rate, leaf area/plant at the flowering stage, productive tillers, grains and grain yield/main spike and per plant and transpiration rate coupled with a decrease in stomatal conductance. Moreover, Ma et al. (1989) working on rice mentioned that during the reproductive stage, silicon is preferentially transported into the flag leaf and interruption of silicon supply at this stage is detrimental for spikelet fertility.
Concerning the effects of boron foliar application treatment, in pots experiments, the two successive seasons, boron increased most of the studied growth characters, but these increases did not reach a significant level when compared with non-sprayed control plants with some exceptions (Table 2 and 3). Moreover, the shoot height and dry weight was significantly increased by boron application (Table 4). Similar results were recorded by Gunes et al. (2003) and Mete et al. (2005) on wheat. Meanwhile, number of leaves, leaves area and number or tillers did not affected with both boron levels, except the lower level of boron which significantly increased the number or tillers as compared with control non-sprayed plants with some exceptions. Similar results were recorded by Grieve and Poss (2000) on wheat. In this respect, several authors noted that, it`s well known that under salinity conditions boron concentration in the plants was decreased (Holloway and Alston, 1992; Wimmer et al., 2001). Moreover, changes in wall properties that resulted from decreased borate cross-linking of pectin might lead to many of the symptoms associated with boron deficiency, which almost associated with salinity in plants (O'Neill et al., 2005).
Concerning the effects of boron on yield components of pot experiments, the two successive seasons all measured yield components significantly increased by boron foliar application when compared with control non-sprayed plants, except of number of spikes/plant and weights of straw/plant which did not significantly affected. Similar results were recorded by Gunes et al. (2003) and Mete et al. (2005) on wheat. Also the interaction between boron and silicon take the same trend especially under non-saline and at 6000 ppm soil salinity conditions (Table 5 and 6).
| Table 5: | Spikes number, straw yield (g)/plant and grains number of wheat plants sprayed by different levels of silicon and boron grown under different levels of soil salinity in pot experiment during two successive seasons (2001-2002 and 2002-2003) |
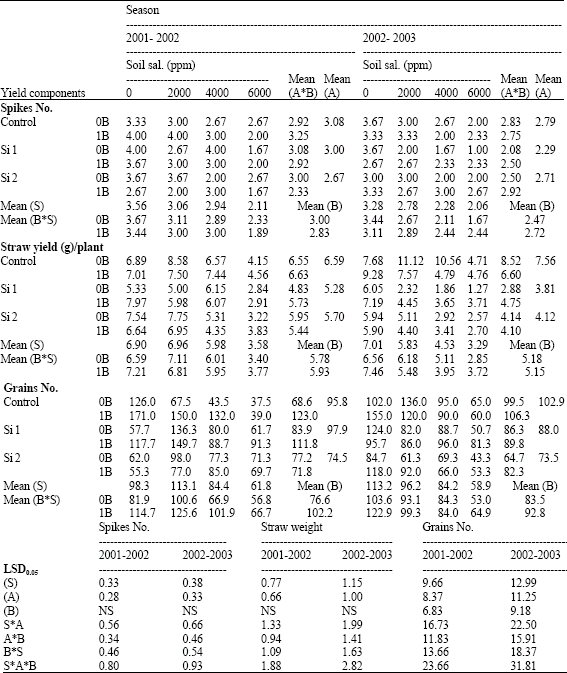 | |
| S: Soil salinity levels, A: Silicon treatment (Si 1 = 250 ppm SiO2 and Si 2 = 1000 ppm SiO2), B: Boron treatment (0B = Tap water and 1B = 25 ppm boron), NS: Non Significant | |
On the other hand, boron at both levels increased spikes number/plant and grains yield when compared with control non-sprayed plants (Table 4). Meanwhile, straw yield and weight of 1000 grains were significantly increased at the lowest level of boron. However, no significance differences were obtained from grains number. In this respect, Subedi et al. (1997) noted that, the soil addition of boron (1 kg B ha-1) significantly increased the number of grains/spike thereby decreasing the total sterility on wheat. Rahman et al. (2005) noted that, boron availability was enhanced by liming which could have significant effects in preventing spike-sterility and thereby increased yield of wheat and rice plants.
| Table 6: | Grains yield (g)/plant and weight of 1000 grain (g)of wheat plants sprayed by different levels of silicon and boron grown under different levels of soil salinity in pot experiment during two successive seasons (2001-2002 and 2002-2003) |
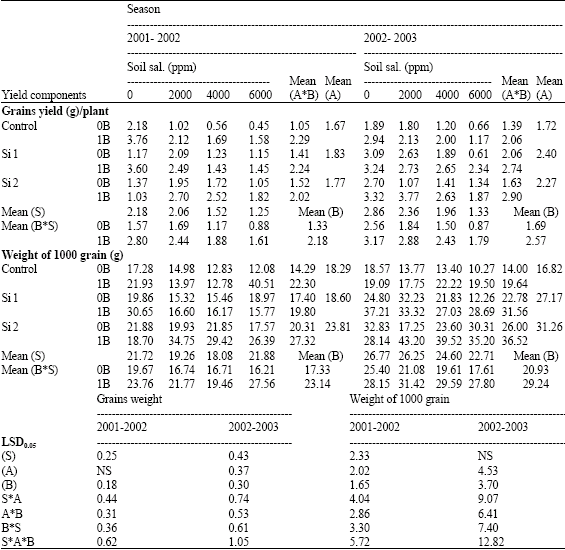 | |
| S: Soil salinity levels, A: Silicon treatment (Si 1 = 250 ppm SiO2 and Si 2 = 1000 ppm SiO2), B: Boron treatment (0B = Tap water and 1B = 25 ppm boron), NS: Non Significant | |
Chemical Composition
Inorganic Components
Concerning the effect of salinity on nutrients concentration, the low values of both N and K concentrations were recorded by shoots growing under salinity when compared with control plants grown under non-saline soil (Table 7). The reduction was more pronounced at the higher salinity level. In grains also a gradual reduction in N concentrations were recorded with increasing salinity level. These results were agree with those reported by Hu and Schmidhalter (1997) and Hanafy Ahmed et al. (2002b) on wheat. It is important here to mention that, salinity induced by using chloride or sodium salts might be implicated indirectly in decreasing N and K concentrations of wheat plants. In this respect, Hanafy Ahmed et al. (1996) mentioned that chloride seems to cause a decrease in nitrate accumulation, since its uptake is antagonistic to nitrate. Leidi et al. (1991) observed the competition between K+ and Na+ in wheat, especially by roots at different rates of salinity. They pointed out that a reduction of transpiration recorded at the highest salinity level could be correlated with a severe reduction in K level.
