
Radhouane Chaffai
Laboratoire de Biologie et Physiologie Cellulaires V6g6tales, Faculte des Sciences de Bizerte, 7021 Zarzmma, Tunisia
Mohamed Anis Elhammadi
Laboratoire de Biologie et Physiologie Cellulaires V6g6tales, Faculte des Sciences de Bizerte, 7021 Zarzmma, Tunisia
Tini Nouhou Seybou
Laboratoire de Biologie et Physiologie Cellulaires V6g6tales, Faculte des Sciences de Bizerte, 7021 Zarzmma, Tunisia
Ali Tekitek
Laboratoire de Biochimie et de Biologie Mol6culaire,
Faculte des Sciences de Bizerte, 7021 Zarzmma, Tunisie
Brahim Marzouk
Laboratoire des Ameliorations et Adaptations des Plantes, Institut National de la Recherche Scientifique et Technique (INRST), BP 95-2050 Harnmarn-Lif, Tunisie
Ezzedine El Ferjani
Laboratoire de Biologie et Physiologie Cellulaires V6g6tales, Faculte des Sciences de Bizerte, 7021 Zarzmma, Tunisia
International Journal of Agricultural Research
Year: 2007 | Volume: 2 | Issue: 2 | Page No.: 126-135







ABSTRACT
The fatty acid composition and the polar lipid profiles are examined in maize (Zea mays L., cv. Alistrong) treated with 100 μM Cd. In roots, analysis of fatty acid composition reveals a general pattern of increase in the unsaturation level of most polar lipid classes, as indicated by the Double Bond Index (DBI). The results also include no alteration of total fatty acid, phospholipid (PL) and Steryl Lipid (SL) contents, but glycolipids (GL) were substantially increased. In shoots, the double bond indices of phosphatidylcholine (PC) and phosphatidylglycerol (PG) were decreased, whereas those obtained from MGDG, DGDG and SQDG showed an opposite response. The PG and MGDG content in shoots were markedly decreased by about 78 and 65%, respectively, indicating that the Cd toxicity caused severe damage to the structure and function of photosynthetic membranes. The increase of the malondialdehyde (MDA) content in roots indicates an oxidative stress, which can be involved in mediating compositional membrane alterations.
PDF Abstract XML References
How to cite this article
Radhouane Chaffai, Mohamed Anis Elhammadi, Tini Nouhou Seybou, Ali Tekitek, Brahim Marzouk and Ezzedine El Ferjani, 2007. Changes in Polar Lipid Composition in Maize Seedlings Induced by Cadmium Stress. International Journal of Agricultural Research, 2: 126-135.
DOI: 10.3923/ijar.2007.126.135
URL: https://scialert.net/abstract/?doi=ijar.2007.126.135
DOI: 10.3923/ijar.2007.126.135
URL: https://scialert.net/abstract/?doi=ijar.2007.126.135
INTRODUCTION
One of the most prevalent environmental challenges encountered by plants is the exposure to heavy metals, such as Cd, Hg and As which have been shown to affect plant growth and development in contaminated sites. Many heavy metals are essential micronutrients, whereas others such as Cd have no known function as nutrient. It has been estimated that the anthropogenic emissions of Cd are in the range of 30, 000 t per year (Sanità di Toppi et al., 1999). In unpolluted soils, Cd is present at concentrations of 0.1-0.5 mg kg-1 but in heavily polluted soils of sewage sludge, concentrations of up to 150 mg kg-1 have been found (Jackson and Alloway, 1991).
According to their chemical and physical properties, three different molecular mechanisms of metal toxicity can be distinguished: (i) production of reactive oxygen species by autoxidation and Fenton reaction, (ii) blocking of essential functional groups in biomolecules and (iii) displacement of essential metal ions from biomolecules (Schützendübel and Polle, 2002). Binding and/or interaction of heavy metals with plasma membrane may lead to an overproduction of free radicals, H2O2 and hydroxyl ions and finally to membrane damage (Gallego et al., 1996) by oxidizing proteins and phospholipids (Mehlhorn et al., 1986). Cadmium can generate reactive oxygen species, inducing functional alterations of membranes (Fodor et al., 1995).
Plants respond to heavy metal toxicity in a variety of different ways, by the induction of general stress responses, which in part include, the immediate changes in the membrane lipid composition (Jones and Harwood, 1993). However, regulation of fatty acid biosynthesis was considered to be one mean of controlling the membrane fatty acid unsaturation that could contribute to the stress tolerance. However, the question whether changes in fatty acid composition and unsaturation, play a role in the defense mechanism, or rather reflect damage remains poorly understood and a subject of controversy. This question has been addressed in many reports by exploiting mutants deficient in lipid fatty acid desaturation and in transgenic plants over expressing fatty acid desaturases, in response to various abiotic stresses, including heat, salt and drought (Zhang et al., 2005). Studies with plants suggested that the majority of them respond to cadmium by decreasing the level of unsaturated fatty acids (Ouariti et al., 1997).
