
A.A. Aliero
Department of Biological Sciences, University of Usmanu Danfodiyo, Sokoto, P.M.B. 2346 Sokoto-Nigeria
International Journal of Agricultural Research
Year: 2007 | Volume: 2 | Issue: 1 | Page No.: 62-68







ABSTRACT
The differential responses of four Solanum species to seasonal dynamics in Sokoto geo-ecological zone of Nigeria were evaluated to exploit their potential for an-all-year-round cultivation. The plant height and number of leaves were markedly affected by the seasonal changes, while leaf length and width were insignificantly influenced (p>0.05). The effect of season on days to 50% flowering, fruit per branch, flower/branch was significantly affected (p<0.05). The number of fruit/plant was insignificantly affected by seasonal dynamics, a trend of horticultural importance that could be exploited for the cultivation of these species irrespective of the season. Correlation study showed significant positive and negative associations in some of the characters evaluated at 5 and 1% probability. The results of this study highlight the need for all-the-year-round cultivation of these species irrespective of the season.
PDF Abstract XML References Citation
How to cite this article
A.A. Aliero, 2007. Responses of Four Solanum Species to Seasonal Dynamics in Sokoto-Nigeria. International Journal of Agricultural Research, 2: 62-68.
DOI: 10.3923/ijar.2007.62.68
URL: https://scialert.net/abstract/?doi=ijar.2007.62.68
DOI: 10.3923/ijar.2007.62.68
URL: https://scialert.net/abstract/?doi=ijar.2007.62.68
INTRODUCTION
Eggplants and cultivated relatives cover a wide range of Solanum, with species widely distributed both in the tropics and temperate regions. Over 100 species of Solanum are known to be indigenous to Africa and several of them have been developed as vegetables in different parts of the world (Lester et al., 1986; Schippers, 2000). The best known species is S. melongena L. domesticated in the Indo-Burma region and is now cultivated worldwide. Other species cultivated that are related to S. melongena are the scarlet eggplant (S. aethiopicum L.) and Gboma eggplants (S. macrocarpon L.) cultivated mainly in Africa. Solanum macrocarpon L. is widely distributed in West Africa, Southeast Asia, Brazil and Southern Europe (Yamaguchi, 1983). In Uganda, it is a minor crop cultivated for its outstanding qualities as a vegetable due to its large leaves (Bukenya, 1994). African eggplant (Solanum aethiopicum L.), is grown as a commercial crop for domestic consumption and also for export (Daunay et al., 2001). In Ivory Coast, it is ranked second only to okra in production (Lester et al., 1990). S. americanum is another unexploited species of horticultural importance in Nigeria but is often cultivated in some localities.
Many members of the genus are have nutritional potential and are used as vegetables in many regions of the world (Asaolu and Asaolu 2002; Oboh et al., 2005). Similarly, phytochemical investigation has revealed the presence of steroidal glycoalkaloids, sesquiterpenoids and other bioactive compounds in many members of the genus which are of repute in medicine (Khan, 1979; Cipollini and Levey, 1997; Nagaoka et al., 2001; Shamim et al., 2004).
Despite the horticultural and medicinal importance of these species, their availability in this part of the world is constrained to seasonal production, largely carried out during the dry season. There is therefore, the need for an-all-year-round-production of these vegetables to meet their demand. Although, information on the effects of season on the growth and productivity of crop plants have been well documented (Siddique and Sedgley, 1986; Klapwijk, 1987; Srinivasan et al., 1998; Clarke and Siddique, 2004). However, such information on the effect of season on the growth of Solanum species in this geo-ecological region is hitherto unavailable. Such type of information is vital in understanding the differential response of these species over season as a strategy for increasing production and availability. This study was carried out to evaluate the effects of seasons on the growth and yield of four Solanum species in Sokoto geo-ecological region of Nigeria.
MATERIALS AND METHODS
The study was conducted between June-October (Rainy season) and January-April (Dry season) of 2003/2004 at the Department of Biological Sciences garden of the Usmanu Danfodiyo University, Sokoto-Nigeria which lies between longitudes 4°8' - 6°54'E and latitudes 12°'-13°58'N. The climate of Sokoto State is tropical continental dominated by two opposing air masses, tropical maritime and tropical continental. The tropical maritime is moist and blows from the Atlantic, while the tropical continental air mass is dry, blows from the Sahara Desert. Much of the rain in this area falls between June and September. Generally, the state is characterized by two extreme temperatures relative to its tropical position viz. the hot and cold seasons. The highest temperature during the hot season is experienced in the months of March/April. Between November and February, there is the prevalence of harmattan, characterized by very cold temperatures and dust laden winds, often accompanied by thick fog of alarming intensity.
