
M. Hasinur Rahman
Department of Soil Science, National University, Gazipur 1704, Bangladesh
T.A. Adjadeh
Department of Soil Science, University of Ghana, Legon, Accra, Ghana
H. Saitoh
Faculty of Agriculture, Iwate University, Morioka 020-8550, Japan
S. Saiga
The United School of Agricultural Sciences, Iwate University, Morioka, Japan
International Journal of Agricultural Research
Year: 2007 | Volume: 2 | Issue: 8 | Page No.: 655-666







ABSTRACT
In this study, it is hypothesized that plants selected and/or bred for improved desired characteristics (e.g., macronutrients) may affect other characteristics (e.g., micronutrients) negatively. To test the hypothesis, we evaluated the metal micronutrient concentrations of four experimental strains viz., HighMgE, HighMgM, LowEq and LowMg and two commercial cultivars viz., Okamidori and Akimidori of orchardgrass. There were no genetic differences among the experimental strains/cultivars of orchardgrass for dry matter accumulation. The concentration of micronutrients varied with genetic differences among the four strains and the two commercial cultivars of orchardgrass. The amounts of Mn, Fe, Zn, Cu, Mo, Co, Ni, Se and I in the shoot, irrespective of strain or cultivars used, were 76.36, 74.83, 14.03, 9.36, 2.19, 5.00, 5.17, 2.00 and 19.33 μg g-1 at first harvest and 85.89, 74.56, 15.06, 12.30, 3.84, 6.14, 4.99, 2.54 and 19.53 μg g-1 at second harvest, respectively. The average amounts at the third harvest were 119.97, 90.36, 17.67, 13.39, 3.36, 4.56, 4.89, 2.36 and 20.06 μg g-1, respectively, whereas the corresponding amounts at the fourth harvest were 128.22, 156.00, 21.92, 10.31, 4.25, 4.53, 4.54, 1.47 and 28.72 μg g-1, respectively. Apart from few exceptions, Mn, Fe, Zn, Mo and I levels were highest in the fourth harvest, whereas the levels of Co and Se were highest in the second harvest. The level of Cu was the highest in the third harvest. The levels of Mn, Fe, Ni, Se and Mo were higher in low Mg containing plants irrespective of strain or cultivar. On the other hand, Co, Cu and I tended to peak in plants containing higher amounts of Mg. The Cu/Mo ratio was highest in the first harvest followed by the third, second and fourth harvests. The amounts of Mn, Fe, Ni, Co, Zn and Mo were within the optimum levels required for maintaining forage quality.
PDF Abstract XML References Citation
How to cite this article
M. Hasinur Rahman, T.A. Adjadeh, H. Saitoh and S. Saiga, 2007. Genetic Variability in Micronutrients Composition of Orchardgrass (Dactylis glomerata L.). International Journal of Agricultural Research, 2: 655-666.
DOI: 10.3923/ijar.2007.655.666
URL: https://scialert.net/abstract/?doi=ijar.2007.655.666
DOI: 10.3923/ijar.2007.655.666
URL: https://scialert.net/abstract/?doi=ijar.2007.655.666
INTRODUCTION
All plants and animals require nutrients in order to maintain their life cycles. The most important sources of nutrients for animals are forage grasses. Metabolic disorders in ruminants have been associated with imbalances of minerals in forages (Hill and Guss, 1976; Sleper et al., 1989). Developing cultivars to minimize the risk of metabolic disorder is an important task for grass breeders. It is important that cattle could obtain optimum level of nutrients while grazing. Grass tetany induced by Mg deficiency, may be the most important health problem in ruminants caused by mineral imbalances (Mayland and Hankins, 2001). Thus, the importance of producing and feeding good quality forage to animals has been clearly established and the potential for genetic improvement of plant constituents associated with quality is particularly appealing. Breeding program for high Mg concentrations has already been conducted for several forage species by producing Magnet Italian ryegrass (Hide and Thomas, 1981), HighMag tall fescue (Mayland and Sleper, 1993) and Mgwell orchardgrass (Saiga et al., 2002). Micronutrients deficiencies and toxicities in forages cause various hazards in grazing