
Andres Araki
Facultad de Ciencias Agrarias, Universidad de Mar del Plata, Balcarce, Argentina
Jorge Rattin
Facultad de Ciencias Agrarias, Universidad de Mar del Plata, Balcarce, Argentina
Adalberto Di Benedetto
Facultad de Agronomia, Universidad de Buenos Aires, Av. San Martin, 4453 (1417), Buenos Aires, Argentina
Patricio Mirave
Facultad de Ciencias Agrarias, Universidad de Mar del Plata, Balcarce, Argentina
International Journal of Agricultural Research
Year: 2007 | Volume: 2 | Issue: 8 | Page No.: 725-730







ABSTRACT
The use of small containers for improving lettuce and celery propagation may cause root restriction which is a physical stress imposed on the root system which leads to a pronounced decrease in both root and shoot growth of the plants. Although a cytokinin decrease and root restriction relationship has been previously suggested, the physiological and biochemical processes involved are largely unknown. The following research was undertaken to show the effect of cytokinin sprays on both lettuce and celery plug nursery dry weight accumulation under a wide range of temperature and commercial yield under the summer-autumn cycle. Dry weight increase in response to a temperature increase for lettuce and celery seedlings during nursery; the better results when cytokinins were sprayed were found. The use of pre-transplant cytokinin sprays increased yield but the same doses, two weeks after the transplant, decreased it. The effects were higher for lettuce than for celery. The relationship between the cytokinins synthesis and translocations and leaf growth were discussed.
PDF Abstract XML References Citation
How to cite this article
Andres Araki, Jorge Rattin, Adalberto Di Benedetto and Patricio Mirave, 2007. Temperature and Cytokinin Relationships on Lettuce (Lactuca sativa L.) and Celery (Apium graveolens L.) Nursery Growth and Yield. International Journal of Agricultural Research, 2: 725-730.
DOI: 10.3923/ijar.2007.725.730
URL: https://scialert.net/abstract/?doi=ijar.2007.725.730
DOI: 10.3923/ijar.2007.725.730
URL: https://scialert.net/abstract/?doi=ijar.2007.725.730
INTRODUCTION
Lettuce and celery are produced throughout the season with many plantings per year and plants are therefore grown under a wide range of environmental conditions. In the juvenile phase the plant expands leaves at a certain temperature-dependent. Temperature is the main factor determining the rate of growth for both lettuce and celery during seedling emergence and the early growth period (Wien, 1997).
Technological advances in transplanting have contributed to the growth of the industry, by reducing costs and increasing reliability of production. The use of containerized vegetable transplants has increased yield and uniformity and allowed a more predictable timing of production relative to direct seeding. However, small containers may cause root restriction effects on growth and flowering, as it has been previously reported for vegetables (Peterson et al., 1991; Bar-Tal and Pressman, 1996; Rattin et al., 2006) and ornamentals (Van Iersel, 1997; Di Benedetto and Klasman, 2004, 2007).
Root restriction is a physical stress imposed on a root system which leads to a pronounced decrease in both root and shoot growth of the plants. Although it has been well established that root stress induced by volume reduction severely suppresses shoot growth (Carmi, 1995; Di Benedetto et al., 2006), the physiological and biochemical processes involved are largely unknown.
One important function of the roots is the synthesis of growth regulators such as cytokinins (Mok and Mok, 2001, Haberer and Kieber, 2002). Because various environmental conditions in the rhizosphere induce noticeable changes in the cytokinin level in xylem exudates, the hormonal fluctuation is considered to reflect the stressed physiological state of the roots. The indication that root cytokinins are part of the mechanism by which the shoot/root ratio is regulated can be drawn from a number of investigations (Howell et al., 2003; Van Norman et al., 2004; Dodd, 2005; Rashotte et al., 2005).
The following research was undertaken to show the effect of cytokinin sprays on both lettuce and celery plug nursery dry weight accumulation under a wide range of temperature and final yield under the summer-autumn cycle.
MATERIALS AND METHODS
Lettuce and celery seeds were germinated and grown in 288 plugs trays (6.18 cm3 plug-1) using a Fafard Growing Mix 2® substrate (Canadian Sphagnum peat moss-perlite-vermiculite 70:20:10 v/v). Plants were grown under greenhouse facilities at the neighborhood of Mar del Plata city, Argentina (37°45’S, 58°18’W) during 2005-2006 with a weekly fertilization according to Styer and Koranski (1997) (Stage 2: 50 mg L-1 N; Stage 3-4: 100 mg L2 N; post-transplant: 150 mg L2 N) and were irrigated with a high quality tap water.
