
A. Karami
Department of Horticultural Science, College of Agriculture, Shiraz University, Shiraz, Iran
M. Khosh-Khui
Department of Horticultural Science, College of Agriculture, Shiraz University, Shiraz, Iran
International Journal of Agricultural Research
Year: 2007 | Volume: 2 | Issue: 1 | Page No.: 97-101







ABSTRACT
In present experiment, the nature of dormancy in seeds of wild Persian buttercup was investigated. Scarification with 50°C hot water for 2 min enhanced seed germination. The highest Germination Percentage (GP) (98.75%) and Mean Daily Germination (MDG) (10.76%) were obtained by treating the scarified seeds with 200 mg L-1 Gibberllic Acid (GA3). These treatments resulted in seed germination, but the hypocotyls grew only to approximately 2 mm in length and then ceased the growth. Moist stratification at 5°C for 30 days significantly increased hypocotyls length. It was concluded that both mechanical and epicotyl dormancies are present in wild Persian buttercup seeds.
PDF Abstract XML References Citation
How to cite this article
A. Karami and M. Khosh-Khui, 2007. Presence of Double Dormancy in Wild Persian Buttercup (Ranunculus asiaticus L.). International Journal of Agricultural Research, 2: 97-101.
DOI: 10.3923/ijar.2007.97.101
URL: https://scialert.net/abstract/?doi=ijar.2007.97.101
DOI: 10.3923/ijar.2007.97.101
URL: https://scialert.net/abstract/?doi=ijar.2007.97.101
INTRODUCTION
The cultivated Persian buttercup (Rananculus asiaticus L.) is mainly used as a perennial garden plant, cut flower or flowering potted plant. Species of Ranunculus generally found in the eastern Mediterranean area, southern Europe, Asia Minor, Syria, Iran and the Greek Island. Native populations of Persian buttercup are found in many parts of Iran. These populations have a tuberous root up to 40 cm long and single flowers with five red petals.
Although Ranunculus is normally grown by seeds, propagation by division of tuberous roots is also possible, but the annual multiplication rate is only 2 to 5 (Beruto and Debergh, 2004). Moreover, different diseases caused by viruses [e.g., Cucumber Mosaic Virus (CMV); Tomato Spotted Wilt Virus (TSWV); Tobacco Necrosis Virus (TNV); Tobacco Rattle Virus (TRV) and different potyviruses] and fungi [e.g., Fusarium oxysporum f. sp. ranunculi, Fusarium tabacinum, Pythium sylvaticum and Rhizoctonia sp.] are major factors requiring consideration in vegetative propagation (Beruto and Debergh, 2004). To overcome two aforementioned problems, breeders have developed seed lines of Ranunculus flowering the first year for cultivation.
Germination controlling mechanisms are important in nature, because they contribute to natural survival. Seed dormancy is a common condition found in many species. It is an adaptation that allows a species to determine the time of germination for seeds in a native population.
Cultivated Persian buttercup seeds germinate in one to four weeks at 20°C (Kim et al., 1991). Morphological and non-deep physiological dormancy have been reported in Ranunculus (Atwater, 1980; Geneve, 1998). Dormancy in tuberous root of this plant has been also reported (Dole and Wilkins, 1999; Dole, 2003). Acid scarification for 25 min increased seed germination in bur buttercup R. testiculatus Crantz). Gibberllic acid (0.289 mmol L-1) and 0.1 mol L-1 potassium nitrate (KNO3) synergistically enhanced the germination of acid scarified seeds of bur buttercup (Young et al., 1992). Germination in creeping buttercup (R. repens L.) normally occurred but in bulbous buttercup (R. bulbosus L.) fresh seed was normally dormant and required a period of after-ripening to achieve maximum germination (Roberts and Boddrell, 1985).
So far no report is available on the dormancies in wild Persian buttercup seeds. The objective of the present study was to determine the nature of seed dormancy and to find the optimum temperature/day for hypocotyl elongation in this species.
MATERIALS AND METHODS
Seeds of Persian buttercup were collected from a native population distributed in southern parts of Kazeroun city, Fars province, Iran. Collected seeds were cleaned and stored at room temperature in the laboratory until tested.
Each treatment contained 100 seeds placed on a filter paper in a Petri dish wetted with 5 mL distilled water. The Petri dishes were covered with a transparent polyethylene sheet to reduce the evaporation to a minimum, then in 4 different experiments Germination Percentage (GP), Mean Daily Germination (MDG) (Hartmann et al., 2002) and hypocotyl elongation were investigated. The experiments 1 and 3 were carried out as a completely randomized design with four replications per treatment and the experiments 2 and 4 were arranged in a factorial manner in a completely randomized design replicated 4 times.
Experiment No. 1
The effects of different treatments on seed germination of Persian buttercup were evaluated. The treatments consisted of control, stratification for 1 month at 5°C, scarification with hot water (50°C) for 2 min, scarification with hot water (50°C) for 2 min + stratification for 1 month at 5°C, soaking the seeds in water for 12 and 24 h, soaking the seeds in water for 12 and 24 h + stratification for 1 month at 5°C.
Experiment No. 2
Seeds were scarified with 25, 50 or 75°C hot water for 2, 4, or 6 min.
Experiment No. 3
Scarified seeds were placed in Petri dishes containing solution of 0, 25, 50, 100 or 200 mg L-1 benzyladenine (BA), 0, 50, 100, 200 or 400 mg L-1 GA3, 50 mg L-1 BA + 100 mg L-1 GA3 and 200 mg L-1 BA + 400 mg L-1 GA3.
