
M.M. Begum
Department of Plant Protection, Faculty of Agriculture, University Putra Malaysia, 43400 UPM, Serdang, Selangor D.E., Malaysia
M. Sariah
Department of Plant Protection, Faculty of Agriculture, University Putra Malaysia, 43400 UPM, Serdang, Selangor D.E., Malaysia
A.B. Puteh
Department of Crop Science, Faculty of Agriculture, University Putra Malaysia, 43400 UPM, Serdang, Selangor D.E., Malaysia
M.A. Zainal Abidin
Department of Plant Protection, Faculty of Agriculture, University Putra Malaysia, 43400 UPM, Serdang, Selangor D.E., Malaysia
International Journal of Agricultural Research
Year: 2007 | Volume: 2 | Issue: 9 | Page No.: 812-819







ABSTRACT
A study on seed-borne infections of soybean [Glycine max (L.) Merrill] demonstrated the infectivity of 17 fungal species belonging to 11 genera using blotter and potato dextrose agar methods. Among them Colletotrichum truncatum, Diaporthe phaseolorum var. sojae and Fusarium oxysporum f. sp. glycines were found in higher frequencies and well established within and on the external surfaces of seeds. Histopathology of C. truncatum infection in seeds of soybean was examined under Light Microscopy (LM) and Scanning Electron Microscopy (SEM). No mycelium and acervulus were observed in any tissues of asymptomatic (healthy) seeds. Seeds colonized with C. truncatum produced irregular gray spots with black specks. C. truncatum produced compact dark mycelium both intra- and intercellularly in the seed coat, cotyledon and embryo. Mycelial growth was more abundant in the hourglass layer of the seed coat and hypodermis, where large inter-cellular spaces were present. Acervuli with setae and abundant hyaline sickle-shaped conidial masses were observed abundantly on the surface of infected seeds. Similar observations were found beneath the inner layers of the seed coat and upper surfaces of embryo and cotyledonary tissues. Brown conidial masses were produced during incubation and liberated in the form of ooze resulting in maceration and disintegration of the parenchyma tissues of the seed coat, cotyledon and embryo.
PDF Abstract XML References Citation
How to cite this article
M.M. Begum, M. Sariah, A.B. Puteh and M.A. Zainal Abidin, 2007. Detection of Seed-Borne Fungi and Site of Infection by Colletotrichum truncatum in Naturally-Infected Soybean Seeds. International Journal of Agricultural Research, 2: 812-819.
DOI: 10.3923/ijar.2007.812.819
URL: https://scialert.net/abstract/?doi=ijar.2007.812.819
DOI: 10.3923/ijar.2007.812.819
URL: https://scialert.net/abstract/?doi=ijar.2007.812.819
INTRODUCTION
Soybean [Glycine max (L.) Merrill] seeds have achieved great attention as a vital source of vegetable oil and protein in the world today (Wang, 2002). Moreover, several soy products have played an essential part in Asian cuisine for centuries (Mckevith, 2005). Anthracnose caused by Colletotrichum truncatum [(Schw.) Andrus and Moore] is regarded as the most important seed-borne disease of soybean that reduces seed germination and seedling emergence (Srichuwong, 1992; Ploper and Backman, 1992). It is a common disease in most soybean growing areas worldwide, especially in the tropical and sub-tropical areas (Sinclair and Backman, 1989). Seeds colonized by C. truncatum showed small brown speckled lesions that are dark and shrivelled externally and sometimes remain dormant as a latent infection without showing any obvious symptom (Sinclair, 1991; Manandhar and Hartman, 1999). It was demonstrated that the seed coat is a common infection site for C. truncatum (Nik and Lim, 1984). The colonization of this fungus was also confined in the seed coat tissues with mixed infections by Phomopsis spp. or Cercospora sojina in soybean seeds (Kunwar et al., 1985).
However, limited information is available on the exact distribution and location of C. truncatum in the different seed tissues of soybean. Therefore, this study was attempted with the aim to elucidate seed-borne infections and the location of C. truncatum in naturally-infected soybean seeds using Light Microscopy (LM) and Scanning Electron Microscopy (SEM).