Two Experiments Were Performed
Experiment 1 included a fortnightly sowing of Dolly-Shirley lettuce and Golden Boy-Green Fox celery seeds. Seedlings were sprayed to runoff with a 100 mg L-1 6-benzylaminopurine (SIGMA) solution when 1st true leaves pair were developed. Plants were grown at greenhouse facilities and harvested at the transplant stage.
For the Experiment 2, Dolly lettuce and Golden Boy celery seeds sowed in January 2006 were transplanted and grew in a typic argiudol soil with an organic matter of 5.2% in the first 25 cm depth. Seedlings were sprayed with cytokinin solutions (50, 100 and 200 mg L-1) when 1st true leaves pair were developed (pre-transplant lots) or two weeks later the transplant (post-transplant lots).
Plants were harvested at the transplant stage for Experiment 1; they were dried at 80°C for 48 h and weighed. Yield from Experiment 2 was expressed as fresh weight per plant. Head diameter for lettuce and both plant height and leaf number for celery were recorded at harvest too.
Half hourly averages of environmental conditions (Experiment 1) were monitoring with sensors which were continually logged with data loggers. Air temperature was measured using a HOBO H08-001-02 data logger (Onset Computer Corporation, MA, USA) protected from direct radiation by aluminum foil shades.
Dry weight accumulation vs. average air temperature relationships during the nursery pre-transplant stage (Experiment 1) were estimated through a straight-line regression. The significance of differences was determined through the test for zero slopes and test for zero intercept from Kleinbaum and Kupper (1978).
Data from Experiment 2 were subjected to a one way ANOVA for a completely design analysis and means were separated by the Tukey test (p≤0.05).
RESULTS
High correlation coefficients between the dry weight accumulation rate (DWR) and the Average Air Temperature (AAT) for Dolly and Shirley lettuce during the nursery period were found (Table 1). At the transplant stage, the higher AAT the higher DWR for both control and cytokinin-treated seedlings was found. However, when plants were sprayed with a 100 mg L-1 6-benzylaminopurine solution, there were an increase in DWR for all AAT tested; significant differences between control and cytokinin sprayed lots for Dolly and Shirley lettuce seedlings were found.
| Table 1: | The effect of cytokinin spray on Dolly and Shirley lettuce Dry Weight Accumulation (DWA) at the end of the nursery period (transplant stage) |
 | |
| AAT: Average Air Temperature | |
| Table 2: | Fresh weight (g plant-1) and Head Diameter (cm plant-1) at harvest for Dolly lettuce in response to three cytokinin solutions (50, 100 and 200 mg L-1) and two application time (pre- and post-transplant) (Experiment 2) |
 | |
| Data are the mean of twenty replicates; Different capital letter (s) indicate statistically differences (p≤0.05) between control and cytokinin spray solutions while different lower case letters indicate statistically differences (p≤0.05) for each cytokinin concentration in pre- and post-transplant times | |
| Table 3: | Dry Weight Accumulation (DWA) in Golden Boy and Green Fox celery seedlings as a function of Average Air Temperature (AAT) and cytokinin spray during nursery period |
 | |
| Table 4: | Fresh Weight (g plant-1), plant height (cm plant-1) and leaf number for Golden Boy celery at harvest in response to two application times (pre- and post-transplant) and three cytokinin concentrations (50, 100 and 200 mg L-1) (Experiment 2) |
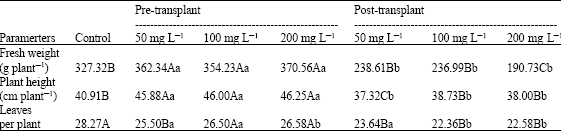 | |
| Data are the mean of twenty replicates; Different capital letter (s) indicate statistically differences (p≤0.05) between control and cytokinin spray solutions while different lower case letters indicate statistically differences (p≤0.05) for each cytokinin concentration in pre- and post-transplant times | |
There was a yield increase for Dolly lettuce at the end of Experiment 2 when plants were sprayed with cytokinin as a pre-transplant treatment (Table 2); the response showed a top at the 100 mg L-1 dose. By the other hand, increasing cytokinin concentration during the post-transplant, significantly decrease fresh weight.