| Table 7: | Nitrogen, phosphorus and potassium concentrations (mg g-1 D.W) in the shoots of wheat plants sprayed by different levels of silicon and boron grown under different levels of soil salinity in pot experiment during two successive seasons (2001-2002 and 2002-2003) |
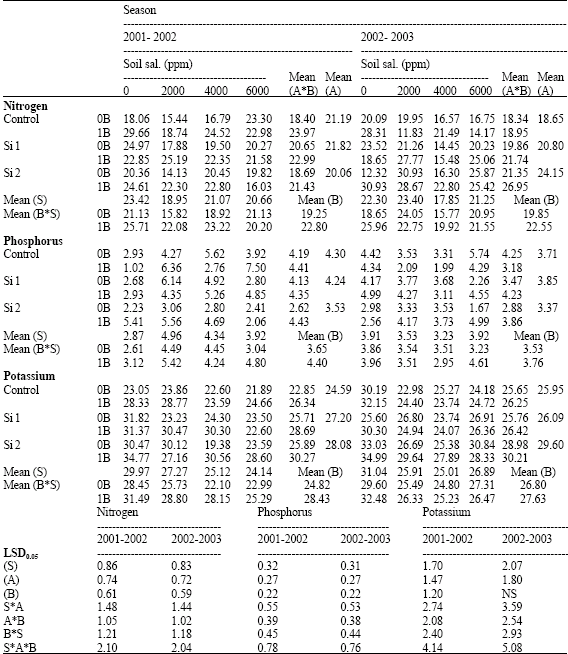 | |
| S: Soil salinity levels, A: Silicon treatment (Si 1 = 250 ppm SiO2 and Si 2 = 1000 ppm SiO2), B: Boron treatment (0B = Tap water and 1B = 25 ppm boron), NS Non Significant | |
In addition, higher values of Na, Ca and Mg concentrations were detected in saline stressed plants, which were more pronounced at higher than at lower salinity level (Table 8). This higher internal Na, Ca and Mg concentrations might be risen as a function of external addition of NaCl, CaCl2 and MgSO4 in the soil. However, no constant trend could be detected for P concentration at both seasons.
In this connection, Hanafy Ahmed et al. (2002b) suggested that the several detrimental effects attributed to salinity stress on growth characters and yield of wheat might be partially due to decreases in N and K concentrations and/or increases in P, Na, Ca, Mg values. Leidi et al. (1991) working on wheat reported that growth, leaf expansion and yield under saline conditions were limited by lowering intake of N and/or K by the plant in the presence of NaCl.
| Table 8: | Sodium, calcium and magnesium concentrations (mg g-1 D.W) in the shoots of wheat plants sprayed by different levels of silicon and boron grown under different levels of soil salinity in pot experiment during two successive seasons (2001-2002 and 2002-2003) |
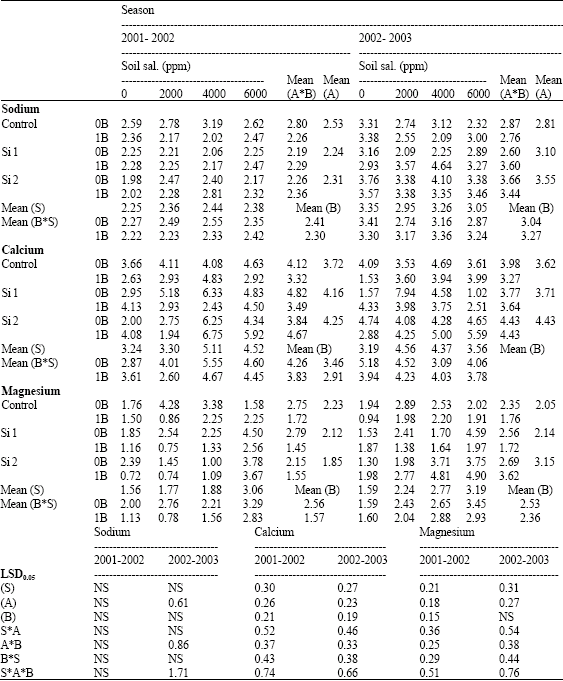 | |
| S: Soil salinity levels, A: Silicon treatment (Si 1 = 250 ppm SiO2 and Si 2 = 1000 ppm SiO2), B: Boron treatment (0B = Tap water and 1B = 25 ppm boron), NS: Non Significant | |
Moreover, high Na concentration may negatively affect plant growth due to the decrease in the absorption and translocation of water in these plants (Bernstein and Pearson, 1956), disturbing the fine structure of the plant cell causing swelling of the chloroplasts and plastids which may decreasing the net photosynthesis (Yeo et al., 1985) and affect the availability of most macronutrients in the plant (Hu and Schmidhalter, 1997). In addition, Hu and Schmidhalter (1997) reported that relative growth rate and yield were positively correlated with potassium content and negatively correlated with sodium content in root and shoot of wheat plants.
In pot experiments, the two successive seasons, high values of N, P, K and Ca concentrations were recorded by the shoots and grains with silicon application either alone or combined with boron comparing with control silicon non-sprayed plants, especially in shoots sprayed with boron, with some exceptions (Table 7-10). On the other hand, a reverse trend was recorded on Na concentration in grains. However, no constant trend could be detected for Na concentration in shoot and for Mg concentration in both shoots and grains by spraying silicon either alone or combined with boron when compared with control non-sprayed plants.
In this respect, Chen et al. (2002) reported that, absorption of N, P and K by rice plants was enhanced by silicon application. Moreover, Liang et al. (1996) and Hanafy Ahmed et al. (2002b) reported that, under saline soil conditions, low values of both Ca and Mg were recorded by the plants treated with 500 ppm SiO2, while a reverse trend was detected by spraying of 1000 ppm SiO2. Moreover, Hanan (1996) working on rice, suggested that the beneficial effects of Si might be ascribed to the isomorphus replacement of the phosphate ions with the silicate ions. However, Marschner (1995) mentioned that Si had no direct effect on P uptake or translocation to the roots. Liang et al. (1996) pointed out that Si could enhance the uptake of K and inhibit the uptake of Na in salt-stressed barley. In the same concern, Trivedi et al. (2004) reported that addition of silicon to the nutrient medium reversed the salt inhibited growth mainly by reducing the shoot Na uptake.
In field experiment, silicon was significantly increased N and K concentrations in shoots as well as N, P and Ca concentrations in grains, especially in the plants treated with the higher rate of silicon (Table 11). In this respect, Hanan (1996), Liang et al. (1996) and Hanafy Ahmed et al. (2002b) pointed out that N and K uptake was significantly increased in shoots when Si was added to soil. The stimulating effect of Si on N content might be due to the pH rise by sodium meta silicate which stimulates ammonification (Kai, 1978). Moreover, Nayar et al. (1982) reported that the application of silicate materials might help further increasing in grain yields, especially through efficient use of applied N. So, the data indicated that, the concentration of P increased in shoots whereas concentration of Na and K in grains were increased but not reached to significance, with some exceptions. However, Na and Mg concentrations decreased significantly in shoots as well as concentration of Mg in grains, with some exceptions, when compared with control silicon non-sprayed plants.
In this respect, it can be suggested that the decrease in Na might be induced by dilution effect as a result of beneficial effects of Si application on dry matter accumulation. Similar suggestion was recorded by Hanafy Ahmed et al. (2002b) on wheat plants. Moreover, Ma and Takahashi (1993) working on rice, suggested that Ca uptake and translocation are closely related to the transpiration. They also mentioned that transpiration rate was significantly decreased by the Si addition and caused the decreased on Ca uptake.
