
B. Baninasab
Department of Horticulture, College of Agriculture, Shiraz University, Shiraz, Iran
M. Rahemi
Department of Horticulture, College of Agriculture, Shiraz University, Shiraz, Iran
B. Kholdebarin
Department of Biology, College of Science, Shiraz University, Shiraz, Iran
International Journal of Agricultural Research
Year: 2007 | Volume: 2 | Issue: 1 | Page No.: 43-52







ABSTRACT
The effects of photosynthetate demand on photosynthetic activity of pistachio (Pistacia vera L.) trees cv. Owhadi, which are characterized by strong alternate bearing were studied. Changes in seasonal leaf gas exchange, the amount of chlorophyll, leaves dry weight, leaf area and leaf specific weight in fruit bearing branches of ON trees and non-fruit bearing branches of OFF trees were determined. Rate of photosynthesis, stomatal conductance, transpiration rate, leaf mesophyll efficiency of ON trees measured forty and seventy Days After Full Blooming (DAFB) were significantly higher than those of OFF trees. Seventy DAFB, rates of photosynthesis, stomatal conductance, transpiration rates and leaf mesophill efficiency decreased in the two three groups, so that after this period there was no significant difference between ON and OFF trees with respect to these parameters. There was a decrease in internal CO2 concentration with the increase in photosynthetic rates. The amount of chlorophyll was higher in ON trees. However, the differences were only significant hundred and hundred thirty DAFB. Leaves on tress were smaller and had less weight than OFF trees. Leaf specific weights of OFF trees at hundred thirty and hundred sixty DAFB were significantly higher than that of ON trees.
PDF Abstract XML References Citation
How to cite this article
B. Baninasab, M. Rahemi and B. Kholdebarin, 2007. Seasonal Variations in Photosynthetic Activities of Pistachio Trees: A Comparison Between Fruiting (ON) and Non-Fruiting (OFF) Trees. International Journal of Agricultural Research, 2: 43-52.
DOI: 10.3923/ijar.2007.43.52
URL: https://scialert.net/abstract/?doi=ijar.2007.43.52
DOI: 10.3923/ijar.2007.43.52
URL: https://scialert.net/abstract/?doi=ijar.2007.43.52
INTRODUCTION
Pistachio (Pistacia vera L.) is a deciduous tree with a very strong alternate bearing character. Its yield on years is 3-5 times greater than on OFF years (Johnson and Weinbaum, 1987). Although a great amount of studies have been made on pistachio tree alternate bearing, its physiological mechanism is still not clearly understood. Most studies are emphasizing on the roles of carbohydrates and inorganic nutrients in alternate bearing and pistachio flower bud abscission (Brown et al., 1995; Vemmos, 1999). However, not enough information is available on the relations between leaf gas exchange variables and alternate bearing and pistachio fruits growth and development. In most plants the rate of fruit growth and development as a sink, affects the rate of photosynthesis. Clearly this effect depends on the magnitude of sink demand. Some studies have indicated the absence of any correlation between rate of photosynthesis and sink strength. In chest nut (Proietti et al., 2000) and citrus (Syvertsen et al., 2003) fruits are strong sinks for photosynthates and cause an increase in photosynthesis in leaf next to them. Schaffer et al. (1986) have shown that the effects of fruits on photosynthesis in strawberry leaves were variable. The effects were an increase in some cases and a decrease in others. Contrary to above reports, fruits effects on leaf photosynthesis were not observed in apple (Rom and Ferree, 1986) and in olive (Proietti, 2000) trees.
In fruit trees changes in photosynthetic components with respect to alternate bearing have seldom been studied. Some studies have shown that whereas in ON and OFF olive trees the stomatal conductance and transpiration rates were similar (Proietti, 2000), these parameters were higher in ON orange trees than in OFF ones (Syvertsen et al., 2003). Fruits removal has also caused an increase in leaf internal CO2 concentration (Schaffer et al., 1987) and a decrease in chlorophyll per unit leaf area (Nii et al., 1995) without a change in chlorophyll “a” to “b” ratios (Nii et al., 1995). The effects of fruits on morphology of trees leaves have also been studied. Some studies have shown an increase in leaf area and leaf specific weight after fruit removal (Schaffer et al., 1986; Nii et al., 1995). Considering the above findings, one can support the hypothesis that the fruit trees need for carbohydrates at the time of fruit removal play an important role in leaf photosynthetic activity as has been proposed by Ferre and Palmer (1982).