animals all over the world. While some crops show micronutrient deficiencies, others growing on the same types of soils in the same localities, do not exhibit such deficiencies. In many places, it is more common practice to apply micronutrients to high value crops such as vegetables and fruits than to grains or forages. This discriminatory application of micronutrients to high value crops may be partly due the fact that their nutrient requirements have been more extensively studied. Zinc fertilization of citrus trees is a common practice in the USA (Viets, 1966), although it may not be used on other crops growing in the same locality. Similarly, the use of B fertilizers is very common in apple cultivation but very rarely used for wheat. Also, varieties of the same crop species may differ in their susceptibility to trace element deficiencies and toxicities. Varietal differences in susceptibility to Fe deficiencies have been described by Brown (1961) and to Mn toxicity by Foy et al. (1969). There is a relationship between genotype and nutritive value of forage grasses. The nutritive value of any forage plant is also influenced by the stage of growth at which it is harvested and the environmental conditions under which it grew (Riewe and Mordart, 1985). Introducing new cultivars to minimize the hazardous effects of nutrient deficiencies and toxicities is an important concern for grass breeders. Many grass breeders have studied genetic variations within species with respect to mineral elements associated with grass tetany. Results have indicated that crop genetic factors were more important than fertility, soil and environmental factors in determining the concentration of various trace elements in selected crops (Wolnik et al., 1983, 1985). Thus, the above discussion concludes that it is important to measure micronutrients in plants, which are bred for high Mg concentration. However, there is almost no published information available on genetic variability of micronutrients in orchardgrass strains and/or cultivars, which are bred for increased Mg concentration. Present study has provided information on the relationship between genetic variability of cultivars of orchardgrass and their ability to accumulate Mg.
MATERIALS AND METHODS
Experimental Field and Soil
The experiment was initiated in 2000-2001 on an Umbric Andisol under rainfed conditions at the Iwate University Experimental Farm, Japan. Meteorological data such as temperature, relative humidity, wind speed, solar radiation, sunshine, rainfall and snowfall of the experimental site during the growing period is shown in Table 1. The soil was sandy loam and had pH 6.02. It was naturally low in P (6.71 mg 100 g-1 soil) but high in other nutrients such as N (0.34%), K (39.6 mg 100 g-1 soil), Ca (316 mg 100 g-1 soil), Mg (31.8 mg 100 g-1 soil), Cu (1.42 μg 100 g-1 soil), Mn (29.4 μg 100 g-1 soil), Fe (98.8 μg 100 g-1 soil) and Zn (2.18 μg 100 g-1 soil) in the top 15 cm composite soil sample.
| Table 1: | Meteorological data of experimental field during the experiment* |
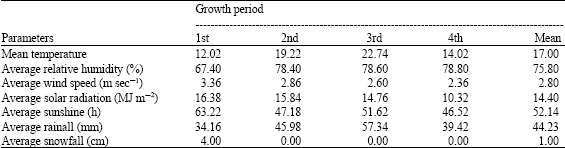 | |
| *Source: Morioka Meteorological station, Iwata, Japan | |
The soil showed a high level of P-retention capacity (1962) and organic matter content (184 g kg-1).
Experimental Plants and Management
Four experimental strains of orchardgrass viz., HighMgE, HighMgM, LowEq and LowMg and two commercial cultivars viz., Okamidori and Akimidori which differed in morphological and chemical properties and also showed genetic variability (Saiga et al., 2002) were used in this experiment. HighMgE, HighMgM and Okamidori were high magnesium containing plants bred to minimize tetany risk. The nutrients applied to the plants were 180 kg N, 180 kg P2O5 and 160 kg K2O ha-1 in early spring and immediately after each harvest. Forage was harvested (6 cm above ground) at four different times (i.e., 31st May, 19th July, 6th September and 24th October) maintaining 49 days growing period.