Thus, concerning the effect of silicon on alleviating the harmful effects of salinity, it can be suggested that, Si addition may improve nutritional balance under saline soil conditions, thereby a better growth performance and consequence yield production obtained. In this respect, Gharib and Hanafy Ahmed (2005) noted that, silicon foliar applications enhanced N, K and Na concentrations in pea shoots. It might be suggested that, B could improve the nutritional balance under saline soil conditions leading to better performance of silicon.
In addition, in field experiment, boron significantly increased P and K concentrations in shoots, while N was significantly decreased at the highest level (50 ppm) (Table 11). Meanwhile, Mg and Ca concentrations in shoots decreased significantly by the lowest level of boron (25 ppm) but increased significantly at the highest level of boron. While, no significantly differences could be detected on Na concentrations under both levels of boron. Moreover, the highest level of boron was significantly increased N whereare significantly decreased P and Ca in grains. While, no significant trend could be reported at the lowest level of boron on all of the studied minerals concentrations.
| Table 9: | Nitrogen, phosphorus and potassium concentrations (mg g-1 D.W) in grains of wheat plants sprayed by different levels of silicon and boron grown under different levels of soil salinity in pot experiment during two successive seasons (2001-2002 and 2002-2003) |
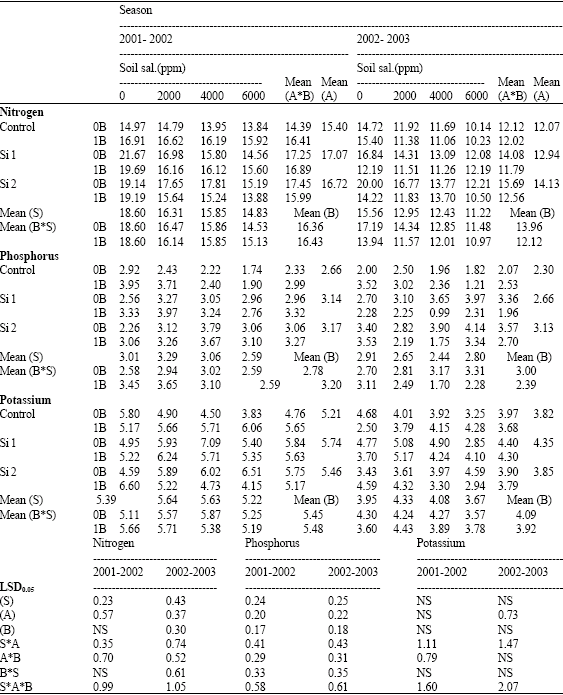 | |
| S: Soil salinity levels, A: Silicon treatment (Si 1 = 250 ppm SiO2 and Si 2 = 1000 ppm SiO2), B: Boron treatment (0B = Tap water and 1B = 25 ppm boron), NS: Non Significant | |
In this respect, Schon et al. (1990) noted that B deficiency induced inhibition of H+-ATPase activity causes a decrease in cation uptake, such as K+. Moreover, El-Magid et al. (2000) reported that, spraying wheat with B as boric acid at 0.06% reduced N content. Meanwhile, Shaaban et al. (2004) reported on wheat that, the highest uptake of K and Ca was obtained with foliar application of B (25 or 50 ppm), which suggested that boron stimulates metabolic functions facilitate absorption and translocation of these nutrients, especially in plants grown under lime stress conditions.
| Table 10: | Sodium, calcium and magnesium concentrations (mg g-1 D.W) in grains of wheat plants sprayed by different levels of silicon and boron grown under different levels of soil salinity in pot experiment during two successive seasons (2001-2002 and 2002-2003) |
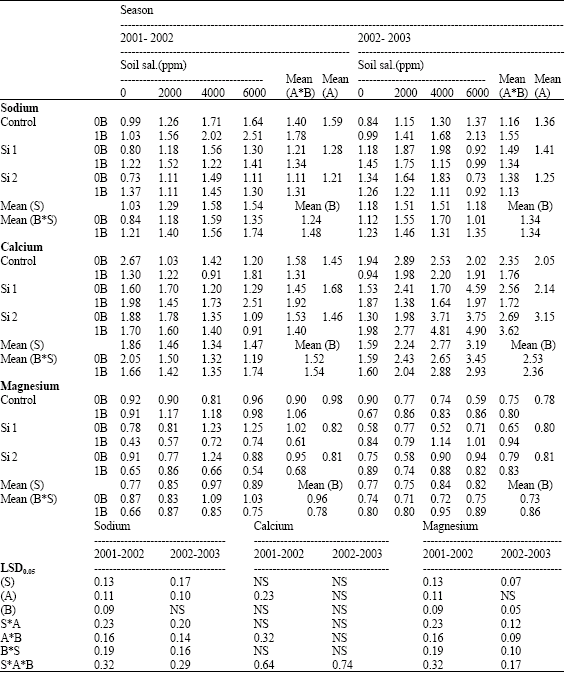 | |
| S: Soil salinity levels, A: Silicon treatment (Si 1 = 250 ppm SiO2 and Si 2 = 1000 ppm SiO2), B: Boron treatment (0B = Tap water and 1B = 25 ppm boron), NS: Non Significant | |
Also, Lopez-Lefebre et al. (2002) noted that, increasing B application (5, 10 and 20 mmol L-1 H3BO3) induced a general improvement in the nutritional state, particularly of the essential macronutrients N and P in tobacco plants (Nicotiana tabacum L.) which could explain the positive effects of B on increase dry-material production in the roots and leaves.
Organic Components
Concerning the effects of salinity on studied organic components of wheat plants, there is a gradual reduction on total chlorophyll of the two successive seasons by increasing salinity level when compared with control plants grown under non-saline soil (Table 12). In this respect, El-Bagoury et al. (1999) suggested that, biosynthesis of chlorophylls in generally might be inhibited by the depressive effect of stress conditions on the absorption of some ions involved in the chloroplast formation, such as Mg and Fe and/or an increase of growth inhibitors, such as ethylene or abscisic acid production which enhance senescence. Salt could induce weakening of protein-pigment-lipid complex thus decreasing chlorophyll concentration (Strogonove et al., 1970) or increased chlorophyllase activity (Stivesev et al., 1973).
| Table 11: | Nitrogen, phosphorus, potassium, sodium, calcium and magnesium concentrations (mg g-1 D.W.) in shoots as well as grains of wheat plants sprayed by different levels of silicon and boron grown under saline field (4400 ppm) experiment |
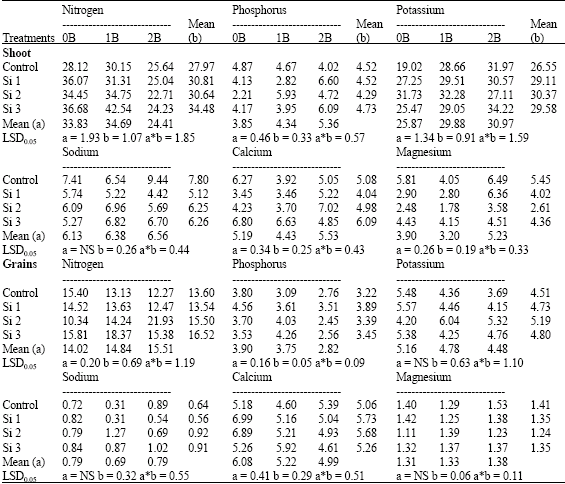 | |
| b: Silicon treatment, Si 1 = 250 ppm SiO2, Si 2 = 500 ppm SiO2, Si 3 = 1000 ppm SiO2, (a) Boron treatment, 0B = Tap water, 1B = 25 ppm boron, 2B = 50 ppm boron, NS: Non Significant | |
In addition, the higher values of total sugars, free amino acids, soluble phenols and proline concentrations were detected in saline-stressed plants and these increases were more pronounced at the highest salinity level (Table 12 and 13).