There is no significant differences in lettuce head diameter at harvest between controls and pre-transplant cytokinins sprayed plants; however, a decrease was observed when the same cytokinin concentrations at the post-transplant time were used (Table 2).
Although an increase in AAT determined higher DWR for Golden Boy and Green Fox celery (Table 3), a better response with 6-benzylaminopurine at each AAT tested were found; higher correlation coefficients for both control and cytokinin treated lots were found too. Differences in straight regression line slopes were significant but lower than for the lettuce varieties (Table 1) at the end of Experiment 1.
Golden Boy celery plants sprayed with cytokinins early during nursery showed only minor yield differences related to the controls but, the same concentrations post-transplant significantly decreased fresh weight; the higher cytokinin dose (200 mg L-1) showed the worst result (Table 4).
The use of pre-transplant cytokinin sprays on Golden boy celery plants increased plant height over the controls, but did not showed statistical differences in leaves expanded per plant. However, all the post-transplant cytokinin treatments decreased both plant height and leaves per plant (Table 4).
DISCUSSION
The dry weight increase in response to a temperature increase showed in Table 1 and 3 for lettuce and celery seedlings, respectively during nursery are in agreement with previous reports (Van Henten, 1994); it was used for growth modeling and is the base for a precise greenhouse climate control.
Transplants with optimal vegetative development can better withstand the stresses caused by the field environment, pests and diseases following transplanting. Specific characteristics are claimed to be necessary to lessen transplant shock and obtain maximum yield at harvest.
The initiation and development of the root systems are highly complex processes, influenced by both endogenous and exogenous factors; cytokinins are considered to be inhibitors of adventitious root formation (Bollmark et al., 1988; Taylor and van Staden, 1997). The application of cytokinins to stem cuttings generally inhibits adventitious root formation, the early stages of the process being the most sensitive, but the mechanism for this improvement is not clear.
Simpson et al. (1982) suggested that the synthesis of cytokinins in the root depends on assimilates reallocated from the shoot; conversely, as the sink strength of the shoot increased, the cytokinin level in xylem decreased. The cytokinin synthesis in the root is considered to be a function affected rapidly by the physiological state of the shoot (Ronhzina, 2004). Although no statistical significant differences in root:shoot ratio at the transplant stage expressed as a mean of the 25 sowing dates for the lettuce and celery cultivars tested were found (data not shown), the higher total dry weight when seedlings were sprayed with a 6-benzylaminopurine solution increasing with the average air temperature. Examples where the application of cytokinins has promoted root formation, such as the results plotted in Table 1 and 3 are scarce.
Results showed both an increase and a decreasing in Dolly lettuce (Table 2) and Golden Boy (Table 4) yield for cytokinins sprays pre- and post-transplant, respectively. The beneficial response showed a top at the 100 mg L-1 dose and is in agreement with previous experiments (data not shown) using lower cytokinin levels. Cytokinins induce de novo formation of phloem elements. Ronzhina (2004) and Chernyadev (2005) showed that exogenous cytokinins contributed to enlargement of mesophyll cells and lignification of mechanical leaf tissues; the number of vascular bundles, xylem vessels and phloem elements increased and could explain the health increase in celery and fresh weight increase in lettuce and celery (Table 2 and 4).
A root restriction during nursery would give to a decrease of both root growth and cytokinin synthesis or translocation; by the other hand, a higher internal concentration when the adventitious root system begin to develop would inhibit growth processes and let explain the celery and lettuce yield decrease (Table 2 and 4), the lettuce health diameter reduction (Table 2) and both the smaller celery plant height and leaf number (Table 4) for the post-transplant applications. By the other hand, the lettuce and celery yield decrease when post-transplant applications were used would be associated to the cytokinin solution dose too; the higher cytokinin dose (200 mg L-1) showed the worst result.
Eriksen (1974) has indicated that the influence of cytokinin changes with the stage of root development. There is a possible interaction between cytokinins and other growth hormones, specifically auxins. High cytokinin endogenous concentrations may have an inhibitory effect on an early stage of rooting by blocking the activity of auxins. The loss of the inhibitory effect of cytokinin during the latter phase of initiation suggests that at this stage, developing root primordia are capable of controlling the level of active cytokinin and thus do not react to the exogenous application of its. This then implies that the control of cytokinins levels during the early phases of root initiation lies elsewhere. There is the possibility that the ability of the basal stem tissue to metabolize cytokinins, influence their transport to and from the leaves. By the other hand, it was suggested that cytokinins are essential growth substances for the latter part of the initiation phase (i.e., when root primordia develop into roots) and that partially initiated root primordia can synthesize cytokinins, being self-sufficient for these hormones. Although, it is possible that higher cytokinin rates could be related to a plant growth decrease via suppression of root growth, this hyposthesis can not be tested with the experimental design used.