Plant Analysis
The samples were dried at 80°C for 24 h in a forced-air oven. After drying, samples were ground to pass through a 0.5 mm mesh with a cyclone mill and 0.5-1.0 g sample was pressed (with a coherent disc of 2.5 cm) by applying 15.0 tons pressure to make a pellet with a uniform surface. Concentrations of minerals on both sides of the pellet were measured with a live time of 100 sec by energy reflectance X-ray fluorescence spectrometry (JEOL Co., JSX-3200, Element Analyzer) as described by Hutton and Norrish (1977) and Norrish and Hutton (1977). Each plant sample was replicated three times and the results were obtained as dry matter percentage (% DM). Average data for the growing seasons of 2000 and 2001 were evaluated together.
Experimental Design and Statistics
The experimental design was a randomized complete block design with three replications in an individual plot area of 6 m2. Statistical analysis was performed using least square ANOVA procedure of SAS and means were separated using the Duncan’s Multiple Range Test (Duncan, 1955) at 5% level of significance. The Least Significant Differences (LSD) test was also used to determine whether differences between means were statistically significant (p<0.05). Pearson correlation coefficients were determined among the nutrients.
RESULTS
Dry Matter Production
The highest dry matter yield was obtained in LowMg and lowest in HighMgE (Table 2). Dry matter accumulation decreased with increasing harvesting time for all the strains as well as cultivars although there were no significant differences in dry matter accumulation among the strains/cultivars across each harvest as well the total of four harvests.
Manganese
Manganese content of the plants increased gradually from the 1st harvest to the 4th harvest (Table 3). In the 1st harvest, Okamidori showed the highest level of Mn (86.17 μg g-1) while HighMgM showed the lowest amount (62.67 μg g-1). LowMg showed the largest amounts of Mn in the 2nd and 3rd harvests, while HighMgM and Okamidori recorded the smallest amounts in the 2nd and 3rd harvests, respectively. In the 4th harvest, Akimidori showed the largest amount of Mn while LowEq showed the smallest. In all the harvests, the HighMg containing strains/cultivars tended to show low levels of Mn. The mean Mn contents of the four harvests ranged from 76.36-128.22 μg g-1 regardless of strains/cultivars.
| Table 2: | Dry matter yield (t ha-1) of orchardgrass grown on an Andisol over two years |
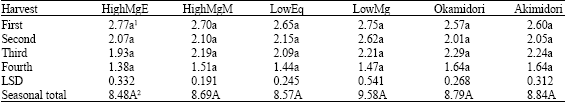 | |
| 1,2Mean in a row within the harvest followed by the same letter(s) are not statistically different at p<0.05 | |
| Table 3: | Micronutrients in the shoot of six orchardgrass grown on an Andisol |
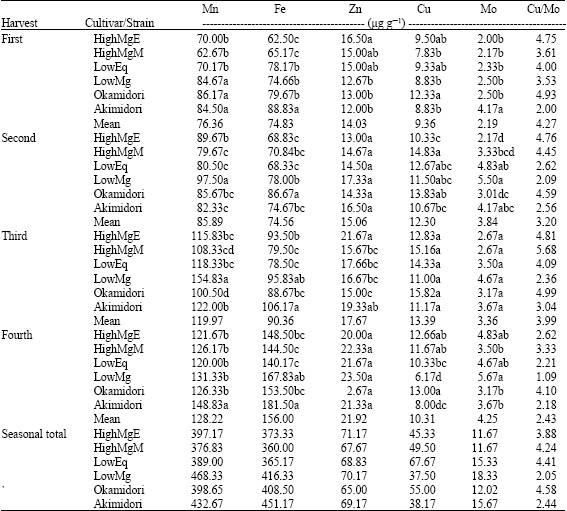 | |
| 1Means in a column within the harvest followed by the same letter(s) are not statistically different at 0.05 probability level | |
Iron
Iron content (Table 3) of the plants increased gradually from the 1st harvest to the 4th harvest. LowEq showed the lowest levels of Fe in all the harvest except in the 1st harvest while Akimidori showed the largest amounts in all the harvest except in the 2nd harvest. HighMgE showed the lowest level of Fe in the 1st harvest. Okamidori contained the largest amount of Fe in the 2nd harvest. From the mean values of the four harvests, LowMg showed higher Fe concentrations than HighMg. The mean Fe levels in all the strains/cultivars ranged from 74.56-156.00 μg g-1.