MATERIALS AND METHODS
The experiment was conducted at Plant Protection Laboratory, Faculty of Agriculture, University Putra Malaysia, Serdang, Selangor, Malaysia in 2006.
Isolation Techniques
Soybean seeds var. Palmetto were collected from the Department of Crop Science, Faculty of Agriculture, University Putra Malaysia, Malaysia which had been stored for one year at 10°C. Seed moisture content was maintained in the range of 8-11% throughout the storage period. Four hundred seeds were used for each method of isolation. External seed-borne infection was determined by plating the seeds directly on moist blotter paper. Internal seed-borne infection was determined by surface sterilization of seeds with 10% Clorox® for 3 min, followed by repeated washings with sterilized distilled water and then plated on potato dextrose agar. The plates were incubated for 5-7 days at ambient temperature (28±2°C) in alternate cycles of 12 h Near Ultraviolet Light (NUV) and darkness and examined for fungal infection using stereo and compound microscopes. The experiment was laid out as a Completely Randomized Design (CRD) and the mean percentage of isolation was calculated based on 400 seeds.
Histopathological Studies
Seed samples were randomly selected and plated on moist blotter paper and incubated in alternating cycle of 12 h Near-Ultra Violet (NUV) light and darkness. After four days of incubation, naturally infected seeds with typical fruiting structures of C. truncatum as well as asymptomatic (healthy) seeds as control were selected. Different seed components viz. seed coat, cotyledon and embryo were separated carefully and cut transversely into 2-3 mm2 pieces.
Light Microscopy (LM)
Individual seeds and it’s components from both infected and healthy seeds were fixed in Bouin’s solution (picric acid, formaldehyde 40%, glacial acetic acid 15:5:1 v/v/v) and vacuum extracted overnight. Samples were then dehydrated in eight different concentrations of ethanol (30, 40, 50, 60, 70, 80, 90 and 100%) and washed two times in 98% methyl benzoate plus 2% celloidine for 96 h. Samples were then infiltrated in oven at 60°C overnight in each of the mixture of xylene and paraplast with proportion of 75:25; 50:50; 25:75 and 100%, respectively. Finally, the samples were embedded in paraffin wax and mounted in small aluminum foil box. Serial paraffin sections of 10-12 μm thick were cut using an ordinary rotary microtome. Fine sections were placed on glass slides using albumin-glycerin solution. Sections were deparafinized with xylene and stained with 0.05% toluidine blue. All stained sections were mounted in DPX mounting media and viewed under light microscope (Model Nikon FX-35DX) (Johansen, 1940).
Scanning Electron Microscopy (SEM) Seed samples from symptomatic and asymptomatic (healthy) seeds were fixed separately in 2.5% buffered glutaraldehyde for 24 h at 4°C. Samples were washed with 0.1 M sodium cacodylate buffer (pH 7.7) and post-fixed in 1% osmium tetroxide for 2 h at 4°C and then washed again with 0.1 M sodium cacodylate buffer for three times 10 min each. A series of dehydration was performed in seven different concentrations of ethanol (30, 40, 50, 60, 70, 80 and 90%) three times 10 min each and finally 15 min at 100% ethanol. Samples were dried in Baltec 030 Critical Point Drying (CPD) apparatus for 30 min. Dried samples were stuck on aluminum stubs and coated with gold in a Polaron Sputter Coater and viewed under SEM (JOEL JSM 6400) (Benhamou and Chet, 1996).