The goal of present research is to study changes taking place in ON and OFF pistachio tree leaves without interfering with the natural status of trees i.e., partial or total fruit removal which could have secondary effects on trees performance (Daie, 1985). In present study, we attempt to verify the hypothesis that in pistachio trees like in other fruit trees, the presence of fruit as a sink would cause an increase in photosynthetic rates.
MATERIALS AND METHODS
Experiments were carried out in central part of Iran (Kerman Province, 30°15´ N and 56°58´ E) under natural conditions using 30 years old pistachio trees cv. Owhadi. Upper leaflets of the leaves present in the middle of branches bearing similar amount of fruits from 8 ON and branches without fruit from 8 OFF trees were used for experimental purposes. Measurements were made monthly from April 16, ten days after full bloom (DAFB) till September 13 (160 DAFB) which was the harvest time.
Portable infrared gas analyzer (ADC, LCA-4, Analytical Development Co., Hoddesdon, UK) and Parkinson leaf chamber (PLC4) were used to measure photosynthesis, stomatal conductance, transpiration and leaf internal CO2 concentration. Measurements were made between 9 and 11 AM in the orchard. To determine the above parameters, the upper leaflets of each leaf were placed in leaf chamber and were exposed to direct sunlight under field conditions. A quantum sensor attached to leaf chamber was used to measure the Photosynthetic Photon Flux Density (PPFD) the magnitude of which was more 1600 μmole m-2 s-1 throughout the experiments. The temperature inside the leaf chamber was 1-3°C higher than the ambient temperature in all experiments, the values of which for 10, 40, 70, 100, 130 and 160 DAFB were 26-29, 30-36, 36-40, 33-35, 32-36 and 27-31°C, respectively. The ambient relative humidity for the first and last measurements was 32±3% and for the remaining was 24±2%. The two parameters, WUE and leaf mesophyll efficiency were determined by ratio of photosynthesis over transpiration and by the ratio of photosynthesis over internal CO2 concentration, respectively. Chlorophyll content was determined by spectrophotometer (Saini et al., 2001).
Planimeter (ADC, AM200, Analytical Development Co., Hoddesdon, U.K) was used to measure leaf area. After this, leaves were placed in an oven for 48 h set at 70°C after which the leaves dry weight was determined. Using the ratio of dry weight over leaf area, the leaf specific weight was calculated.
The experimental design was a complete randomized block with 4 replicates and each replicate was composed of 2 trees. MSTAT-C soft ware was used to analyze the data. The lowest significant difference test (LSD) was used to compare the means. Excel soft ware was used to determine the linear relation between photosynthesis and other parameters (stomatal conductance, transpiration, internal CO2 concentration and chlorophyll content).
RESULTS
Data shown in Table 1 indicate that the state of pistachio trees as being ON or OFF with respect to fruit production has a significant effect on photosynthesis, stomatal conductance, transpiration and leaf mesophyll efficiency.
| Table 1: | Statistical significance of effects of fruiting state of tree (OFF or ON) and time of measuring, as a factorial experiment, on different variables in leaf of pistachio trees |
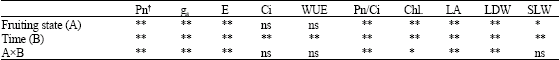 | |
| * p<0.05, ** p<0.01, ns = Not significant, † Pn = Photosynthesis rate, gs = Stomatal conductance, E = Transpiration rate, Ci = Internal CO2 concentration, WUE = Water Use Efficiency, Pn/Ci = Mesophyll efficiency, Chl. = Chlorophyll content, LA = Leaf Area, LDW = Leaf Dry Weight, SLW = Specific Leaf Weight | |
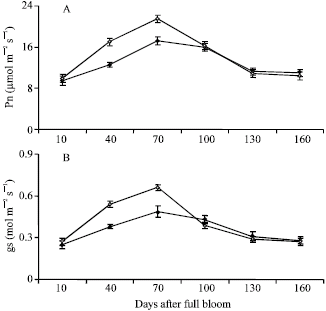 | |
| Fig. 1: | Seasonal changes in the (A) net photosynthesis (Pn) and (B) stomatal conductance (gs) in leaf of OFF (closed symbols) and ON (opened symbols) of pistachio trees. The vertical bars indicate±SE |
Date of measurements and samplings throughout growth season has also a significant effect on all leaf photosynthetic parameters. Results also showed that for parameters such as internal CO2 concentration and water use efficiency, there was no interaction effect between tree status with respect to yield and date of measurements throughout growth season (Table 1).