Zinc
The highest amount of Zn was recorded in the 4th harvest (Table 3). In the 1st harvest, HighMgE and Akimidori showed the highest and lowest levels of Zn, respectively. In the 2nd harvest, however, HighMgE and Akimidori, showed a trend opposite to that of the 1st harvest. The mean values of the four harvests showed that HighMgE and Okamidori had the highest and lowest levels of Zn, respectively.
Copper
The concentration of Cu increased up to the 3rd harvest and then decreased regardless of strain or cultivar (Table 3). In the 1st harvest, the HighMg containing cultivar, Okamidori, showed the highest level of Cu whereas, the HighMg containing strain, HighMgM, showed the lowest level. In the 2nd harvest, the highest level of Cu was observed in HighMgM and the lowest level in HighMgE. In both the 3rd and 4th harvests, the highest and lowest levels of Cu were observed in Okamidori and LowMg, respectively. However, in the 3rd harvest, the levels of Cu in the Okamidori and LowMg were not significantly different.
Molybdenum
Molybdenum content of orchardgrass cultivars/strains increased steadily from the 1st harvest to the 4th harvest except in the Akimidori (Table 3). In Akimidori, Mo content tended to decrease from the 1st harvest to 4th. However, it is clear from the mean harvest results that the amounts of Mo tended to increase steadily in all strains/cultivars with increasing number harvests. From the mean harvest results, it was observed that the HighMg containing plants showed lower Mo concentration while LowMg showed higher levels of Mo.
Cobalt
Cobalt concentrations in the strains/cultivars were highest in the 2nd harvest followed by the 1st, the 3rd and then the 4th harvest (Table 4). Okamidori, Akimidori, LowMg and HighMgM showed the lowest levels of Mo in the 1st, 2nd, 3rd and 4th harvests, respectively. The largest amounts of Co were observed in the HighMg containing plants in all the harvests as well as in the mean results. The strains/cultivars showed mean Co levels from 4.53-6.14 μg g-1 whereas the harvest means ranged between 4.37 and 6.14 μg g-1. Increase in air temperature during the growing period (Table 1) appeared to have been accompanied with decrease in Co content.
Nickel
No definite trend was observed in Ni concentration in the various orchardgrass strains/cultivars harvests (Table 4). In the 1st and 2nd harvests, differences in Ni concentrations were not statistically significant. In the 3rd harvest, LowMg and Okamidori showed significantly the highest and lowest levels of Ni. In 4th harvest, the largest amount of Ni was observed in LowMg whereas the smallest amount was in HighMgE. The mean Ni levels from four harvests of the strains/cultivars ranged from 4.54-5.17 μg g-1.
Selenium
Orchadrgrass grown on an Andisol in the temperate region showed large seasonal variations in Se concentrations (Table 4). The mean Se content of the strains/cultivars ranged from 1.47-2.54 μg g-1 whereas the harvests contained mean values, which ranged from 1.38-3.29 μg g-1. In the 1st harvest, the LowMg containing strains as well as the cultivar showed high concentration of Se. On the other hand, in the 2nd and 4th harvests, the LowMg containing strain, LowMg, showed the largest amount of Se whereas the HighMg containing cultivar, Okamidori, showed the lowest level of Se. In the 3rd harvest, LowMg and HighMgM contained significantly the highest and lowest levels of Se, respectively.