In this respect, Hanafy Ahmed et al. (2002b) working on wheat mentioned that salinized plants accumulated soluble carbohydrate and showed a remarkable higher levels of amino acids, soluble phenols and proline for osmoregulation. This might be explained on the assumption that such salt-stressed plants could have less efficiency to condensate simple organic compounds into more complex ones. Margna (1977) reported that the major determinant of phenol production is the supply of the prerequisites for its synthesis, namely the amino acid precursor`s phenylalanine and tyrosine.Under pot experiment, in the second season, the total free-polyamines concentrations gradually increased with increasing salinity level then decreased under the highest salinity level (6000 ppm), but, still higher than control non-saline plants (Table 14). These results are in agreement with those obtained by Santa-Cruz et al. (1997) on tomato and Hanafy Ahmed et al. (2002a) on Myrtus communis plants. Katiyar and Dubey (1990) working on rice, indicated that NaCl stress induced accumulation of Polyamines (PAs) which possibly might helping the cells in maintaining more favorable ionic environment and in regulating cellular pH.
| Table 12: | Effect of silicon and boron spraying at different levels on concentrations of total chlorophyll in leaves (mg g-1 F.W.) as well as total sugars (mg glucose/g F.W.) and total free amino acids (mg g-1 F.W.) in shoots of wheat plants grown under different soil salinity levels in pot experiment during two successive seasons (2001-2002 and 2002-2003) |
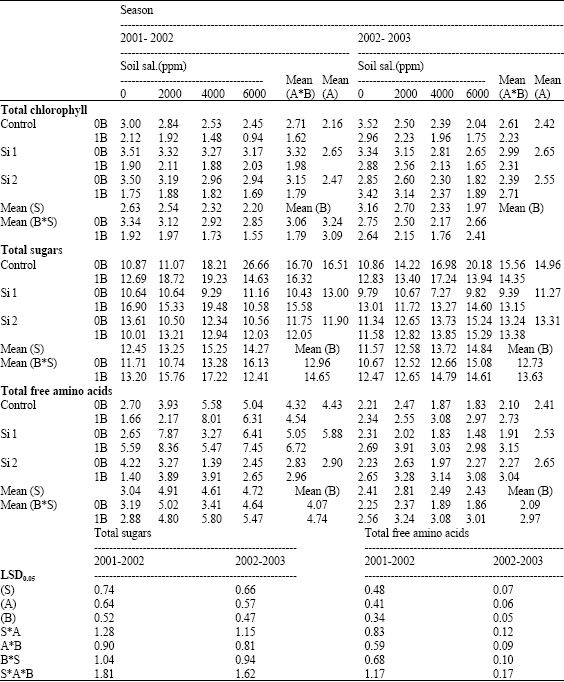 | |
| S: Soil salinity levels, A: Silicon treatment (Si 1 = 250 ppm SiO2 and Si 2 = 1000 ppm SiO2), B: Boron treatment (0B = Tap water and 1B = 25 ppm boron), NS: Non Significant | |
Recently, Legocka and Kluk (2005) reported that salinity stress (NaCl 260mM) induced biosynthesis of Putrescine (Put) and other PAs in the roots, as well as induced Put accumulation in the leaves, which might indicate for translocation of Put from the roots to the shoot in lupin (Lupinus luteus L. var. Ventus). In this connection, Willadino et al. (1996) pointed out that Put is synthesized in plants from arginine and/or ornithine by the action of the biosynthetic enzymes Arginine Decarboxylase (ADC) and Ornithine Decarboxylase (ODC). ADC activity showed significant increments in relation to salt stress, which may be related to Put and some amino acid variations. Several studies have demonstrated that the accumulated Put in stressed tissues is derived from L-arginine and is due largely to increased activities of the ADC pathway. Krishnamurthy and Bhagwat (1989) pointed out that the Put accumulation observed in rice in response to salt stress might have resulted from an enhancement of Put synthesis, alternatively, from an inhibition of spermidin and spermin synthesis through the inhibition of the activity of the enzyme S-adenosylmethionine. Starvation of K might accumulate Put in leaves to replace K+ as a result of unbalanced uptake of ions under saline conditions to reflect a homeostatic mechanism for controlling cellular pH in highest plants (Watson et al., 1998).
| Table 13: | Effect of silicon and boron spraying at different levels on concentrations of total soluble phenol (mg g-1 F.W.) and proline (mg g-1 D.W.) in shoots of wheat plants grown under different soil salinity levels in pot experiment during two successive seasons (2001-2002 and 2002-2003) |
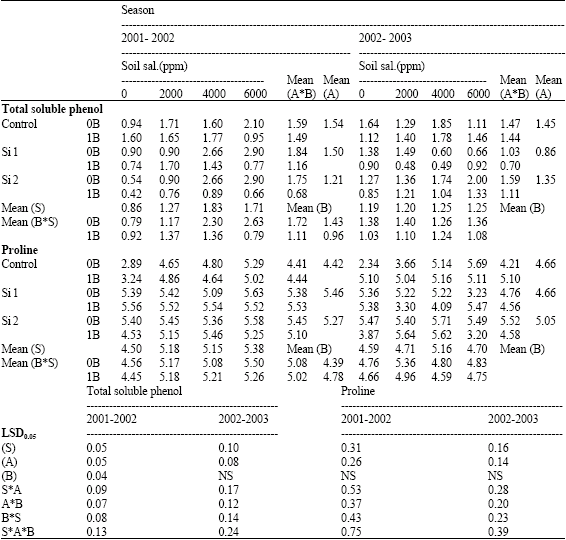 | |
| S: Soil salinity levels, A: Silicon treatment (Si 1 = 250 ppm SiO2 and Si 2 = 1000 ppm SiO2), B: Boron treatment (0B = Tap water and 1B = 25 ppm boron), NS: Non Significant | |
Concerning the effect of silicon on organic components, the pot experiment of both seasons, total chlorophyll concentrations increased by silicon foliar application, especially at the lowest level of silicon under both non-saline and saline soil conditions (Table 12, 15). Meanwhile, under field experiment chlorophyll concentration increased at the higher levels of silicon, e.g., 500 and 1000 ppm. Similar results were reported by Trivedi et al. (2004) on wheat.