Analyzing the trend in seasonal variation of photosynthesis shared that at the beginning of season the rates of photosynthesis of ON and OFF trees were similar. There after till 70 DAFB the rate of photosynthesis increased in both groups, however, the increase in ON trees was significantly higher than OFF trees. At 40 and 70 DAFB the rates of photosynthesis in ON trees were 34.7 and 24.8% higher than OFF trees, respectively (Fig. 1A). From 70 DAFB till harvest time, the rates of photosynthesis in both ON and OFF trees were almost the same.
Stomatal conductance in OFF trees, increased continuously till 70 DAFB. At this time, its value was 0.49 mol m-2 s-1 and from then on started to decrease and reached the value of 0.28 mol m-2 s-1 at harvest time (Fig. 1B). In ON trees, stomatal conductance was higher than OFF trees till 70 DAFB and from then on it decreased at a faster rate than OFF trees and at the time of harvest reached a value of 0.27 mol m-2 s-1 (Fig. 1B).
At the beginning of growth season, there was no significant difference in the rate of transpiration in the two groups. However, after this stage till 70 DAFB the rate of transpiration in ON trees was significantly higher than in OFF trees. At this time, the rate of transpiration in ON trees was 29.3% higher than OFF trees (Fig. 2A).
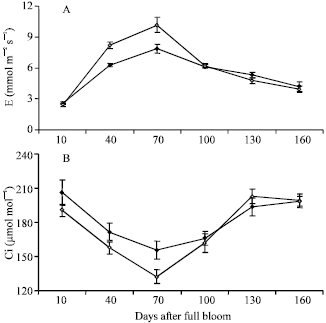 | |
| Fig. 2: | Seasonal changes in the (A) transpiration rate (E) and (B) internal CO2 concentration (Ci) in leaf of OFF (closed symbols) and ON (opened symbols) of pistachio trees. The vertical bars indicate±SE |
The trend in seasonal change in internal CO2 concentration in the two tree groups was opposite to that of photosynthesis i.e., with increase in the rate of photosynthesis, the internal CO2 concentration decreased and vise versa (Fig. 1A and 2B). It was also revealed that till 100DAFB the internal CO2 concentration in OFF trees was more than the amount in ON trees. However, after 100 DAFB, the internal CO2 concentration in ON trees was slightly higher than OFF trees, but the difference was not significant (Fig. 2B).
The seasonal change in leaf mesophyll efficiency was similar to that of photosynthesis, i.e., from the beginning of growth season till 70 DAFB there was an increase in leaf mesophyll efficiency in both groups and from then on it started to decrease (Fig. 3A). At 40 and 70 DAFB, the leaf mesophyll efficiency was significantly higher in ON trees (Fig. 3A).
Figure 3B shows that the water use efficiency (WUE) decreases rapidly at the beginning of growth season and from then on it stays constant in both ON and OFF trees till the end of season.
The relation between the amount of chlorophyll, leaf area, dry weight and specific weight on one hand and the amount of fruit yield on the other was significant (Table 1). Results presented in Table 2 show that the amount of chlorophyll present in the leaves of the two tree groups increases till 100 DAFB and there after, it decreases. The comparison of the means shows that only at 70 and 100 DAFB the chlorophyll concentration in leaves of ON trees is significantly higher than that of OFF trees. At the beginning of growth season (10 DAFB), the leaf dry weight of the two groups was similar. After this period till 70 DAFB the leaf dry weight increased. The increase in OFF trees was significantly higher than ON trees. These results also reveal that in the period between 70 DAFB till harvest time the leaf dry weight of ON trees decreases somehow (Table 2). Although the leaf area of ON trees at 10 DAFB was higher than that of OFF trees, at the end of growth season the leaf area (74.11 cm2) of ON trees was 44.7% smaller than that of (107.24 cm2) OFF trees (Table 2). Results also show that leaf specific weights of both OFF and ON trees till 100 DAFB were similar and after this period till the end of the experiments, their amounts in OFF trees were significantly higher than in ON trees (Table 2).