Although the detailed mechanisms of cytokinin action remain to be worked out, in many cases it seems that changes in the root environment, such as root restriction during nursery, modify the hormone production in the root, change the export of hormone via the xylem sap to the shoot and thereby elicit changes in the shoot.
In order to obtain high yields of good quality for lettuce and celery, every developmental stage should be carefully managed. Although there are recent information available about the beneficial effects of cytokinin applications (OHare and Turnbull, 2004; Chernyadev, 2005; Pozo et al., 2005), commercial use of cytokinins are unusual. However, our results showed that cytokinin sprays during nursery would be a new alternative tool for suppressed root restriction effects after transplant, but a precise dose and time calibration would be necessary for improving results are needed.
ACKNOWLEDGMENTS
This study was funded by UBA C y T (Project G031/2004-2007).
REFERENCES
- Bar-Tal, A. and E. Pressmann, 1996. Root restriction and potassium and calcium concentrations affect dry matter production, cation uptake and blossom-end root in greenhouse tomato. J. Am. Soc. Hort. Sci., 121: 649-655.
Direct Link - Carmi, A., 1995. Growth, water transport and transpiration in root-restricted plants of bean and their relation to abscisic acid accumulation. Plant Sci., 107: 69-76.
Direct Link - Di Benedetto, A. and R. Klasman, 2004. The effect of plug cell volume on the post-transplant growth for Impatiens walleriana pot plant. Eur. J. Hortic. Sci., 69: 82-86.
Direct Link - Di Benedetto, A., M. Jorge and R. Jorge, 2006. The effect of transplant in sweet maize (Zea mays L.) II. Container root restriction. Int. J. Agric. Res., 1: 555-563.
CrossRefDirect Link - Di Benedetto, A. and R. Klasman, 2007. The effect of plug cell volume, paclobutrazol height control and the transplant stage on the post-transplant growth for Impatiens wallerana pot plant. Eur. J. Hort. Sci.
Direct Link - Dodd, I.C., 2005. Root-to shoot signalling: Assessing the roles of up in the up and down world of long-distance signalling in planta. Plant Soil, 274: 251-270.
Direct Link - Howell, S.H., S. Lall and P. Che, 2003. Cytokinins and shoot development. Trends Plant Sci., 8: 453-459.
Direct Link - Mok, D.W.S. and M.C. Mok, 2001. Cytokinin metabolism and action. Ann. Rev. Plant Physiol. Plant Mol. Biol., 52: 89-118.
CrossRefDirect Link - OHare, T.J. and C.N. Turnbull, 2004. Root growth, cytokinin and shoot dormancy in lychee (Litchi chinensis Sonn.). Sci. Hortic., 102: 257-266.
Direct Link - Pozo, del J.C., M.A. Lopez-Matas, E. Ramirez-Parra and C. Gutierrez, 2005. Hormonal control of the plant cell cycle. Physiol. Plant, 123: 173-183.
Direct Link - Rashotte, A.M., H.S. Chae, B.B. Maxwell and J.J. Kieber, 2005. The interaction of cytokinin with other signals. Physiol. Plant, 123: 184-194.
Direct Link - Simpson, R.J., H. Lambers and J. Michael, 1982. Kinetin application to roots and its effect on uptake, translocation and distribution of nitrogen in wheat (Triticum aestivum) grown with a split root system. Physiol. Plant, 56: 430-435.
CrossRef - Taylor, J.L.S. and J. van Staden, 1997. Variation in the level and type of cytokinin with the stage of root development in Impatiens wallerana Hook. f. stem cuttings. Plant Growth Regul., 22: 175-180.
Direct Link - Van Henten, E.J., 1994. Validation of a dynamic lettuce growth model for greenhouse climate control. Agric. Syst., 45: 55-72.
Direct Link - Van Iersel, M., 1997. Root restriction effects on growth and development of salvia (Salvia splendens). HortScience, 32: 1186-1190.
CrossRefDirect Link - Van Norman, J.M., R.L. Frederick and L.E. Sieburth, 2004. BYPASS1 negatively regulates a root-derived signal that controls plant architecture. Curr. Biol., 14: 1739-1746.
Direct Link