Experiment No. 4
After scarifying the seeds with hot water (50°C) for 2 min, they were placed on filter paper in Petri dish and were exposed to moist stratification at 0, 5 or 10°C, for 10, 20, 30 or 40 days.
In all above experiments means were compared using Duncan’s New Multiple Range Test (DNMRT).
RESULTS
Experiment No. 1
The maximum seed germination percentage obtained in the treatment of hot water (50°C) for 2 min (85.75%). However, this treatment was not different with using hot water (50°C) for 2 min + stratification for 1 month at 5°C (Table 1).
Experiment No. 2
The interaction of water temperature and duration resulted in maximum germination when the seeds were treated with hot water (50°C) for 2 min and was significantly different compared to other treatments (Table 2). However hypocotyls stopped growing when they were approximately 2 mm.
Experiment No. 3
Results indicated that both GA3 and BA enhanced seed germination (Table 3), but they did not affect hypocotyl elongation (data not shown).
Maximum germination percentage and MDG were obtained when the seeds were treated with 200 mg L-1 GA3 or 200 mg L-1 BA + 400 mg L-1 GA3. Similarly, the highest GP obtained when 200 mg L-1 GA3 was used which was significantly different compared to control and other treatments.
Experiment No. 4
Hypocotyls lengths of wild Persian buttercup increased at 5 and 10°C as the days of stratification period increased, with hypocotyl lengths being longer at 5 than 10°C. The highest hypocotyl length was obtained at 5°C for 30 days. However, this treatment was not significantly different with stratifying at 5°C for 40 days (Table 4).
| Table 1: | Effects of different treatments on seed germination of wild Persian buttercup |
 | |
| † Means followed by the same letter are not significantly different at 5% of probability using DNMRT | |
| Table 2: | Effects of hot water temperature and duration of application on germination percentage of wild Persian buttercup |
 | |
| † Mean followed by same letters (small letters for means and capital letters for means of rows and columns) are not significantly different at 5% of probability using DNMRT | |
| Table 3: | Effects of GA3 and BA on seed germination of wild Persian buttercup |
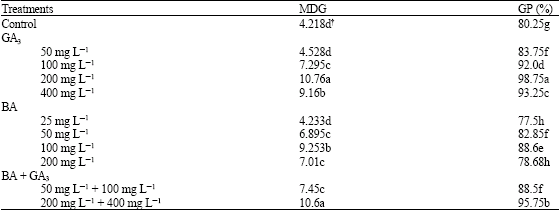 | |
| † Means in each column followed by same letters are not significantly different at 5% of probability using DNMRT | |
| Table 4: | Effects of cold stratification on hypocotyl length (mm) in wild Persian buttercup |
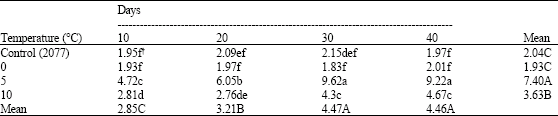 | |
| † Mean followed by same letters (small letters for means and capital letters for means of rows and columns) are not significantly different at 5% of probability using DNMRT | |
DISCUSSION
This research indicated that wild population of Persian buttercup probably has mechanical dormancy because imbibition was seen in seeds, but seed covering restricted germination. Moreover, scarification, stratification and soaking in water resulted in softening of seed covering and enhanced seed germination. This is in agreement with the results of Geneve (1998 and 2003) for some vegetable and flower species.
GA3 and BA increased seed germination and also overcame epicotyl dormancy in cultured peony (Buchheim et al., 1994). In this research GA3 and BA enhanced seed germination, but had no effect on hypocotyl elongation. It was assumed that GA3 converts complex seed reserves to simple compounds making them available for growing points and BA counteracts the effects of inhibitors. These hormonal activities could enhance seed germination.
Although no dormancy is reported for cultivated Persian buttercup (Dole and Wilkins, 1999; Hartmann et al., 2002), but apparently there are some variabilities in some species or cultivars and also wild varieties regarding seed dormancy conditions. It has been shown that in Anemone coronaria L. cultivated ‘de Caen’ seeds showed only morphological dormancy, while wild populations of Anemone had morphophysiological dormancy (Horovitz et al., 1975).
Double dormancy has been reported in Sanguinaria canadensis L., Trillium sp., Convallaria majalis L., Polygonatum multiflorum L. and Smilacina sp. seeds. Seeds in this group required a chilling period to relieve radicle dormancy, followed by a warm period to allow the radices to grow, then a second cold period to release the epicotyls from dormancy (Geneve, 1998 and 2003). Wild Persian buttercup seeds required a chilling period to relieve hypocotyls from dormancy. Presumably cold temperature reduces growth inhibitor in hypocotyls, as with increasing the cold period hypocotyl length was increased.
REFERENCES
- Beruto, M. and P. Debergh, 2004. Micropropagation of Ranunculus asiaticus L.: A review and perspectives. Plant Cell Tiss. Org. Cult., 77: 221-230.
Direct Link - Buchheim, A.T., L.F. Burkhart and M.M. Meyer, 1994. Effect of exogenous gibberellic acid, abscisic acid and benzylamonipurine on epicotyl dormancy of cultured herbaceous peony embryos. Plant Cell Tiss. Org. Cult., 36: 35-43.
Direct Link - Dole, J.M. and H.F. Wilkins, 1999. Floriculture: Principles and Species. 1st Edn. Prentice Hall, Upper Saddle River, N.J., USA.
Direct Link - Dole, J.M., 2003. Research approaches for determining cold requirements for farcing and flowering of geophytes. HortSci., 38: 341-346.
Direct Link - Young, J.A., E. Martens and N.E. West, 1992. Germination of bur buttercup seeds. J. Range Manage., 45: 358-362.
Direct Link