| Table 2: | Seasonal patterns of leaf chlorophyll (chl.) content, Leaf Dry Weight (LDW), leaf Area (LA) and Specific Leaf Weight (SLW) in OFF and ON pistachio trees |
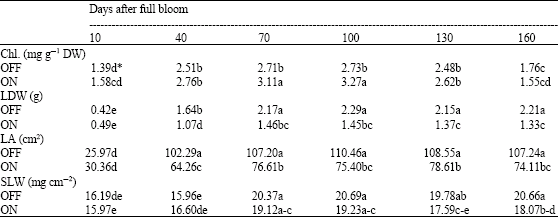 | |
| * Mean separation within columns and rows by LSD, at 5% level | |
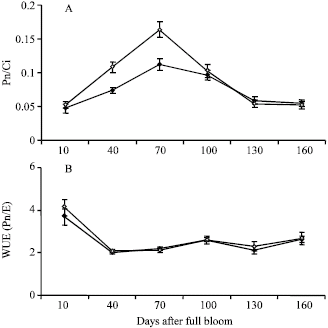 | |
| Fig. 3: | Seasonal changes in the (A) mesophyll efficiency (Pn/Ci) and (B) water use efficiency (WUE) in leaf of OFF (closed symbols) and ON (opened symbols) of pistachio trees. The vertical bars indicate±SE |
DISCUSSION
Results of this investigation show the significant effects of fruits in pistachio trees on the rate of photosynthetic CO2 fixation under field condition in central parts of Iran until 70 DAFB. Part of these enhancing fruits effects on photosynthesis could be due to an increase in the demand for photosynthetic products by growing fruits. The needs of plant organs which act as sinks for such products can regulate the rate of photosynthesis (Ho, 1992). There are numerous reports indicating that the increase in sink demand in plants will result in both an increase in the rate of photosynthetic CO2 fixation and the rate of photosynthetate export from leaves and vise versa (Geiger, 1976; Hall and ilthrope, 1978; Peet and Kramer, 1980; Tekalign and Hammes, 2005). Pammenter et al. (1993) have also proposed that low in sink demand will result in an accumulation of photosynthetic products in leaves causing a decrease in the rate of photosynthesis. Other aspects of fruit effects on photosynthesis could be due to growth regulators effects (Lenz, 1979). Since young and developing fruits are rich in growth regulators (Luckwill, 1975), one can expect that these hormones to regulate genes expression responsible for the synthesis of key photosynthetic CO2 fixation enzymes such as ribulose-1,5-bisphosphate carboxylase/oxygenase (RUBISCO) (Wareing, 1968). There are reports in support of the enhancing effects of auxin, cytokinin and gibberellin on RUBISCO activity in leaves (Huber and Sankhla, 1973). Tamas et al. (1972) have also reported that IAA increases photosynthetic efficiency by increasing photophosphorylation. Hoad et al. (1977) found that in grape plants, removing fruits first resulted in a decline in GA and cytokinin concentrations followed by a decrease in the rate of photosynthesis. However, in present study we found that the fruit effect on pistachio tree photosynthesis was not as great as it has been reported for strawberry (Forney and Breen, 1985) and for orange (Syvertsen et al., 2003). Present results show a decline in photosynthesis in both ON and OFF trees in the middle of growing season which could be due to natural senescence of pistachio leaves. Similar decline in photosynthesis by aging leaves has been reported for apple trees (Watson and Landsberg, 1979) and for olive trees (Proietti, 2000). It has also been reported that the decrease in leaf photosynthesis in summer could be due to temperature damage to photosystems and to an increase in the rate of photorespiration (Beyschlag et al., 1987; Angelopoulos et al., 1996). Also the faster and earlier leaf senescence of ON trees could be the reason for the similarity of photosynthetic rates in both ON and OFF trees in the middle of growth season. In present study, the maximum values of photosynthetic rates coincided with the values which have been reported for P. atalantica and P. vera (Lin et al., 1984).
