
Fakher Ayed
Higher Agronomic Institute of Chott-Mariem, 4042 Sousse, Tunisia
Mejda Daami-Remadi
National Institute of Agronomic Research of Tunisia, CRRHAB, Chott-Mariem, 4042 Sousse, Tunisia
Hager Jebari
National Institute of Agronomic Research of Tunisia, Street of Hèdi Karray, Ariana, 2080, Tunisia
Hayfa Jabnoun-Khiareddine
Higher Agronomic Institute of Chott-Mariem, 4042 Sousse, Tunisia
Mohamed El Mahjoub
Higher Agronomic Institute of Chott-Mariem, 4042 Sousse, Tunisia
International Journal of Agricultural Research
Year: 2007 | Volume: 2 | Issue: 8 | Page No.: 719-724







ABSTRACT
In 2005, a crown and root rot, a yellowing and a death of muskmelon plants were observed in some tunisian melon-growing regions. Isolation was made from diseased tissues. Twelve isolates of Fusarium solani were identified and used to inoculate melon-seedlings by the root dip method for 1 min. These 12 isolates caused typical symptoms of Fusarium crown rot and their pathogenicity indicated that they are identical to Fusarium solani f. sp. cucurbitae. Disease severity on melon-plants, cv. “Ananas d’Amérique”, was estimated by indexes of the leaf damage and crown rot. Analysis of variance revealed significant difference between control and inoculated plants. A significant difference was also observed between some F. solani f. sp. cucurbitae isolates in causing leaf damage and crown rot and a negative effect was observed mainly on plant fresh weight revealing impact of this pathogen on muskmelon growth.
PDF Abstract XML References Citation
How to cite this article
Fakher Ayed, Mejda Daami-Remadi, Hager Jebari, Hayfa Jabnoun-Khiareddine and Mohamed El Mahjoub, 2007. Effect of Some Tunisian Fusarium solani f. sp. cucurbitae Isolates On Muskmelon Growth and on Crown and Root Rot Severity. International Journal of Agricultural Research, 2: 719-724.
DOI: 10.3923/ijar.2007.719.724
URL: https://scialert.net/abstract/?doi=ijar.2007.719.724
DOI: 10.3923/ijar.2007.719.724
URL: https://scialert.net/abstract/?doi=ijar.2007.719.724
INTRODUCTION
In Tunisia, 6.150.000 tons of vegetables are produced on 2004. Approximately 7% of this production belongs to Cucurbitaceous species (Anonymous, 2004). Melon (Cucumis melo L.) is one of vegetables widely cultivated of this group with approximately 115.857 tons of production (Jebari et al., 2004). However, important economic losses were noted on this crop due to the development of a severe root rot and vine decline disease during the last decades. These problems were caused by some soilborne fungi, in particular Fusarium and Verticillium species (Martyn, 1983; El Mahjoub and Ben Khedher, 1987; Jabnoun-Khiareddine et al., 2006). Symptoms, including leaf yellowing and die back of the crown leaves, cortical rot of the taproot and lateral roots and discolouration of the vascular system in the root and crown, were similar to Fusarium wilt caused by Fusarium oxysporum f. sp. melonis and Fusarium crown and root rot caused by Fusarium solani f. sp. cucurbitae (Champaco et al., 1993).
During 2005 and 2006 growing seasons, melon plants, showing symptoms of root rot, were frequently observed. Isolation from diseased fragments showed presence of F. solani. Thus, the objectives of this research were to identify the forma specialis and to study the incidence of Fusarium solani f. sp. cucurbitae on muskmelon plant growth.
MATERIALS AND METHODS
Fungal Isolates
Twelve isolates of F. solani were obtained during 2005 and 2006, from tunisian melon-growing areas, by isolation from melon plants showing typical symptoms of crown and root rot. Surface disinfected diseased fragments were placed on Potato Dextrose Agar (PDA) containing 300 mg L-1 of streptomycin sulfate and incubated at 25°C for 4-5 days in the dark (Table 1). Single-spore cultures of all isolates were stored at -20°C.