RESULTS
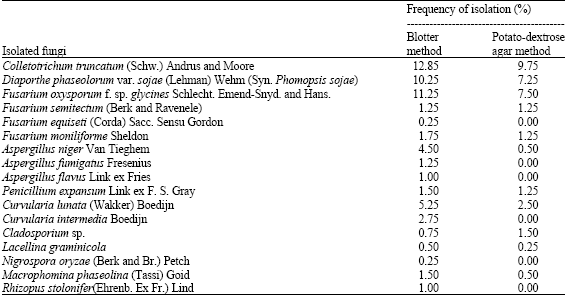
Seventeen species of fungi belonging to 11 genera were isolated from soybean seeds using both blotter and agar plate methods (Table 1). Colletotrichum truncatum, Fusarium oxysporum f. sp. glycines and Diaporthe phaseolorum var. sojae were isolated at higher frequency from both methods of isolation compared to the other fungi. The frequency of C. truncatum was 12.85 and 9.75%, F. oxysporum f. sp. glycines was 11.25 and 7.5% and D. phaseolorum var. sojae was 10.25 and 7.25% in blotter method and agar plate method, respectively. Other isolated fungi were namely Fusarium semitectum, F. equiseti, F. moniliforme, Aspergillus niger, A. fumigatus, A. flavus, Penicillium expansum, Curvularia lunata, C. intermedia, Cladosporium sp., Lacellina graminicola, Nigrospora oryzae, Macrophomina phaseolina and Rhizopus stolonifer from both methods with less than 7% frequencies.
Histopathological studies of transverse sections of healthy seeds were observed under LM and SEM. The soybean seeds contain mainly a seed coat, cotyledon and embryo (Carlson and Lersten, 1987). The seed coat contains a cuticle, palisade cell layer, hourglass cell layer with prominent air-spaces and parenchyma cell (Fig. 1A and B). The cotyledons and embryo are mainly formed by parenchyma cells (Fig. 1C, D, E and F). No fungal propagules were observed in any tissues of asymptomatic seeds where tissues of all seed components appeared to be well developed (Fig. 1A, B, C, D, E and F).
Histopathological studies of transverse sections of infected soybean using LM and SEM showed the presence of fungal propagules on the surface and inside the seed (Fig. 2 and Fig. 3). C. truncatum produced numerous acervuli with acute black setae over the seed surface. Acervuli with setae and abundant hyaline sickle-shaped conidial masses were observed on the transverse sections of the seed coat and the cuticle was distended upwards and ruptured (Fig. 2A and 3A). Acervuli were formed subsequently beneath the inner layer of seed coat on the upper surfaces of the cotyledon and embryonic tissues (Fig. 2C and 3E).
| Table 1: | Percentage infection of seed-borne fungi in soybean var. Palmetto using blotter method and potato-dextrose agar method |
 | |
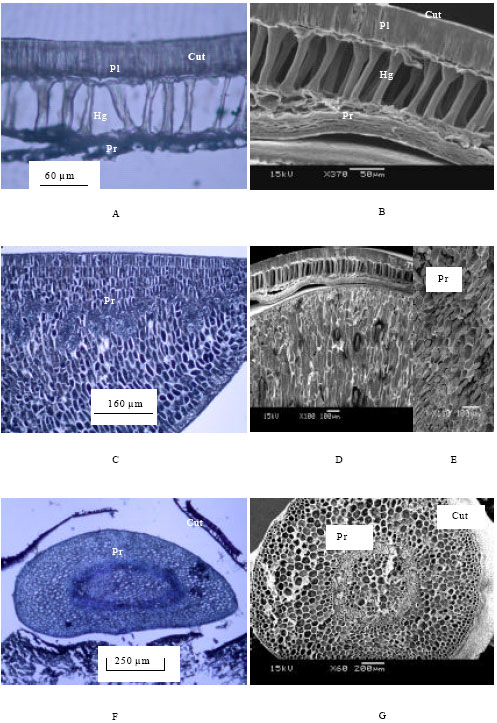 | |
| Fig. 1: | Photomicrographs showing transverse sections of seed components of asymptomatic (healthy) soybean seeds: (A) Light microscopy of seed coat (B) SEM view of seed coat (C) Light microscopy of cotyledon (D) SEM view of seed coat with cotyledon (E) SEM view of part from D magnified the tissue of cotyledon (F) Light microscopy of embryo (G) SEM view of embryo (Abbreviations : Cut, Cuticle; Pl, Palisade layer; Hg, Hourglass cell; Pr, Parenchyma cell) |
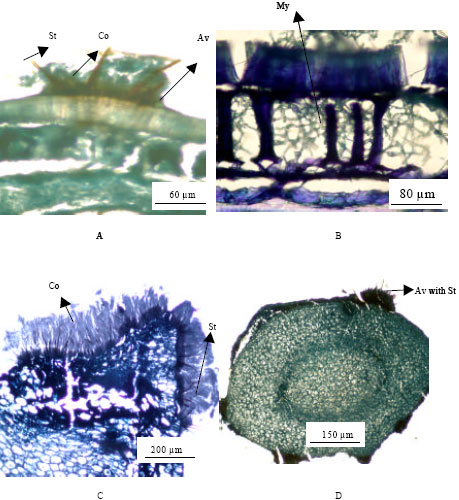 | |
| Fig. 2: | Light microscopic micrographs showing transverse sections of seed components in naturally C. truncatum infected soybean seeds (A) Acervulus on the seed coat; (B) Mycelial growth on and inner layer of seed coat; (C) Heavily infected cotyledon showing acervuli with conidial mass surrounded by black setae (D) Infected embryo showing acervuli on the ruptured cuticle (Abbreviations : Av, Acervuli; St, Setae; Co, Conidia and My, Mycelia) |