Our results show that the presence of fruit has a significant effect on pistachio leaf stomatal conductance and transpiration. Also the enhancing effect of fruits on photosynthesis could be due to an increase in both stomatal conductance and leaf transpiration. As shown by Fig. 4A and B, there is a close and positive correlation (r = 0.97) between photosynthesis and stomatal conductance and also between photosynthesis and transpiration (r = 0.88). Such a positive correlations have also been reported by others (Proctor, 1981; Flexas et al., 2001) for some trees. It is not exactly known as how fruits control stomatal conductance and transpiration rate. However, some researchers propose the hypothesis that the decrease in ABA level by growing fruits cause an increase in stomatal conductance followed by an increase in transpiration. The role of ABA in stomatal movement has been very well documented (Salisbury and Ross, 1991). Luckwill (1975) has also reported that leaves at the proximity of growing fruits have less ABA and thus more stomatal conductance than the more distal ones. Loveys and Kriedmann (1974) studied a large number of plants and showed that fruit removal causes an increase in leaves ABA level.
The increase in pistachio leaf photosynthesis was accompanied by a decrease in internal CO2 pressure and an increase in leaf mesophyll efficiency. Present results and as well as those reported by Flexas et al. (2001) show that in pistachio, photosynthesis regulation is affected by both stomatal and non-stomatal factors. Our results also show a negative correlation (r = 0.92) between photosynthesis and leaf internal CO2 pressure (Fig. 4C). Similar results have been reported by Proieti (2000) for olive trees.
Water use efficiency by OFF and ON trees at 40 DAFB, was reduced by 45.3 and 49.5%, respectively (Fig. 3B). These reductions were probably due to a higher increase in the rate of transpiration compared to photosynthesis (Fig. 1A and 2B). In our study, the fruit bearing status of trees did not have any effects on WUE. Similar results have been reported by Novello and Palma (1995) for pistachio trees cv. Bianca.
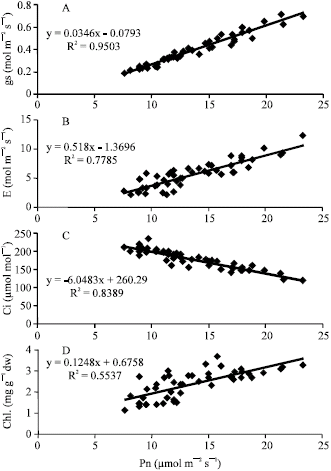 | |
| Fig. 4: | Relationship between net photosynthesis (Pn) and (A) stomatal conductance (gs), (B) transpiration rate (E), © internal CO2 concentrations (Ci) and (D) chlorophyll content (Chl.) of pistachio leaf |
The leaf chlorophyll content was also affected by trees alternate bearing state (Table 1 and 2). Present results along with those of Baninasab and Rahemi (2006) indicate that the higher chlorophyll content of ON trees is partially due to the presence of more fruits on trees and probably also due to carbohydrate accumulation in the leaves of OFF trees. In our Investigation a positive correlation (r = 0.74) between the rate of photosynthesis and the level of chlorophyll in the leaves was observed (Fig. 4D). Similar findings have been reported by Schaffer et al. (1986) for strawberry.
Our results also show that ON trees at the beginning of growth season have had enough nutrient reserve for leaf emergence and expansion, but later on due to competition between leaves and growing fruits, they have reached maturity and senescence stages faster (Table 2). On the other hand, slower growth rate and smaller leaf size of OFF trees at the beginning of growth season could be due to the fact that these trees have been on cycle in the previous year and have been exhausted of their reserve materials. Newell (1987) has also shown that branches of Aesculus california Spach that were bearing fruits in the previous year, produced smaller leaves in the following year. Similar observations have been reported for cherry (Kappel, 1991) and for olive (Rallo and Suarez, 1989) trees.
In the present study the leaf specific weight of ON trees was smaller than that of OFF trees. This could be due to the higher rates of photosynthetate export from ON trees leaving less structural and non-structural materials in the leaves (Forney and Breen, 1985; Wooge and Barden, 1987; Thomas, 1990). ON trees had leaves with less dry weight than OFF trees (Table 2), which again results from higher rate of photosynthetates export from ON trees leaves to supply materials needed for growing fruits and seeds. At 70 and 130 DAFB, the weight of ON trees leaves was reduced by 0.13 g which may be due to higher rates of carbohydrates (Nzima, 1994) and nitrogenous compounds (Weinbaum et al., 1994) export from these leaves.