For plant inoculation, mycelium was taken from the edge colony of each isolate and placed in Erlenmeyer flasks containing 150 mL of Potato Dextrose Broth (PDB) and incubated at 25°C for 5 days in a rotary incubator (120 rpm). The liquid culture was filtered and the conidial suspension was adjusted to 107 conidia mL-1 by a Malassez cystometer.
Plant Material
Seeds of the melon cultivar “Ananas d’Amérique”, the most cultivated cultivar in Tunisia, were disinfected by immersion for 2 min in fresh 10% Ethanol. They were then rinsed in sterile water and placed on moistened cotton wool in Petri dishes in the dark at 27±2°C. The germinated seeds were planted in an autoclaved peat and incubated in a growth chamber at 26±1°C by day and 20±1°C by night with 14 h photoperiod.
Pathogenicity Tests
Pathogenicity of the isolates was established by dipping seedlings, at the expansion of the first-true leaf, for 1 min in the inoculum suspension adjusted at 107 conidia mL-1 (Jacobson and Gordon, 1988). Five seedlings were inoculated with every isolate and five seedlings dipped in sterile distilled water served as controls. The whole were transplanted to plastic pots (0.7l) filled with an autoclaved mixture of peat and perlite (3:1) and placed in a growth chamber at 26±1°C by day and 20±1°C by night with 14 h photoperiod. They were irrigated regularly and fertilized with the nutrient solution of El Mahjoub (1985) (per liter): 0.225 g of KNO3, 0.3 g of KH2PO4, 0.225 g of NH4Cl, 0.45 g of MgSO4,7H2O, 0.3 g of (NH4)2SO4, 1.2 g of Ca(NO3)2, 0.045 g of EDTA Na, Fe and other oligo-elements.
Symptoms, as described by Champaco et al. (1993), traduced by leaf yellowing, crown or cortical rot were first observed 20 days after inoculation. Fungal isolates implicated in typical symptom development, determined via re-isolations made from rotted roots and crown of inoculated melon seedlings, were considered pathogen and then attributed to the forma specialis cucurbitae.
| Table 1: | Isolates of Fusarium solani from some melon cultivars and their geographic origin |
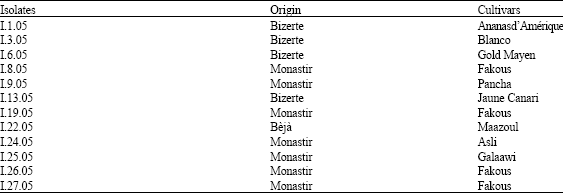 | |
Disease Incidence on the Melon Growth
Disease severity was assessed via an index of leaf damage (ILD) where a scale of 0-4 was used every 3-4 days: 0 = asymptomatic leaf, 1 = leaf wilted, 2 = leaf with yellowing, 3 = leaf with necrosis, 4 = dead leaf. Isolate’s virulence were expressed as an index of leaf damage (ILD) which is calculated per muskmelon plant (Béye and Lafay, 1985):
ILD = Σ notes/max |
| ILD | : | Index of leaf damage. |
| Σ notes | : | Total notes. |
| Max | : | 4 times of the developed-leaves number. |
Another scale of 0-3 was used, 48 days after inoculation, to assess disease severity on crown necrosis or rots: 0 = asymptomatic crown, 0.5 = some necrotic traces, 1 = necrosis of 1/3 of the crown, 2 = necrosis of 2/3 of the crown, 3 = necrosis of more than 2/3 of the crown.
Plant height and fresh weight was also recorded 48 days after inoculation. Pathogen isolations were done at the end of the bioassay.
Statistical Analysis
Five plants were used for each elementary treatment (isolates and control). Analysis of variance were conducted for each dependant variable to determine the disease severity and the effect of isolates on plants development. Data are arranged by completely randomized design where treatments (seedlings inoculated by each of 12 Fusarium isolates and control seedlings) are the only fixed factor. They were analyzed using SPSS and subjected to analysis of variance and Fisher’s least significant difference test LSD (at p≤0.05).
RESULTS
Pathogenicity Tests of F. solani
Several fungal species were isolated from rotted root and crown melon samples on PDA, but Fusarium species, especially F. solani, were predominant. In pathogenicity tests, F. solani isolates used were pathogenic by inducing yellowing, 20 days after inoculation and death of inoculated seedlings. Similar symptoms developed in naturally diseased plants. This result showed that it is the forma specialis cucurbitae race 1. The fungus was re-isolated from rotted roots and crown of all inoculated seedlings.