Seeds of four Solanum species used were obtained from Kebbi state Agricultural supply company, Nigeria (between latitude 10°8'-13°15'N and longitude 3°30'- 6°02'E). The seeds were stored in brown paper bags at room temperature before used for experimental trials. Nursery practices and planting were done at the Biological Sciences garden of the Usmanu Danfodiyo University Sokoto. Seedlings of 6 cm height were transplanted in 10 cm diameter pots filled with soil homogenized with manure 4:1, both in dry and wet season planting. The growth conditions for the two seasons were the same, except the environmental flux. The experiment was a completely randomized design and replicated five times. At flowering, morphological parameters were evaluated. The floral and yield components were evaluated at maturity and at harvest. The data collected were analysed using SAS statistical package.
RESULTS AND DISCUSSION
The effects of seasonal variation on the growth of four Solanum species evaluated are presented in Fig. 1 and 2. The combined analysis of variance indicated that the two season had a significant effect on plant height and the number of leaves per plant (p<0.01). A comparison of the plant height dynamics shows that there were substantial differences in the plant height between seasons (Fig. 1a). In particular, the rainy season influenced plant height greatly in S. melongena and S. macrocarpon, while S. americanum and S. aethiopicum grew faster in dry season planting (Fig.1a). The temperature requirement for optimum growth and yield of aubergine has been determined to be 22-23°C and increase in temperature from 10 to 32°C resulted increased plant height in tomato and aubergine (Atherton and Harns, 1986; Uzun, 2001). In this study, the increased plant height could be associated with temperature fluctuation which can reach 42°C and 30°C low. It has been suggested that high soil temperature is more detrimental for plant growth than high air temperature (Kuroyanagi and Paulsen, 1988; Paulsen, 1994; Huang and Xu, 2000). High soil temperature alone has been reported to inhibits shoot growth (McMichael and Burke, 1994), photosynthesis and carbohydrate metabolism (Xu and Huang, 2000) and cytokinin synthesis (Udomprasert et al., 1995; Wang et al., 2003) and also induces leaf senescence (Kuroyanagi and Paulsen, 1988; Wang et al., 2003). On the other hand, leaves production were high in dry season except in S. americanum whose leaves number was found to be low.
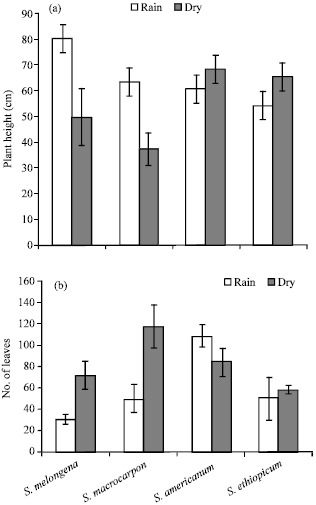 | |
| Fig. 1: | Effects of season on (a) plant height and (b) No. of leaves of Solanum species |
The decreased leaves number in this species could be attributed to its exotic nature and high transpoevaporation rate and leaf falls could have a significant effect on leaves number. However, in rainy season number of leaves increased significantly in this species (Fig. 1b). Considering the function of leaves in transpiration and CO2 assimilation (Grime, 1979; Schulze, 1986), seasonal fluctuation has been reported to affect stomatal characteristic thereby reducing transpotranspiration (Hsiao, 1973; Rodiyati et al., 2005).
The effect of season on the length of leaves was found to be insignificant (p>0.05). However, a significant interaction between the genotype and season in leaf length was observed (p<0.01). In contrast, leaf length in S. americanum and S. aethiopicum was wider during the dry season and shorter during the rain season, a trend of great significance in cultivating these species for leaf production. The leaf length in S. melongena and S. macrocarpon were comparatively high in dry season (Fig. 2a). The effect of season on the leaf width was insignificantly affected by seasonal fluctuation (p>0.05). However, leaf width was found to be broader in S. melongena and S. macrocarpon in rainy season, while in S. americanum and S. aethiopicum the leaf width were broader dry season (Fig. 2b). A significant interaction between the species and season was found for the plant height, number of leaves, leaves/plant, leaflet length and width. A significant for interaction between species and season implies that there is a differential response over season. A similar trend on the growth of Abelmoschus esculentus cultivars in South-western Nigeria has been reported (Morakinyo and Makinde, 1991).