Results presented in this investigation point to the effects of source-sink relationships on photosynthesis in pistachio trees. Pistachio trees on cycle had higher rates of photosynthesis, stomatal conductance, transpiration and leaf mesophyll efficiency till 70 DAFB than those of OFF trees for the same period. However, after this period, because of faster aging process in ON trees leaves, there was no significant difference with respect to the above parameters between the two tree groups.
ACKNOWLEDGMENTS
The generous support and assistance of the Iranian Pistachio Research Institute is greatly appreciated. We also like to thank Drs. A. Javanshah and B. Panahi for their scientific inputs in this study. The technical help received from H. Alipour and A. Ershadi is acknowledged.
REFERENCES
- Angelopoulos, K., B. Dichio and C. Xiloyannis, 1996. Inhibition of photosynthesis in olive trees (Olea europaea L.) during water stress and rewatering. J. Exp. Bot., 47: 1093-1100.
CrossRefDirect Link - Beyschlag, W., O.L. Lange and J.D. Tenhunen, 1987. Diurnal Patterns of Leaf Internal CO2 Partial Pressure of the Sclerophyll Shrub Arbutus Unedo Growing in Portugul. In: Plant Response to Stress, Tenhunen, J.D., F.M. Catarino, O.L. Lange and W.C. Oechel (Eds.). Springer-Verlag, Berlin, Heidelberg, New York, London, Paris, Tokyo, pp: 355-368.
- Brown, P.H., S.A. Weinbaum and G.A. Picchioni, 1995. Alternate bearing influences annual nutrient consumption and total nutrient content of mature pistachio trees. Trees, 9: 158-164.
Direct Link - Flexas, J., J. Gulias, S. Jonasson, H. Medrano and N. Mus, 2001. Seasonal patterns and control of gas exchange in local populations of the mediterranean evergreen shrub Pistacia lentiscus L. Acta Oecol., 22: 33-43.
Direct Link - Kappel, F., 1991. Partitioning of above-ground dry matter in Lambart sweet cherry trees with and without fruit. J. Am. Soc. Hortic. Sci., 116: 201-205.
Direct Link - Lin, T.S., J.C. Crane, K. Ryugo, V.S. Polito and T.M. Dejong, 1984. Comparative study of leaf morphology, photosynthesis and leaf conductance in selected Pistachio species. J. Am. Soc. Hortic. Sci., 109: 325-330.
Direct Link - Novello, V. and L. Palma, 1995. Observation on the pistachio photosynthetic activity in southern Italy. Acta Hortic., 419: 97-102.
Direct Link - Pammenter, N.W., F. Loreto and T.D. Sharkey, 1993. End product feedback effect on photosynthetic electron transport. Photosyn. Res., 35: 5-14.
Direct Link - Proietti, P., 2001. Effect of fruiting on leaf gas exchange in olive (Olea europaea L.). Photosynthetica, 38: 396-402.
Direct Link - Proietti, P., A. Palliotti, F. Famiani, E. Antognozzi, F. Ferranti, R. Andreutti and G. Frenguelli, 2000. Influence of leaf position, fruit and light availability on photosynthesis of two chestnut genotypes. Sci. Hortic., 85: 63-73.
Direct Link - Syvertsen, J.P., C. Goni and A. Otero, 2003. Fruit load and canopy shading affect leaf characteristics and net gas exchange of spring navel orange trees. Tree Physiol., 23: 899-906.
Direct Link - Tekalign, T. and P.S. Hammes, 2005. Growth and productivity of potato as influenced by cultivar and reproductive growth. I. Stomatal conductance, rate of transpiration, net photosynthesis and dry matter production and allocation. Sci. Hortic., 105: 13-27.
Direct Link - Thomas, H., 1990. Leaf development in Lolium temulentum: Proteing metabolism during development and senescence of attached and excised leaf tissue. J. Plant Physiol., 136: 45-50.
Direct Link - Vemmos, S.N., 1999. Carbohydrate content of inflorescent buds of defruited and fruiting pistachio (Pistacia vera L.) branches in relation to biennial bearing. J. Hortic. Sci. Biol., 74: 94-100.
Direct Link - Weinbaum, S.A., G.A. Picchioni, T.T. Muraoka, L. Ferguson and P.H. Brown, 1994. Fertilizer nitrogen and boron uptake, storage and allocation vary during the alternate bearing cycle in pistachio trees. J. Jap. Soc. Hortic. Sci., 119: 24-31.
Direct Link