Incidence of F . solani f. sp. cucurbitae on the Development of Muskmelon Plants
Effect of F. solani f. sp. cucurbitae Inoculation on the Leaf Damage Index
The incidence of 12 F. solani f. sp. cucurbitae isolates from muskmelon was studied by the root dip method inoculation of seedlings. On the basis of analysis of variance of disease severity from seedlings inoculated with some isolates, statistically significant differences were found in disease severity among some isolates and between healthy and some diseased plants (Table 2). Twenty to twenty four days after inoculation, I.24.05, I.26.05 and I.19.05 were found to be the most virulent isolates. Their leaf damage index reached 0.2627, 0.2166 and 0.2168, respectively. However, I.6.05, I.8.05 and I.22.05 had the least ILD. with 0.025, 0.0332 and 0.0166, respectively. No significant difference between indexes of inoculated plants was observed from 28 days after inoculation (Table 2). At the end of the bioassay, I.6.05 appeared the most pathogenic with an ILD of about 3.5; but I.22.05 was the least with 0.3642.
| Table 2: | Evolution of the Index of Leaf Damage (ILD) for muskmelon plants, cv. “Ananas d’Amérique”, inoculated by F. solani f. sp. cucurbitae isolates observed 20 days after inoculation |
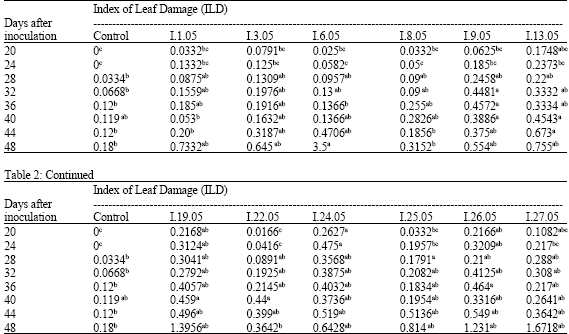 | |
| In.05: F. solani f. sp. cucurbitae isolates.zWithin lines, means followed by the same letter(s) are not significantly different (p≤0.05) according to SNK test | |
| Table 3: | Effect of inoculation by different F. solani f. sp. cucurbitae isolates on muskmelon plant growth (cv. “Ananas d’Amérique”), observed 48 days after inoculation |
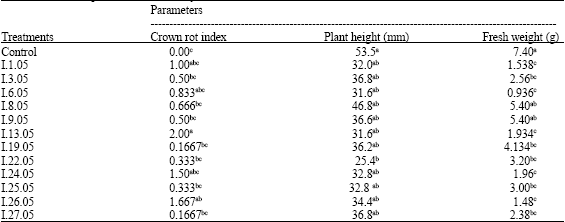 | |
| zWithin column, means followed by the same letter(s) are not significantly different (p = 0.05) according to SNK-test | |
Effect of F. solani f. sp. cucurbitae Inoculation on the Crown Rot Index
Analysis of variance revealed significant difference between control and inoculated plants. A significant difference was observed between some isolates in causing crown rot (Table 3). The most important crown rot index was caused by the isolate I.13.05. A significant necrosis was observed on crown of plants inoculated with isolates I.24.05, I.26.05, I.1.05 and I.6.05. However, these symptoms were not present on non-inoculated plants (Table 3).
Effect of Fusarium Crown and Root Rot on Melon Plant Height
Significant difference was observed between noninoculated and inoculated plant heights. Moreover, variance analysis showed a significant difference between F. solani f. sp. cucurbitae isolates. In fact, I.22.05 had the most important height reduction with 53% comparatively to control plants. Furthermore, isolates I.1.05, I.6.05 and I.13.05 reduced plant height by 40 to 52% (Table 3).
Effect of Fusarium Crown and Root Rot on Plant Fresh Weight
Significant decreases in the fresh weight of muskmelon plants inoculated with F. solani f. sp. cucurbitae were noted compared to control plants (Table 3). In fact, all inoculated melon plants showed varying degree of fresh weight reduction. Isolate I.6.05, I.1.05 and I.26.05seemed to have the most important effect in reducing weight by 87.35, 80 and 79.22%, respectively when compared to the noninoculated control. I.13.05 and I.24.05also revealed very aggressive. However, I.8.05 and I.9.05 had the least effect on muskmelon plant fresh weight (Table 3).