| Table 14: | Total free polyamine concentration (mmol Put/g F.W.) in shoots of wheat plants sprayed by different levels of silicon and boron grown under different soil salinity levels in pot experiment during season 2002-2003 |
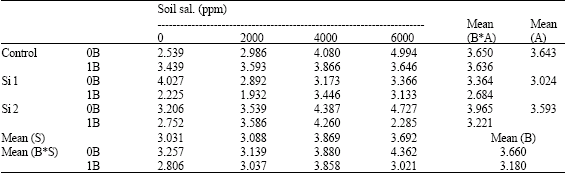 | |
| S: Soil salinity levels, A: Silicon treatment (Si 1 = 250 ppm SiO2 and Si 2 = 1000 ppm SiO2), B: Boron treatment (0B = Tap water and 1B = 25 ppm boron) NS: Non Significant | |
| Table 15: | Effect of silicon and boron spraying at different levels on concentrations of total chlorophyll in leaves (mg g-1 F.W.) as well as proline (mg g-1 D.W.), total free polyamine (m mol Put/g F.W.), total sugars (mg glucose/g F.W.), total free amino acids (mg g-1 F.W.) and total soluble phenol (mg g-1 F.W.) in shoots of wheat plants grown under saline filed (4400 ppm) experiment |
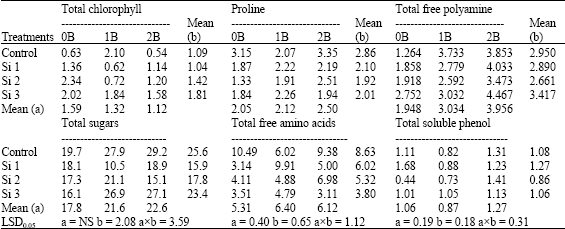 | |
| b: Silicon treatment, Si 1 = 250 ppm SiO2, Si 2 = 500 ppm SiO2, Si 3 = 1000 ppm SiO2, 2B = 50 ppm boron, NS: Non Significant | |
In these respect, Ma et al. (2004) reported that, under drought stress silicon decreased the decomposition of chlorophyll in cucumber plants, limited plasma membrane permeability, alleviated malondialdehyde content in leaves, as well as the activities of peroxidase, superoxide dismutase and catalase. These physiological biochemical reactions showed positive correlations with the amount of silicon supply.
Total chlorophyll concentrations decreased by boron foliar application either alone or combined with silicon under both non-saline and saline soil conditions when compared with control boron non-sprayed plants. Also under field experiment, revealed that, chlorophyll decreased by increasing boron concentration. Similar results were reported by Hanafy Ahmed et al. (1996) on faba bean.
Concerning total sugars concentration, both pot experiments, silicon either alone or combined with boron under saline soil conditions significantly decreased total sugars concentration. However, under non-saline soil conditions a reverse trend was obtained with some exceptions. Meanwhile, total soluble phenols concentration was decreased in shoots of both seasons when compared with control non-sprayed plants under both saline and non-saline soil conditions. In this respect, Hanafy Ahmed et al. (2002b) suggested that silicon could inhibit the synthesis of total soluble phenols under saline soil conditions. Moreover, Parry and Kelso (1975) reported that, Si seems to influence the content and metabolism of polyphenols in xylem cell walls.
In addition, in both pot experiments, silicon either alone or combined with boron significantly increased total free amino acids and proline concentrations in shoots under both non-saline and saline soil conditions when compared with control non-sprayed plants, with some exceptions. In field experiment, silicon significantly decreased total sugars, total free amino acids, total soluble phenols and proline concentrations in shoots when compared with control non-sprayed plants. In this connection, Hanafy Ahmed et al. (2002b) noted that, the favorable effects of silicon on alleviating the harmful effects of salinity on wheat plants could be indirectly through increase the ability of plants under salinity stress to condensate simple organic compounds, e.g., sugars, free amino acids, soluble phenols into more complex ones, e.g., carbohydrates, protein, polyphenols. They added that, silicon might be an adaptive response to salinity, through raising proline concentration. In addition, Schmidt et al. (1999) noted that silicon might positively affect the activity of some enzymes involved in the photosynthesis of turfgrass Regarding the effect of boron, total sugars and total free amino acids concentrations significantly increased in shoots by boron foliar application in both pot experiments especially under high salinity levels. Also, total sugars, total free amino acids and proline concentrations increased under field experiment when compared with control boron non-sprayed plants. However, proline concentration in shoots of both pot experiments did not affected by boron spraying. On the other hand, total soluble phenols concentration in shoots was significantly decreased in the first season of pot experiment as well as under field conditions at the lower level of boron (25 ppm) (Table 12, 13 and 15).
In this connection, Mengel and Kirkby (1979) pointed out that B might modify the activity of the enzyme 6-phosphogluconate dehydrogenase by complexing with its substrate 6-phosphogluconate. The enzyme regulates the first step in the pentose phosphate pathway. It is therefore argued that when B is present the activity of the pentose phosphate pathway is decreased in favor of glycolysis. On the other hand, when boron is deficient the pentose phosphate pathway is favored and consequently induces the accumulation of phenolic compounds. They added that B and Cu may have possible effects on DNA and RNA synthesis, which could increase the rate of protein synthesis.
In this respect, Eweida et al. (1984) reported that spraying sugar beet plants with boron at 100 ppm gave the highest sugar concentration. Moreover, Acerbo et al. (1973) and Dugger (1983) mentioned that the conversion of soluble sugars or sugar phosphate to starch was inhibited by boron deficiency in plants.
Concerning the effect of silicon and/or boron on concentration of total free-polyamines, under pot experiment conditions, all studied treatments reduced total free-polyamines when compared with control non-sprayed plants especially under saline soil conditions with some exceptions (Table 14). In this connection, Watson et al. (1998) reported that, boron deficiency induced free putrescine accumulation in Arabidopsis due to the differential and unbalanced uptake of ions and cell pH. So, it may be suggested that boron could rebalance the uptake of ions resulting in rebalance in cell pH, leading to decrease total free-polyamines level under different salinity levels as compared with control non-sprayed plants. Moreover, Camacho-Cristóbal et al. (2005) working on tobacco plants reported that, levels of free Put and Spd in leaves were similar in both boron-sufficient and boron-deficient plants. However, boron-deficient plants had higher conjugated polyamine concentration than boron-sufficient plants.
Under field experiment, both silicon levels, i.e., 250 and 500 ppm lowered total free-polyamines, while the highest level (1000 ppm SiO2) increased it when compared with control non-sprayed plants. On contrary, a reverse trend was recorded for the plants treated with boron. This means that polyamines increased with increasing boron concentration either alone or combined with silicon when compared with control non-sprayed plants.
In this connection, it is well known that polyamine metabolism is altered in response to an inadequate nutrition in K, Mg or P (Slocum et al., 1984). For example, free putrescine dramatically increases in K-deficient plants, suggesting that polyamines could play a role in maintaining cation-anion balance in plant tissues (Flores and Martin-Tanguy, 1991). In relation to the short-term effects caused by boron deficiency, Camacho-Cristóbal et al. (2002) have described an accumulation of caffeoylputrescine in leaves from boron-deficient tobacco plants, which lead to suggest a possible role of boron in polyamine metabolism.