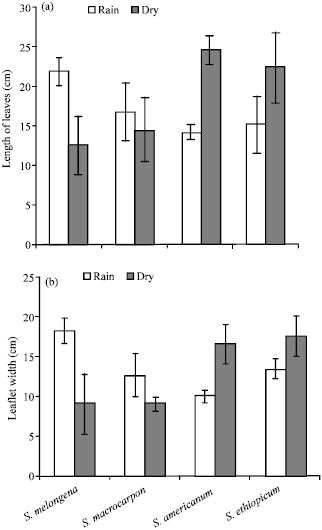 | |
| Fig. 2: | Effects of season on (a) Length of leaves and (b) leaf width of Solanum species |
The floral and yield components evaluated are presented (Table 1). The effect of season on the number of days to 50% flowering and fruits per branch were significantly different at p<0.05. It is of interest to note that, the number of fruit per plant and number of flower per plant were insignificantly affected by the seasonal variation (p>0.05). However, a closer look at the result (Table 1), the number of fruits per plant was high in dry season, which support traditional horticultural practices of these species in the dry season. The decrease in the number of days to flowering in rainy season is however, noteworthy. In most of the species, flowers were produced 10 days earlier than the dry season. It is possible that early flowering in the rainy season could be in response to average temperature due to the moist tropical maritime which blows from the Atlantic Ocean. According to Thomas and Raper (1981), temperature has a very significant effect on time of anthesis and low temperature has been reported to delay flowering in Soybean (Wang et al., 1997). High temperatures enhance vegetative production but suppress reproductive activity such as flowering, resulting in lower yield (Nkansah, 2001). Photosynthesis has been reported to increase with increasing temperature (Augustine et al., 1976; Nkansah and Ito, 1994).
The interrelationship of the variables evaluated as affected by seasonal changes is presented in Table 2. Highly significant correlations were determined between the parameters studied at 5% and 1% probability. The significant correlation between plant height and leaf length is expected, as plant grew taller its leaf length increase.
| Table 1: | Mean±SD for the floral and yield parameters evaluated |
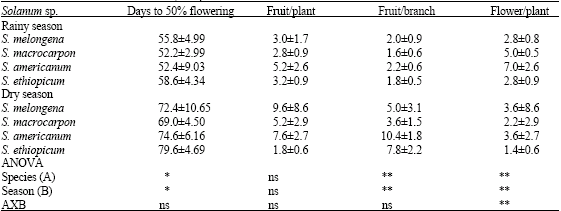 | |
| Values are mean±SD (n = 5). Ns, *, ** = not significant at 5 and 1% probability levels, respectively | |
| Table 2: | Correlation for the eight variables evaluated |
 | |
| *, ** = Significant at 5 and 1% probability level | |
A significant positive and negative correlation implies the degree of relatedness of a character on each other as affected by seasonal fluctuation. The concept of character association has been emphasized by Mather and Jinks (1992) and it is a feature of total phenotype which is of significance in selection. The presence of variability as influence by genotype and or environment could be attributed to pleiotrophy. Phenotype and environmental correlations have been reported Aliero (2000) and Morakinyo (1996).
CONCLUSIONS
Although horticultural practices of these vegetables are carried out traditionally in dry season, the result of this study indicates that these species possesses potential for being cultivated in rainy season. It was observed that plants grown in the rainy season exhibited a lower fruit yield and growth than those grown in the dry season; however, the level of variation was not substantial enough to hamper all-year-round cultivation. This study highlights on the need for the cultivation of these species in order to augment their demand during the rainy period.
REFERENCES
- Asaolu, M.F. and S.S. Asaolu, 2002. Proximate and mineral composition of cooked and uncooked Solanum melongena. Int. J. Food Sci. Nutr., 53: 103-107.