Brown conidial masses were produced during sporulation and liberated in the form of ooze. Consequently, infected parenchyma tissues of cotyledon and embryo appeared ruptured, shriveled and empty (Fig. 2C, 2E, 3B and 3D). These fungal hyphae could be distinguished in the seed tissue based on hyphal color and morphometric measurements. The hyphae of C. truncatum were dark black in color, septate and width ranges from 3.84 to 7.68 μm. A compact dark black mycelial growth was observed on the seed coat, in all seed coat layers, in the embryo and cotyledonary tissues of the seed (Fig. 2B, 3C and 3F). However, mycelial growth was more abundant in the hourglass layer of the seed coat and in the hypodermis, where large inter-cellular spaces were present (Fig. 2B and 3A).
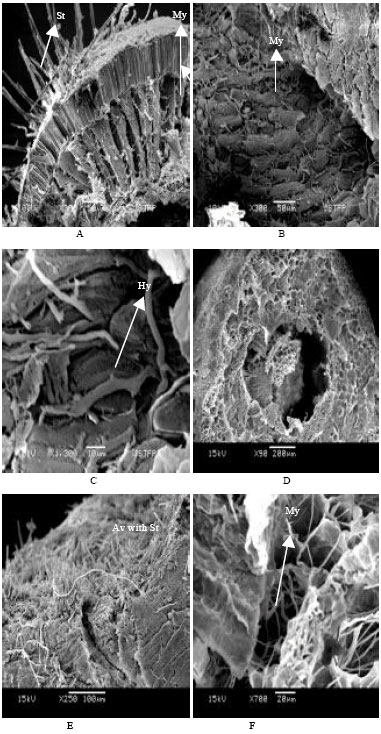 | |
| Fig. 3: | Scanning electron micrographs showing the transverse sections of seed components in naturally C. truncatum infected soybean seeds (A) Acervuli with black setae over seed surface and mycelia inside the seed coat layer (B) Infected cotyledon showing huge mycelial infection (C) Magnified view from part B showing hyphae in parenchyma tissue (D) Infected embryo showing shrivelled and empty (E) Magnified view from part D showing acervulus with setae on the cuticle of embryo (F) Magnified view from part D showing mycelia inside the parenchyma tissue of embryo (Abbreviations : Av, Acervuli; St, Setae; My, Mycelia; Hy, Hyphae) |
DISCUSSION
Colletotrichum truncatum is the most important fungus causing anthracnose disease of soybean and can produce different symptoms in a wide range of leguminous and non-leguminous host plants (Manandhar and Hartman, 1999; Mayonjo and Kapooria, 2003). This study demonstrated that soybean seeds were colonized by a number of pathogenic and saprophytic fungi. Results of isolation study revealed that C. truncatum, D. phaseolorum var. sojae and F. oxysporum f. sp. glycines were found most frequently which isolated both on moist blotter and potato-dextrose agar, suggesting that these fungi were present on the seed surface as well as inside of the seed tissues. Similar research works have documented on seed-borne infections where the most active and potential fungi recorded being C. truncatum, Phomopsis spp. and Fusarium spp. from soybean seeds (Nik and Lim, 1984; Garcia et al., 1993; Park et al.,1999; Nasir, 2003; Agarwal et al., 2006). Light microscopic and scanning electron microscopic micrographs from transverse sections confirmed that C. truncatum was internally borne in naturally-infected soybean seeds. The fungus has the ability to infect the different tissues of the seed such as seed coat, cotyledon and embryo. The acervuli with numerous setose (setae and conidia) were observed on the seed coat and cotyledon during incubation. During maturation, the acervulus ruptured and conidia were dispersed in the form of ooze and serve as a primary inoculum source for the spread of the disease (Sariah and Nik, 1988). The inter and intra cellular dark mature mycelia of C. truncatum were found in all layers of the seed coat, cotyledon and embryo and could remain dormant as a latent infection for a prolonged period without visible symtoms (Sinclair, 1991). However, mycelial growth was more abundant in the hourglass layer than that of palisade layer and parenchyma cells of the seed coat. This might be due to the large intercellular spaces which allow for more growth than closely packed palisade cell layer and endodermis (Kunwar et al., 1985).