Several detrimental effects attributed to salinity stress on most of the studied growth characters might be partially due to an increase in the total free polyamine concentrations. In this respect, Smith (1977) stated that salt stress caused a significant change in intracellular concentrations of polyamine. Moreover, Di Tomaso et al. (1989) pointed out that excess Put may have some negative effects on corn plant. Hsiao (1973) reported that Put rise appears to be more sensitive to turgor deficit, causing inhibition of cell growth, cell wall synthesis and ABA accumulation. Moreover, N'Doye et al. (1994) working on tomato, proposed that a high PA content in light might have a deleterious effects on the photosynthetic apparatus. Additionally, Strogonove (1973) noted that, Put feeding is the direct cause of the necrosis observed in salt-stressed plants. Thus, it appears that, the effect of silicon and boron foliar application on decreasing PAs accumulation might be indirectly implicated on reducing the hazard effects attributed to salinity stress on most of the studied growth characters.
Concerning plant hormones concentrations in the shoots of plants growing under pot experiment in the second season, gradual decreases were detected in IAA, GA3 and CK with increasing salinity level (Table 16). Meanwhile, an opposite trend was obtained with abscisic acid (ABA). These findings are in agreement with those obtained by El-Shafey et al. (2003) and Nesiem and Ghallab (1999) on wheat plants.
In this respect, Zeinab and Sallam (1996) reported that with increasing salinity level, the tryptophan synthase α-monomers were gradually dissociated from the oligomers producing less active soenzyme. This reduced the biosynthesis of L-tryptophan and consequently IAA. Also, CK activity decreased with increasing salt stress in barley plants. They added that, gibberellin inhibitors accumulated, so plant growth was retarded or even stopped.
| Table 16: | IAA, GA, ABA and CK concentrations (μg g-1 F.W.) in shoots of wheat plants sprayed by different levels of silicon and boron grown under different soil salinity levels in pot experiment during season 2002-2003 |
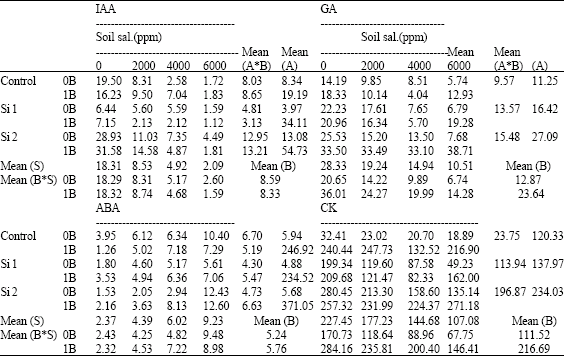 | |
| S: Soil salinity levels, A: Silicon treatment (Si 1 = 250 ppm SiO2 and Si 2 = 1000 ppm SiO2), B: Boron treatment (0B = Tap water and 1B = 25 ppm boron), NS: Non Significant | |
Silicon either alone or combined with boron under different salinity levels increased GA3 and CK concentrations. The boron-silicon combinations have the superiority effect. However, no constant trend could be detected from spraying silicon on IAA. IAA decreased by the lowest level of silicon either alone or combined with boron, while a reverse trend was detected at the highest level of silicon either alone or combined with boron. On the other hand, ABA concentration decreased by both levels of silicon either alone or combined with boron when compared with control non-sprayed plants, with some exceptions under high salinity levels. Silicon sprayed alone has the superiority effect on decreasing ABA than silicon combined with boron.
It could suggest that, reducing the transpirational loss of water as a beneficial effect of silicon under saline soil conditions might increase the cell turgor, which could be more helpfully in rebalancing phytohormones levels through its effect on either the biosynthesis (Amzallage et al., 1992) or its destruction. The enhancement of the phytohormone levels in treated plants within a suitable concentration range might improve plant growth and yield of wheat plants. Al-aghabary et al. (2004) studied the influence of silicon (2.5 mM), sodium chloride (NaCl 100 mM) and Si (2.5 mM) + NaCl (97.5 mM) supply on H2O2 level and activities of some enzymes related to antioxidation protection against the potentially cytotoxic forms of activated oxygen under salinity. Silicon partially offset the negative impacts of NaCl stress by raising superoxide dismutase and catalase activities. Also, Si addition decreased ascorbate peroxidase activity and significantly reduced H2O2 level in the leaves of salt-treated plants (Rout and Show, 2001). It could be suggested that, silicon might alleviate the effects of salt stress by reducing peroxidase activity, which led to increase IAA by decreasing its degradation.
Concerning the effect of boron foliar application on plant hormones concentrations in shoots, data in Table 16 revealed that, boron increased GA3 and CK concentrations reaching about 10-fold either alone or combined with silicon as compared to control non-sprayed plants under both non-saline and saline soil conditions. Moreover, boron either alone or combined with silicon decreased ABA under both non-saline and saline soil conditions, except of a slight increases were found at 4000 or 6000 ppm soil salinity. However, no constant trend could be detected for boron on IAA.
In this respect, Hanafy Ahmed et al. (1996) noted that, boron treatments (25, 50 and 75 ppm) increased CK and GAs, but free auxins were drastically reduced by all boron treatments on faba bean, while ABA did not affect except of an increase was recorded with 75 ppm B treatment. Moreover, Hanafy Ahmed et al. (1999) noted that, boron foliar application (0.0, 100, 250 and 500 ppm as boric acid) increased IAA and GAs in peas plants shoots, while ABA tended to decline.
In this connection, Wagner and Michael (1971) and Maynard and David (1987) reported that, B deficiency has been found to depress CK concentrations but an increase in IAA production was obtained as a result in the reduction of IAA oxidase activity. Moreover, Das (2000) noted that boron deficiency induced dramatic increase in the activity of IAA oxidase in the roots which falls rapidly when boron is supplied. Moreover, Srivastava and Gupta (1996) noted that boron could regulate endogenous auxin in plants by protecting the IAA oxidase system through complexation of o-diphenols; inhibitors of IAA oxidase. Excessive auxin activity found under B deficiency could cause excessive proliferation of cambial cells, rapid and disproportionate enlargement of cells and collapse of nearby cells.
It is well known that, in response to various environmental stresses, such as drought and salinity, stomata is close as a process that involves the phytohormone ABA (Schroeder et al., 2001). Tanaka et al. (2005) found that, CK and auxins inhibit the effects of ABA on induced stomatal closure. Also, CK decreased the decomposition of chlorophyll and photosynthesis. So, It could suggest that, the reducing of the transpirational loss of water as a beneficial role of silicon (Agrie et al., 1992), could kept the transpiration rate at a relatively steady rate during stress, leading to increase the cells turgor which might be more helpful in rebalancing of phytohormones by increasing IAA, GA and CK and decreasing ABA which was reflected on improving plant growth and consequently on yield of wheat plants.
CONCLUSION
In conclusion, the results of this study highlight the role of silicon in improving wheat yield under non-saline or saline soil conditions either alone or combined with boron. We suggest that under non-saline soil conditions silicon should be sprayed by concentration 250 ppm combined with 0.25 ppm boron at 40 and 70 days after sowing with under investigation study significantly increased grains yield/plant. Furthermore under different saline soil all levels of silicon (250 and 1000 ppm SiO2 under pots experiment as well as 250, 500 and 1000 ppm SiO2 under field experiment) either alone or combined with boron significantly increased grains yield/plant with some exceptions. However, the lowest level of silicon combined with boron had the superiority effect in all measured yield components under both pots and field experiments.