CrossRefDirect Link - Atherton, J.G. and G.P. Harris, 1986. Flowering. In: The Tomato Crop: A Scientific Basis for Improvement, Atherton, J.G. and J. Rudich (Eds.), Springer, Dordrecht, Netherlands, ISBN-13: 978-94-009-3137-4, pp: 167-200.
CrossRefDirect Link - Cipollini, M.L. and D.J. Levey, 1997. Antifungal activity of Solanum fruit glycoalkaloids: Implications for frugivory and seed dispersal. Ecology, 78: 799-809.
Direct Link - Clarke, H.J. and K.H.M. Siddique, 2004. Response of chickpea genotypes to low temperature stress during reproductive development. Field Crop. Res., 90: 323-334.
Direct Link - Hsiao, T.C., 1973. Plant responses to water stress. Annu. Rev. Plant Physiol., 24: 519-570.
CrossRefDirect Link - Huang, B. and Q. Xu, 2000. Root growth and nutrient element status of creeping bentgrass cultivars differing in heat tolerance as influenced by supraoptimal shoot and root temperatures. J. Plant Nutr., 23: 979-990.
Direct Link - Lester, R.N., P.M.L. Jaeger, B.H.M. Bleijendaal-Spierings, H.P.O. Bleijendaal and H.L.O. Holloway, 1990. African eggplants-a review of collecting in West Africa. Plant Genet. Resour. Newsl., 81-82: 17-26.
Direct Link - Morakinyo, J.A. and S.C. Makinde, 1991. Variability and heritability in local cultivars of okra (Abelmoscus esculentus (L.). Moench). Nig. J. Bot., 4: 33-40.
Direct Link - McMichael, B.L. and J.J. Burke, 1994. Metabolic activity of cotton roots in response to temperature. Environ. Exp. Bot., 34: 201-206.
Direct Link - Nagaokaa, T., K. Goto, A. Watanabe, Y. Sakata and T. Yoshihara, 2001. Sesquiterpenoids in root exudates of Solanum aethiopicum. Z Naturforsch, 56c: 707-713.
Direct Link - Nkansah, G.O., 2001. Some physiological features of the African eggplant, Solanum aethiopicum group >Gilo=. Sci. Hortic., 90: 181-186.
Direct Link - Oboh, G., M.M. Ekperigin and M.I. Kazeem, 2005. Nutritional and haemolytic properties of eggplants (Solanum macrocarpon) leaves. J. Food Compos. Anal., 18: 153-160.
CrossRefDirect Link - Rodiyati, A., E. Arisoesilaningsih, Y. Isagi and N. Nakagoshi, 2005. Responses of Cyperus brevifolius (Rottb.) Hassk. and Cyperus kyllingia Endl. to varying soil water availability. Environ. Exp. Bot., 53: 259-269.
Direct Link - Schippers, R.R., 2000. African Indigenous Vegetables an Overview of Cultivated Species. Natural Resources Institute, Chatham, UK., pp: 89-98.
Direct Link - Shamim, S., S.W. Ahmed and I. Azhar, 2004. Antifungal activity of Allium, Aloe and Solanum species. Pharm. Biol., 42: 491-498.
Direct Link - Srinivasan, A., C. Johansen and N.P. Saxena, 1998. Cold tolerance during early reproductive growth of chickpea (Cicer arietinum L.): Characterization of stress and genetic variation in pod set. Field Crops Res., 57: 181-193.
Direct Link - Udomprasert, N., P.H. Li, D.V. Davis and A.H. Markhart III, 1995. Root cytokinin level in relation to heat tolerance of Phaseolus acutifolius and Phaseolus vulgaris. Crop Sci., 35: 486-490.
Direct Link - Wang, Z., V.R. Reddy and B. Quebedeaux, 1997. Growth and photosynthetic responses of soybean to short-term cold temperature. Environ. Exp. Bot., 37: 13-24.
Direct Link - Wang, Z., J. Pote and B. Huang, 2003. Responses of cytokinins, antioxidant enzymes and lipid peroxidation in shoots of creeping bentgrass to high root-zone temperatures. J. Am. Soc. Hortic. Sci., 128: 648-655.
Direct Link - Xu, Q. and B. Huang, 2000. Growth and physiological responses of creeping bentgrass to changes in air and soil temperatures. Crop Sci., 40: 1363-1368.
Direct Link