DISCUSSION
Twelve F. solani isolates were obtained from crown and root rot affecting melon plants in Tunisia. They are responsible of a cortical rot at the stem base and the upper portion of the taproot causing yellowing and wilting of leaves. These symptoms were identical that those described by Martyn (1996) and Armengol et al. (2000) caused by F. solani f. sp. cucurbitae race 1 (Tousson and Snyder, 1961; Messiaen et al., 1991). This pathogen caused important damage to muskmelon traduced by leaf alteration, crown and root rot development and plant fresh weight losses. In the same way, the negative effect was observed by Boughalleb et al. (2005) on watermelon in Tunisia.
This pathogenic effect of F. solani is due to the secretion of a broad range of hydrolytic enzymes involved in penetration and colonization of host plant tissues during infection (Walton, 1994). Thus, isolates of F. solani f. sp. cucurbitae produce similar lesions on all plant parts once the cuticule and epidermis have been broken (Hawthorne et al., 1994). Annis and Goodwin (1997) reported that the importance of cell wall degrading enzymes to pathogenicity might be different in each plant-microbe interaction. It has been also proposed that cell wall depolymerases can function as signals that trigger various plant physiological processes such as the induction of plant defence responses (Walton, 1994).
In this study, the incidence of different F. solani f. sp. cucurbitae isolates, obtained from diverse tunisian regions, was showed on susceptible muskmelon seedlings. A difference in their pathogenicity was observed. This result confirmed that obtained by Nagao et al. (1994). Although the fungus causing Fusarium crown and root rot of muskmelon has been taxonomically classified as Fusarium solani f. sp. cucurbitae, many researchers was indicated that this fungus has a broader Cucurbitaceae host range than only muskmelon (Blancard et al., 1991; Jebari et al., 2004). For this reason and as F. solani is a soilborne pathogen surviving in the soil and in some vegetable debris (Messiaen et al., 1991), its control revealed very necessary as it affected plant weight, growth and consequently production of an important culture in Tunisia. Thus, the research of some chemical and biological control measures were necessary.
ACKNOWLEDGMENTS
The authors would like to thank the Ministry of Scientific Research, Technology and Competency Development of Tunisia and Higher Agronomic Institute of Chott-Mariem (ISAg-CM) for financial contribution. Many thanks for Mohamed Youssef and Aymen Youssef for their excellent technical assistance.
REFERENCES
- Annis, S.L. and P.H. Goodwin, 1997. Recent advances in the molecular genetics of plant cell wall-degrading enzymes produced by plant pathogenic fungi. Eur. J. Plant Pathol., 103: 1-14.
CrossRefDirect Link - Armengol, J., C.M. Jose, M.J. Moya, R. Sales, A. Vicent and J. Garcia-Jimenez, 2000. Fusarium solani f. sp. cucurbitae race L., a potential pathogen of grafted water melon production in Spain. EPPO Bull., 30: 179-183.
CrossRefDirect Link - Boughalleb, N., J. Armengol and M. El Mahjoub, 2005. Detection of races 1 and 2 of Fusarium solani f. sp. cucurbitae and their distribution in watermelon fields in Tunisia. J. Phytopathol., 153: 162-168.
CrossRef - Champaco, E.R., R.D. Martyn and M.E. Miller, 1993. Comparison of Fusarium solani and F. oxysporum as causal agents of fruit rot and root rot of muskmelon. Hortic. Sci., 28: 1174-1177.
Direct Link - Hawthorne, B.T., R.D. Ball and J. Rees-George, 1994. Genetic analysis of variation of pathogenicity in Nectria haematococca (Fusarium solani) on Cucurbita sp. Mycol. Res., 98: 1183-1191.
Direct Link - Jabnoun-Khiareddine, H., M. Daami-Remadi, F. Ayed and M. El-Mahjoub, 2006. First report of Verticillium wilt of melon caused by Verticillium dahliae in Tunisia. New Dis. Rep.
Direct Link