REFERENCES
- Acerbo, S., R. Kastori, H. Sochtig, H. Harms and K. Haider, 1973. Effect of boron on synthesis and transformation of lignin-precursors in Zea mays. Zeitschrift Pflanzenphysiologie, 69: 306-317.
CrossRefDirect Link - Agarie, S., W. Agata, F. Kubota and P.B. Kaufman, 1992. Physiological roles of silicon in photosynthesis and dry matter production in rice plants: I. Effects of silicon and shading treatments. Jpn. J. Crop Sci., 61: 200-206.
CrossRefDirect Link - Al-Aghabary, K., Z. Zhu and Q. Shi, 2005. Influence of silicon supply on chlorophyll content, chlorophyll fluorescence and antioxidative enzyme activities in tomato plants under salt stress. J. Plant Nutr., 27: 2101-2115.
CrossRefDirect Link - Amzallag, G.N., H.R. Lerner and A. Poljakoff-Mayber, 1992. Interaction between mineral nutrients, cytokinin and gibberellic acid during growth of sorghum at high NaCI salinity. J. Exp. Bot., 43: 81-87.
CrossRefDirect Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Bernstein, L. and G.A. Pearson, 1956. Influence of exchangeable sodium on the yield and chemical composition of plants: I. Green beans, garden beets, clover and alfalfa. Soil Sci., 82: 247-258.
Direct Link - Camacho-Cristobal, J.J., D. Anzellotti and A. Gonzalez-Fontes, 2002. Changes in phenolic metabolism of tobacco plants during short-term boron deficiency. Plant Physiol. Biochem., 40: 997-1002.
Direct Link - Camacho Cristobal, J.J., J.M. Maldonado and A. Gonzalez Fontes, 2005. Boron deficiency increases putrescine levels in tobacco plants. J. Plant Physiol., 162: 921-928.
Direct Link - Chen, J.H., G.J. Mao, G.P. Zhang and H.D. Guo, 2002. Effects of silicon on dry matter and nutrient accumulation and grain yield in hybrid Japonica rice (Oryza sativa L.). J. Zhejiang Univ. (Agric. Life Sci.), 28: 22-26.
Direct Link - Di Tomaso, J., J. Shaff and L. Kochian, 1989. Putrescine-induced wounding and its effects on membrane integrity and ion transport processes in roots of intact corn seedlings. Plant Physiol., 90: 988-995.
Direct Link - DuBois, M., K.A. Gilles, J.K. Hamilton, P.A. Rebers and F. Smith, 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem., 28: 350-356.
CrossRefDirect Link - El-Magid, A.A.A., R.E. Knany and H.G.A. El Fotoh, 2000. Effect of foliar application of some micronutrients on wheat yield and quality. Ann. Agric. Sci. Cairo, 1: 301-313.
Direct Link - Galston, A.W., 1983. Polyamines as modulators of plant development. BioScience, 33: 382-388.
CrossRefDirect Link - Grieve, C.M. and J.A. Poss, 2000. Wheat response to interactive effects of boron and salinity. J. Plant Nutr., 23: 1217-1226.
Direct Link - Gunes, A., M. Alpaslan, A. Inal, M.S. Adak, F. Eraslan and N. Cicek, 2003. Effects of boron fertilization on the yield and some yield components of bread and durum wheat. Turk. J. Agric. For., 27: 329-335.
Direct Link - Hanafy Ahmed, A.H., A.M. Farrag, M.R. Nesiem and M.K. Khalil, 1999. Effect of Rhizobium inoculation, nitrogen fertilization and boron on growth, yield and chemical composition of peas. Proceedings of the 1st Congress Recent Technology Agriculture Fac. Agriculture, November 12-14, 1999, Cairo University, pp: 202-217.
- Hanafy, A.H., M.A. Gad-Mervat, H.M. Hassam and A. Amin-Mona, 2002. Improving growth and chemical composition of Myrtus communis grown under soil salinity conditions by polyamines foliar application. Proceedings of the Minia 1st Conference Agriculture Environment Science Minia, March 25-28, 2002, Egypt, pp: 1697-1720.
- Holloway, R.E. and A.M. Alston, 1992. The effects of salt and boron on growth of wheat. Aust. J. Agric. Res., 43: 987-1001.
CrossRefDirect Link - Hsiao, T.C., 1973. Plant responses to water stress. Annu. Rev. Plant Physiol., 24: 519-570.
CrossRefDirect Link - Hu, Y. and U. Schmidhalter, 1997. Interactive effects of salinity and macronutrient level on wheat. II. Composition. J. Plant Nutr., 20: 1169-1182.
CrossRef - Hu, Y. and U. Schmidhalter, 2005. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J. Plant Nutr. Soil Sci., 168: 541-549.
CrossRefDirect Link - Iqbal, R.M., 2005. Effect of different salinity levels on partitioning of leaf area and dry matter in wheat. Asian J. Plant Sci., 4: 244-248.
CrossRefDirect Link - Jackson, M.L., 1973. Soil Chemical Analysis. 1st Edn., Prentice Hall Ltd., New Delhi, India, Pages: 498.
Direct Link - Katiyar, S. and R.S. Dubey, 1990. Changes in polyamine titer in rice seedlings following NaCl salinity stress. J. Agron. Crop Sci., 165: 19-27.
CrossRefDirect Link - Krishnamurthy, R. and K.A. Bhagwat, 1989. Polyamines as modulators of salt tolerance in rice cultivars. Plant Physiol., 91: 500-504.
CrossRefDirect Link - Legocka, J. and A. Kluk, 2005. Effect of salt and osmotic stress on changes in polyamine content and arginine decarboxylase activity in Lupinus luteus seedlings. J. Plant Physiol., 162: 662-668.
CrossRefPubMedDirect Link - Leidi, E.O., M. Silberbush and S.H. Lips, 1991. Wheat growth as affected by nitrogen type, pH and salinity. II. Photosynthesis and transpiration. J. Plant Nutr., 14: 247-256.
CrossRefDirect Link - Liang, Y., Q. Shen, Z. Shen and T. Ma, 1996. Effects of silicon on salinity tolerance of two barley cultivars. J. Plant Nutr., 19: 173-183.
CrossRefDirect Link - Lopez-Lefebre, L.R., R.M. Rivero, P.C. Garcia, E. Sanchez, J.M. Ruiz and L. Romero, 2002. Boron effect on mineral nutrients of tobacco. J. Plant Nutr., 25: 509-522.
CrossRefDirect Link - Ma, J.F., K. Nishimura and E. Takahashi, 1989. Effect of silicon on the growth of rice plant at different growth stages. Jap. J. Soil Sci. Plant Nutr., 35: 347-356.
CrossRefDirect Link - Ma, J.F. and E. Takahashi, 1993. Interaction between calcium and silicon in water-cultured rice plants. Plant Soil, 148: 107-113.
CrossRefDirect Link - Ma, C.C., Q.F. Li, Y.B. Gao and T.R. Xin, 2004. Effects of silicon application on drought resistance of cucumber plants. Soil Sci. Plant Nutr., 50: 623-632.