In this study, we examined the changes in the fatty acid profile of maize plants grown under cadmium stress. The significance of such changes in providing protection against cadmium will be also discussed.
MATERIALS AND METHODS
Growth Conditions
The corn (Zea mays L. cv. Alistrong) seeds were surface-sterilized with 10% (v/v) H2O2 for 20 min, rinsed thoroughly with tap water and germinated on filter paper moistened with distilled water at 25°C in the dark for 3 days. Germinated seedlings were transferred to nutrient solution in 12 L pots and grown for 4 days. The seedlings were then distributed homogeneously in six flat pots of 6 L capacity (12 seedlings per pot) containing identical solution. After growing for 4 days, plants were exposed to 100 μM Cd as Cd(NO3)2 for 4 days. The composition of the nutrient solution and the growth conditions are according to Chaffai et al. (2005). At harvest, plants were divided into roots and shoots. The roots were washed three times with distilled water. Some plants were dried in an oven at 70°C for 72 h and dry weight determined.
Lipid Extraction
The lipids were extracted according to the method of Folch et al. (1957) modified by Bligh and Dyer (1959). The plant tissues were washed with boiling water for 5 min to denature phospholipases (Douce, 1964) and then ground in chloroform:methanol mixture (2:1, v/v). The water of fixation was added and the homogenate was centrifuged at 3000 rpm for 15 min. The lower chloroformic phase was aspirated and dried under N2 stream on a block heater (40°C) and then, the residue was immediately redissolved in toluene:ethanol (4:1, v/v).
Fatty Acid Analysis
The polar lipid classes were separated from total lipids by one dimensional-thin layer chromatography (TLC) on silica gel G plates 60 (Merck, Darmstadt, Germany) according to the method of Lepage (1969). An aliquot (50 μL) of total lipids was loaded on TLC plates and developed using a mixture of chloroform:acetone:methanol:acetic acid:water (50:20:10:10:5, v/v/v/v/v).
The plates were dried under N2 and exposed briefly to I2 vapor and the lipids appeared as dark brown spots. The different lipids were identified by comparison to reference lipids by running authentic standards simultaneously with samples.
Individual identified polar lipids were scraped from the plates and extracted with 2 mL of hexane in glass tubes. Samples were trans-methylated for the analysis of fatty acids, as described by Carreau and Dubacq (1978). Trans-methylation was made by the addition of 0.5 mL of 1% sodium methylate, a known amount of methyl heptadecanoic acid as an internal standard, 0.2 mL of 1 N H2SO4 and 1.5 mL of 5% sodium chloride. The superior phase that contains Fatty Acid Methyl Esters (FAME) was aspired and the solvent volume reduced in a stream of nitrogen, prior to analysis. The total fatty acids in lipid extract from root and shoot tissues were trans-methylated in the same manner.
FAME were analyzed by GC-FID (HP 6890 series) on a capillary column (HP Innowax, 30 mx0.25x250 μm) with a stationary phase made of PEG. The analysis conditions were as follows: carrier gas N2 at 1.5 mL min-1, oven temperatures isotherm at 150°C for 1 min, from 150 to 200°C at the rate of 15°C min-1, from 200 to 225°C at the rate of 2°C min-1 and isotherm at 225°C during 2 min. Injector and detector temperatures were held, respectively, at 250 and 275°C. To identify the different fatty acid components, the GC-FID was calibrated for each fatty acid and quantification was made by using standard calibration curves. An estimation of the lipid total unsaturation level (double-bond index for C18 fatty acids DBI) was calculated from the mol% values derived from the gas chromatographic data, according to the equation: [Σ(mol% 18°C fatty acid content x number of double bonds)]/100 as described by Skoczowski et al. (1994).
Determination of Steryl Lipids
Steryl lipids were determined in total lipids according to Huang et al. (1961). An aliquot of lipid extract (200 μL) was evaporated in glass tubes. After addition of 1 mL of acetic acid, the tubes were vortexed and 2 mL of Liebermann-burchard reagent (1 mL of concentrated H2SO4 was added to 20 mL of acetic anhydride) were added. The tubes were incubated at room temperature in darkness for 1 h and the absorbance was measured at 525 nm. Cholesterol (Sigma) was used as a standard.
Estimation of Lipid Peroxidation
The extent of lipid peroxidation was estimated by measuring the amount of MDA by the method described by Heath and Packer (1968), which takes into account the possible influence of interfering compounds in the assay for thiobarbituric acid (TBA)-reactive materials. The plant tissues were ground in 10% TCA containing 0.25% (w/v) TBA. The samples were heated at 95°C for 30 min and after cooling, were centrifuged at 1000 g for 10 min. The absorbance of the supernatant was read at 532 nm and corrected by subtracting the non-specific absorbance at 600 nm. The MDA concentration was calculated using molar extinction coefficient of 155 mM-1 cm-1.