Similar observations were also made on histopathological studies of C. truncatum infected seeds in soybean and it has been reported that it was internally seed-borne and present in the form of acervulus primordia and hyphae in the cotyledons, seed coats and embryo (Srichuwong, 1992). The present observations differ from other study such as in chili seeds infected by C. capsici (Sariah and Nik, 1988; Sariah, 1992) and C. gloeosporioides (El-Ammari et al., 1997). They found these fungal mycelia only in the cotyledon and seed coat of the infected seeds. However, other researchers have reported that C. truncatum infection and establishment occurred only in seed coat of soybean and failed to establish infection in embryo and cotyledinary tissues (Kunwar et al., 1985; Nik and Lim, 1984). These conflicting results could be due to possible interaction of mixed infections, severity of infection, environmental conditions and seed genotypes. The expansion of mycelium in seed tissues correlated directly with the infection level and favourable environmental conditions. In mixed fungal infections of seeds, fungal hyphae compete for colonization, showing antagonistic or synergistic behavior to each other which affect the extent of invasion in the seed tissues (Kunwar et al., 1985; Sariah, 1992; Singh and Mathur, 2004). Infected tissues of the seed coat, cotyledon and hypocotyl-radical axis appeared shrunk, empty and collapsed, suggesting that the cells might have been enzymatically dissolve by the fungus (Bailey et al., 1992; Srichuwong, 1992).
ACKNOWLEDGMENT
This research is a part of Ph. D. thesis. The authors are sincerely indebted to the Third World Organization for Women in Science (TWOWS) for their financial support to conduct this research at University Putra Malaysia, 43400 UPM, Serdang, Selangor D.E., Malaysia.
REFERENCES
- Agarwal, P.C., U. Dev, B. Singh, R. Indra and R.K. Khetarpal, 2006. Seed-borne fungi detected in consignments of soybean seeds (Glycine max) imported into India. Bull. OEPP, 36: 53-58.
Direct Link - Benhamou, N. and I. Chet, 1996. Parasitism of sclerotia of Sclerotium rolfsii by Trichoderma harzianum and Rhizoctonia solani. Ultrastructural and cytochemical aspects of the interaction. Phytopathology, 86: 405-416.
Direct Link - Mayonjo, D.M. and R.G. Kapooria, 2003. Occurrence and variability of Colletotrichum truncatum on soybean in Zambia. EPPO Bull., 33: 339-341.
Direct Link - Nasir, N., 2003. Detecting seed borne fungi of soybean by different incubation methods. Plant Pathol. J., 2: 114-118.
CrossRefDirect Link - Park, J.S., K.R. Lee, J.C. Kim, S.H. Lim, J.A. Seo and Y.W. Lee, 1999. A Hemorrhagic factor (apicidin) produced by toxic Fusarium isolates from soybean seeds. Applied Environ. Microbiol., 65: 126-130.
Direct Link - Sinclair, J.B., 1991. Latent infection of soybean plants and seeds by fungi. Plant Dis., 75: 220-224.
Direct Link