Direct Link - Ma, J.F., 2004. Role of silicon in enhancing the resistance of plants to biotic and abiotic stresses. Soil Sci. Plant Nutr., 50: 11-18.
CrossRefDirect Link - Mansour, M.M.F., 1994. Changes in growth, osmotic potential and cell permeability of wheat cultivars under salt stress. Biol. Plant., 36: 429-434.
CrossRefDirect Link - Margna, V., 1977. Control at the level of substrate supply: An alternative in the regulation of phenylpropanoid accumulation in plant cells. Phytochemistry, 16: 419-426.
CrossRefDirect Link - Moore, S. and W.H. Stein, 1954. A modified ninhydrin reagent for the photometric determination of amino acids and related compounds. J. Biol. Chem., 211: 907-913.
CrossRefDirect Link - N'Doye, M., B. Millet and M. Paynot, 1994. Diurnal modulation of polyamine content in seedlings of Lycopersicum esculentum Mill.cv. Ace 55 by temperature and light. Plant Sci., 104: 11-15.
CrossRefDirect Link - Moran, R., 1982. Formulae for determination of chlorophyllous pigments extracted with N,N-dimethylformamide. Plant Physiol., 69: 1376-1381.
CrossRefDirect Link - O'Neill, M.A., T. Ishii, P. Albersheim and A.G. Darvill, 2004. Rhamnogalacturonan II: Structure and function of a borate cross-linked cell wall pectic polysaccharide. Annu. Rev. Plant Biol., 55: 109-139.
CrossRefDirect Link - Parry, D.W. and M. Kelso, 1975. The distribution of silicon deposits in the roots of Molinia caerulea (L.) Moench. and Sorghum bicolor (L.) Moench. Ann. Bot. (London) (N.S.), 39: 995-1001.
CrossRefDirect Link - Rahman, M.A., J. Chikushi, J.M. Duxbury, C.A. Meisner, J.G. Lauen and E. Yasunaga, 2005. Chemical control of soil environment by lime and nutrients to improve the productivity of acidic alluvial soils under rice-wheat cropping system in Bangladesh. Environ. Control Biol., 43: 259-266.
Direct Link - Reggiani, R., S. Bozo and A. Bertani, 1994. Changes in polyamine metabolism in seedlings of three wheat (Triticum aestivum L.) cultivars differing in salt sensitivity. Plant Sci., 102: 121-126.
CrossRefDirect Link - Rout, N.P. and B.P. Show, 2001. Salt tolerance in aquatic macrophytes: Possible involvement of the antioxidative enzymes. Plant Sci., 160: 415-423.
CrossRefDirect Link - Sadeghian, E., 1971. EinfluB von Chlorcholinchlorid (CCC) auf den Gibberellingehalt (GA3) von Getreidepflanzen. Z Pflanzenern, Bodenk, 130: 233-241.
CrossRefDirect Link - Santa-Cruz, A., M.T. Estan, A. Rus, M.C. Bolarin and M. Acosta, 1997. Effects of NaCl and mannitol iso-osmotic stresses on the free polyamine levels in leaf discs of tomato species differing in salt tolerance. J. Plant Physiol., 151: 754-758.
CrossRefDirect Link - Schmidt, R.E., X. Zhang and D.R. Chalmers, 1999. Response of photosynthesis and superoxide dismutase to silica applied to creeping bentgrass grown under two fertility levels. J. Plant Nutr., 22: 1763-1773.
CrossRefDirect Link - Schon, M.K., A. Novacky and D.G. Blevins, 1990. Boron induces hyperpolarization of sunflower root cell membranes and increases membrane permeability to K+. Plant Physiol., 93: 566-571.
Direct Link - Schroeder, J.I., G.J. Allen, V. Hugouvieux, J.M. Kwak and D. Waner, 2001. Guard cell signal transduction. Ann. Rev. Plant Physiol. Plant Mol. Biol., 52: 627-658.
Direct Link - Shaaban, M.M., M.M. El-Fouly and A.W.A. Abdel-Maguid, 2004. Zinc-boron relationship in wheat plants grown under low or high levels of calcium carbonate in the soil. Pak. J. Biol. Sci., 7: 633-639.
CrossRefDirect Link - Slocum, R.D., R. Kaur-Sawhney and A.W. Galston, 1984. The physiology and biochemistry of polyamines in plants. Arch. Biochem. Biophys., 35: 283-303.
CrossRefDirect Link - Snedecor, G.W. and W.G. Cochran, 1980. Statistical Methods. 7th Edn., Iowa State University Press, Iowa, USA., ISBN-10: 0813815606, Pages: 507.
Direct Link - Subedi, K.D., C.B. Budhathoki, M. Subedi and D. Yubak, 1997. Response of wheat genotypes to sowing date and boron fertilization aimed at controlling sterility in a rice-wheat rotation in Nepal. Plant Soil, 188: 249-256.
CrossRefDirect Link - Swain, T. and W.E. Hillis, 1959. The phenolic constituents of Prunus domestica. I. The quantitative analysis of phenolic constituents. J. Sci. Food Agric., 10: 63-68.
CrossRefDirect Link - Tanaka, Y., T. Sano, M. Tamaoki, N. Nakajima, N. Kondo and S. Hasezawa, 2005. Ethylene inhibits abscisic acid-induced stomatal closure in Arabidopsis. Plant Physiol., 138: 2337-2343.
Direct Link - Trivedi, H.B., T.V.R. Rao, D.L. Bagdi and G.G. Rao, 2004. Influence of silicon on growth and salt uptake in wheat under salinity. Ind. J. Plant Physiol., 9: 360-366.
Direct Link - Watson, M.B., K.K. Emory, R.M. Piatak and R.L. Malmberg, 1998. Arginine decarboxylase (polyamine synthesis) mutants of Arabidopsis thaliana exhibit altered root growth. Plant J., 13: 231-239.
CrossRefDirect Link - Willadino, L., T. Camara, N. Boget, Y. Claprols, M. Santos and J.M. Torne, 1996. Polyamine and free amino acid variations in NaCL-treated embryogenic maize callus from sensitive and resistant cultivars. J. Plant Physiol., 149: 179-185.
CrossRefDirect Link - Wimmer, M.A., K.H. Muehling, A. Lauchli, P.H. Brown, H.E. Goldbach, 2001. Interaction of Salinity and Boron Toxicity in Wheat (Triticum aestivum L.). In: Plant Nutrition: Food Security and Sustainability of Agro Ecosystems Through Basic and Applied Research, Horst, W.J., M.K. Schenk, A. Burkert, N. Claassen, H. Flessa, W.B. Frommer, H. Goldbach, H.W. Olfs and V. Romheld (Eds.). Springer, Netherlands, ISBN: 978-0-7923-7105-2, pp: 426-427.
Direct Link - Yeo, A.R., S.J.M. Caporn and T.J. Flowers, 1985. The effect of salinity upon photosynthesis in rice (Oryza sativa L.): Gas exchange by individual leaves in relation to their salt content. J. Exp. Bot., 36: 1240-1248.
CrossRefDirect Link