| Table 4: | Benificial nutrients in the shoot of six orchardgrass grown on an Andisol |
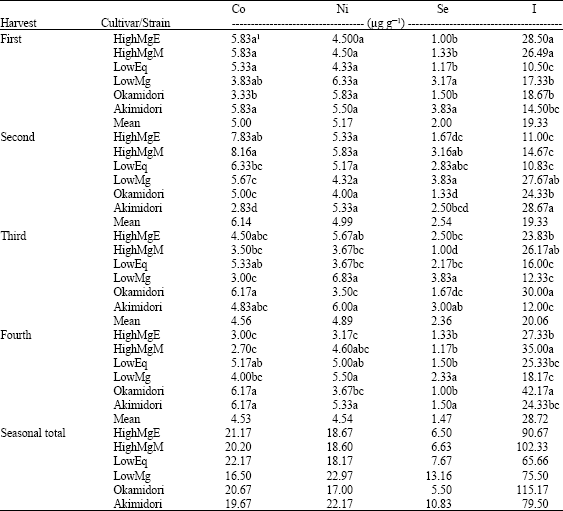 | |
| 1Means in a column within the harvest followed by the same letter(s) are not statistically different at 0.05 probability level | |
Iodine
The concentration of I in the plants varied with their genetic variations. Apart from few exceptions, I content increased steadily from the 1st to the 4th harvest (Table 4). Okamidori showed the highest mean I level (28.67 μg g-1) in the harvests whereas, LowEq showed the lowest value (10.50 μg g-1). The HighMg containing plants showed high I levels in all the harvests except in the 2nd.
Cu/Mo Ratio
In the 1st and 2nd harvests, the highest Cu/Mo ratio was observed in HighMgE (Table 3). In the 3rd harvest, the highest Cu/Mo ratio was observed in HighMgM followed by the level observed in Okamidori. In most cases, HighMg containing plants showed the highest ratios. The same trend was followed by the mean harvest results.
Relationship Between the Nutrients
Various correlations were observed between macro- and micronutrients (Table 5). Manganese and Fe correlated positively with Na, P, S and K and negatively with Mg and Ca. Cobalt correlated negatively with Na, P and K and positively with Mg and Ca. Nickel and Se correlated positively with Na, P and K and negatively with Mg and Ca. Copper and I correlated negatively with Na, S and K. Strong correlations were observed among the micronutrients. Manganese correlated positively with Fe, Ni, Zn, Se and Mo and negatively with Co, Cu and I. Iron correlated positively with Ni and Se and negatively with Co and Cu. Cobalt correlated positively with Cu and negatively with Ni, Se and Mo.
| Table 5: | Phenotypic correlation among the nutrients in the shoot of six orchardgrass in harvest means |
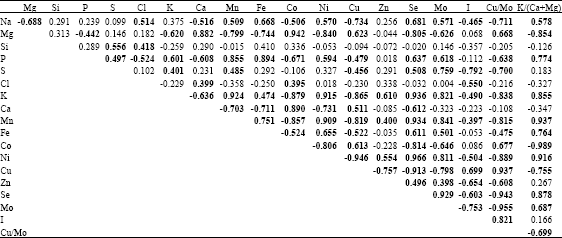 | |
| Bold values are significant at p<0.05 | |
Selenium correlated positively with Mo and negatively with I. Molybdenum showed negative correlation with I.
DISCUSSION
This study has shown substantial decreases in dry matter yield with increasing number of harvests. Usually, HighMg containing plants showed slightly lower yield whereas LowMg containing plants showed higher yield irrespective of strain or cultivar although the values were not statistically significant. Sabreen (2004) also did not find any significant differences in dry matter accumulation among the experimental strains/cultivars of orchardgrass grown in an Andisol.
Changes in environmental factors have a great effect on micronutrient (Moraghan and Mascagni, 1991). During this study, ambient temperature increased gradually up to the 3rd harvest and then decreased. However the concentrations of Mn, Fe and Zn increased up to the 4th harvest. The result may be due to the fact that the soil temperature did not decrease proportionately with decreasing air temperature (Brady, 2000a). It may also have been due to some physiological changes in the plants during the 4th regrowth period. Reid and Hovarth (1980) found that an increase in soil temperature increased Mn accumulation in barely and concluded that plant Mn accumulation was strongly correlated with soil temperature. Climatic factors greatly affect the occurrence of Fe deficiency in plants under field conditions. Temperature changes may either enhance or suppress Fe deficiency depending on their effects on rate of growth, plant metabolism and the status of other elements in the soil (Brown, 1961). Seasonal variations of Zn concentration have been found in A. sativa (Moraghan and Mascagni, 1991), D. dactylon (Kappel et al., 1985) and mixed temperate species (Masters and Sommers, 1980). Copper content of the plants increased up to the 3rd harvest and then slightly decreased in the 4th harvest (Table 3). Seasonal variation of Cu concentration in plants can be caused by differences in temperature (Minson, 1990). Air temperature increased gradually during the growing period (Table 1). Moraghan and Mascagni Jr. (1991) found that increasing soil temperature raised Cu concentration in T. subterrneum. Increase in air temperature, relative humidity and rainfall during the growing period (Table 1) were accompanied with increase in Cu content. Copper is a mobile micronutrient is soil. Movement of Cu into plants depends on the exploitation of soil by the root (Javis and Whitehead, 1981). Since environmental factors affect root development, they indirectly influence the availability of Cu to plants. Copper concentration in forages decreases with increasing age due to a decrease in the portion of leaf in grass, but the effect varies with season of the year (Marschner, 1995).
The amount of Mn in the strains as well as the cultivars (Table 3) were at optimum levels at the first and second harvests and slightly higher at the third and fourth harvests when compared to the adequate range for grasses (Marschner, 1995; Mayland and Shewmaker, 2001). However, the average levels of Mn from the four harvests over two years were within the optimum range for plant nutrition (Table 6).
| Table 6: | Mineral content of cool-season grasses and their requirement by ruminants1 |
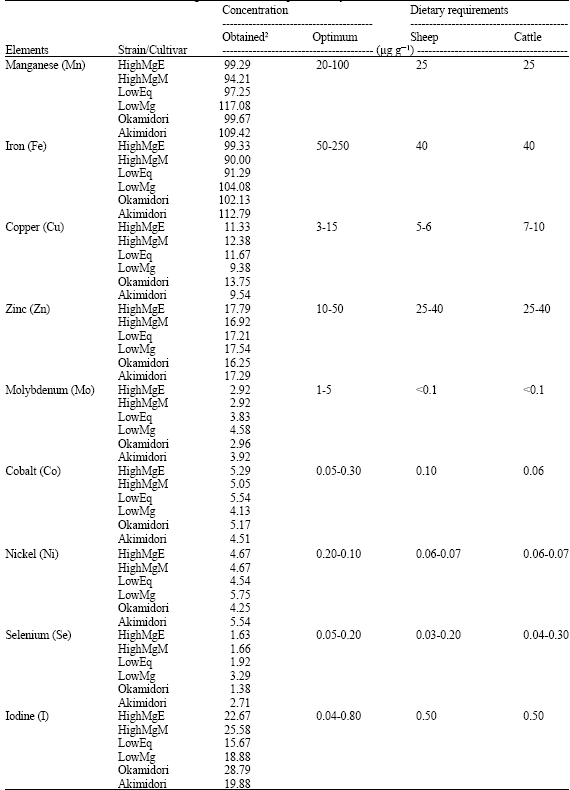 | |
| 1:Reported data from Mayland and Shewmaker (2001); 2:Average of four harvest over two years | |
The mean Fe values for all the strains/cultivars ranged from 74.56-156.00 μg g-1. Iron levels in the plants irrespective of strain/cultivar or harvest (Table 3 and 6) were optimum for plant nutrition when compared to the adequate range for grasses (Marschner, 1995; Mayland and Shewmaker, 2001). Zinc content ranged from 14.03-21.92 μg g-1 for all the harvests irrespective of strain/cultivar, which is below the critical level for dry season forage (McDowell, 1985). However, the level of Zn in the plants fell within the range reported to be adequate for cool-season grasses (Mayland and Shewmaker, 2001). The high Mg containing cultivars and strains, showed lower levels of Cu than the commercial cultivars did. This trend was very obvious in the mean harvest values. In this study, small differences in the levels of Cu were observed between the cultivars of orchardgrass. In temperate grasses, variation exists in the Cu concentration in cultivars of Lolium multiflorum, Lolium arundinaceum (formerly Festuca arundinacea) (Montalvo-Hernandez et al., 1984) and Lolium perenne (Forbes and Gilman, 1981). Apart from one instance when no differences were observed, variable results were found in different cultivars of Dactylis glomerata (Forbes and Gilman, 1981). In the 1st and 4th harvests, all the cultivars except Okamidori and LowMg and Akimidori showed critical levels of Cu that were critical (10 μg g-1) for forages (McDowell, 1985). Nevertheless, the average values of Cu from the four harvests over two years (Table 6) were within the optimum range for plant nutrition (Marschner, 1995; Mayland and Shewmaker, 2001). In the 2nd and 4th harvests, LowMg plants showed toxic levels of Mo (>5 μg g-1) Table 3 since the optimum level of Mo for cool-season grasses is 1-5 μg g-1 (Mayland and Shewmaker, 2001). Nevertheless, the average Mo levels from the four harvests over two years (Table 6) were within the optimum range for plant nutrition. The beneficial effect of Mo fertilizer under field conditions in forages is usually associated with increased N fixation. However, low temperatures could suppress N fixation and consequently lower the amount of Mo required (Anderson, 1956). Molybdenum concentration in alsike clover (Kubota et al., 1963) and alfalfa (Dionne and Pesant, 1986) grown on acid soils increased under wet conditions. Gupta and Sutcliffe (1968) found that Mo availability to plants was decreased under drier soil conditions.
Pasture herbage with minimum levels of 0.08 and 0.11 μg Co g-1 provides adequate Co for sheep and cattle, respectively (Grace, 1983; Grace and Clark, 1991). In this study, Co content of orchardgrass grown on an Andisol was found to be apparently high. However, the excess Co was not toxic to animals (Welch et al., 1991) although the level is higher than dietary requirements of Sheep and Cattle (Mayland and Shewmaker, 2001). Increase in air temperature during the growing period (Table 1) was accompanied with decrease in Ni content. Nickel concentration in excess of 7 μg g-1 dry weight is regarded as the critical toxic level in most grasses. General critical toxicity levels in crops species are in the range of >10 μg g-1 dry weight in sensitive species to >50 μg g-1 in moderately tolerant species (Bollard, 1983; Asher, 1991). The critical toxicity levels increased from 63-112 μg g-1 dry weight in wheat with increasing supply of urea (Singh et al., 1990). Increase in air temperature during the growing period (Table 1) was accompanied with increase in Se content. The same phenomenon was observed with Avena sativa forage grown in the field (Lindberg and Lannek, 1970). It can be inferred from the results that the high Mg-containing plants showed lower Se amounts. The largest Se amount was observed in LowMg in all the harvests. LowMg also showed the highest mean Se levels from the four harvests. On the other hand, the lowest mean Se level was found in Okamidori. Herbage Se concentrations of 0.03 μg g-1 are generally adequate. However, 0.10 μg g-1 may be necessary when high S reduces Se availability. In many areas of the world, grasses and forbs contain adequate (0.03-0.1 μg g-1) to toxic (>5 μg g-1) levels of Se for grazing animal (Mayland et al., 1995). Increase in rainfall during the growing period (Table 1) was accompanied with increase in I content of the grasses. The I contents of LowEq, LowMg and Akimidori fell within the required range for most common livestock (Smart and Cymbaluk, 1991). The upper limits of I in the dry matter of these cultivars were 20 μg g-1. The I content of the other cultivars was slightly above the required level in the harvests mean. However, there is a wide margin between the required level (<0.5 μg I g-1 DM) and the tolerated concentration level (<50 μg I g-1 DM) for species other than the hors (Underwood and Suttle, 1999). Iodine concentration changed with changes in the season. Apart from relative humidity, various environmental factors negatively affect I uptake by orchardgrass grown on Andisols. Iodine volatilizes under sunshine and heat. Solar radiation oxidizes iodide to I, which escape into the air (McDowell, 1992).
The mean Cu/Mo ratios in all the cultivars ranged from 2.43-4.27. Generally, the Cu/Mo ratios were below 5. Animals which consumed forages containing large amounts of Mo were affected by melybdenosis. In the presence of S, high intake of Mo can induce Cu deficiency due to the formation of insoluble Cu-Mo-S complexes in the digestive tract that reduce the absorption of Cu. Several pathways exist by which CuxMoxS interactions mitigate their effect on ruminants. The tolerable level of Cu/Mo ranges from 2:1-5:1. Risk assessment has been only partially successful because the interactions are not yet fully understood (Suttle, 1991).
The correlation coefficients show wide positive and negative relationships between the nutrients studied (Table 5). Iodine showed negative correlation with all the elements except Mg, Co and Cu. Underwood (1977) stated that iodine is antagonistic to Ca, Co and Mn. Molybdenum showed positive correlation with all the elements except Mg, Ca, Co and Cu. Copper may adversely affect the uptake of Mo (Brady, 2000b). Increases in S and Mo cause large reductions in Cu (Minson, 1990). Copper is also reduced by Fe (Campbell et al., 1974) and Zn (Dynna and Havre, 1963). Selenium showed positive correlation with most of the elements except Mg, Si, Cl, Ca, Co and Cu. Copper correlated negatively with Na, K, Mn and Ni. Cobalt correlated positively with Mg and Ca and negatively with K and M. Iron correlated positively with Na, P and Mn and negatively with Mg and Ca. Manganese and iron were antagonistic to Co absorption (Grace, 1983; Grace and Clark, 1991). Manganese correlated negatively with Mg and Ca and positively with P and K. Magnesium correlated positively with Ca and negatively with K. Vogel et al. (1989) found that Mg absorption by both plants and ruminants is negatively affected by K.
CONCLUSIONS
The variations in concentrations of micronutrients in plants are due to the stage of plant development, plant part sampled and time of sampling. Regardless of the sequence of harvest or strain/cultivar, micronutrients like Fe, Cu, Zn and Mo were at optimum levels. Other beneficial nutrients like Co, Ni, Se and I were at levels above the optimum concentrations required. This result may be due to the high levels of Co, Ni, Se and I in the soil (Andisol). High Mg-containing plants showed low levels of Se but contained high levels of Co and I. This finding implies that nutrient absorption and/or metabolism may differ genotypically. For minerals required by sheep and cattle, the mean Fe, Cu, Zn and Mo contents of orchardgrass were within the optimum range in all the cultivars. The levels of Co Ni, Se and I were above the required range. In case of Mn, only the high Mg-containing plants showed the recommended range. Present results suggest that it would be beneficial to breed orchardgrass cultivars with increased Mg without micronutrients (such as Fe, Cu, Zn and Mo) hazardous to animal nutrition are present.
REFERENCES
- Grace, N.D. and R.G. Clark, 1991. Trace Element Requirements, Diagnosis and Prevention of Deficiencies in Sheep and Cattle: Physiological Aspects of Digestion and Metabolism in Ruminants. Proceedings of the 7th International Symposium Ruminants Physiology, September 1989, Sendai, Japan, pp: 321-346.
- Kubota, J., E.R. Lemon and W.H. Allaway, 1963. The effect of soil moisture content upon the uptake of molybdenum, copper and cobalt by alsike clover. Soil Sci. Soc. Am. Proc., 27: 679-683.
CrossRef - Mayland, H.F. and D.A. Sleper, 1993. Developing a tall fescue for reduced grass tetany risk. Proceedings of the 17th International Grassland Congress, February 8-21, 1993, Palmerston North, New Zealand, pp: 1095-1096.
Direct Link - Moraghan, J.T. and H.J. Jr. Mascagni, 1991. Environmental and Soil Factors Affecting Micronutrients Deficiencies and Toxicities. In: Micronutrients in Agriculture, Mortvedt, J.J., F.R. Cox, L.M. Shuman and R.M. Welch (Eds.). Soil Sci. Soc. Am. Inc., Book Series No. 4, Madison, WI, USA., pp: 371-425.
- Suttle, N.F., 1991. The interactions between copper, molybdenum and sulphur in ruminant nutrition. Ann. Rev. Nutr., 19: 121-140.
CrossRef - Welch, R.M., W.H. Allaway, W.A. House and J. Kudata, 1991. Geographic Distribution of Trace Element Problems. In: Micronutrient in Agriculture, Mortvet, J.J., F.R. Cox, L.M. Shuman and R.M Welch (Eds.). SSSA, Madison, WI., USA., pp: 31-57.
Direct Link