Statistical Analysis
All data were statistically analyzed using one-way ANOVA and differences were considered significant at p≤0.05.
RESULTS
Altered Polar Lipid Composition in Maize Roots Under Cd Stress
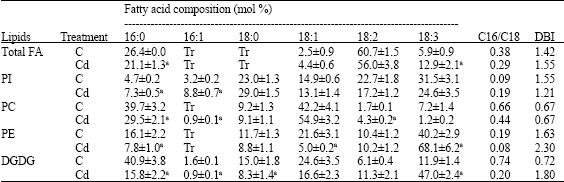
Changes in the relative abundance of the total Fatty Acids (FA) were plotted and are shown in Table 1. Roots of plants exposed to 100 μM Cd, provide fatty acid profile which has decreased saturated palmitic acid 16:0 and increased triunsaturated linolenic acid 18:3, but has unchanged proportions of all other fatty acids (Table 1).
In order to evaluate the toxicity effects of cadmium on membrane lipids, we analyzed the fatty acid composition of various polar lipid classes. There was an increase in the level of 16:0 and 16:1 of phosphatidylinositol (PI) under cadmium stress. In contrast, the level of 16:0 decreased in all other polar lipids: phosphatidylcholine (PC), phosphatidylethanolamine (PE) and digalactosyldiacylglycerol (DGDG). However, the levels of polyunsaturated fatty acids 18:2 (PC) and 18:3 (PE and DGDG) were increased.
| Table 1: | Total and polar fatty acid composition of maize (Zea mays L.) roots. Seedlings were grown in nutrient solutions for 8 days and then treated with 100 μM Cd(NO3)2 for 4 days |
 | |
| The data presented mean values±SE (n = 3) of three separate experiments. a, Significant difference compared to the control at p≤0.05 when applying one-way ANOVA. Tr: Trace | |
| Table 2: | Composition of root lipids of maize (Zea mays L.) seedlings grown in nutrient solutions for 8 days and then treated with 100 μM Cd(NO3)2 for 4 days |
 | |
| The data presented mean values±SE (n = 3) of three separate experiments. Different letters within the same column indicate significant differences (p≤0.05) according to the one-way ANOVA | |
| Table 3: | Polar lipid classes in roots of maize seedlings grown in nutrient solutions for 8 days and then treated with 100 μM Cd(NO3)2 for 4 days |
 | |
| The data presented mean values±SE (n = 3) of three separate experiments. Different letters within the same column indicate significant differences (p≤0.05) according to the one-way ANOVA | |
The levels of monounsaturated fatty acids (16:1 or 18:1) were increased or decreased. A general estimation of the membrane unsaturation level was provided by the double bond indices (DBI) derived from the mol% fatty acid data. As shown in Table 1, cadmium exposure led to an increase in DBI of total FA. The DBI obtained from individual polar lipid classes showed a tendency to increase, while the response pattern to cadmium treatment was opposite for C16/C18 ratio (Table 1).
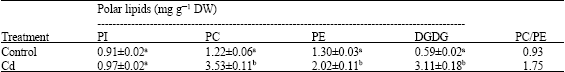
There was no statically significant difference in total FA, phospholipid (PL) and Steryl Lipid (SL) contents in roots, when compared to control plants (Table 2). The SL/PL ratio measured in roots of Cd-treated plants showed a little decrease. In addition, cadmium led to a 2.9, 1.6 and 5.3-fold rise in PC, PE and DGDG, respectively. Consequently, the PC/PE ratio was markedly increased from 0.93 to 1.75 (Table 3).
The level of lipid peroxidation was expressed as MDA content and was determined as 2-thiobarbituric acid (TBA) reactive metabolites. The MDA concentrations increased 21.2% with Cd (Fig. 1a).
Altered Polar Lipid Composition in Maize Shoots Under Cd stress
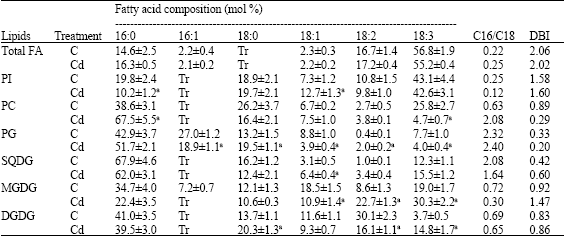
The total FA profile did not change in maize shoots after cadmium exposure. Also, DBI of total FA was not affected. The fatty acid profile of PC showed an increase of the level of 16:0, with a concomitant decrease in 18:3 (Table 4). The fatty acid profiles of major components of chloroplastic membranes PG and MGDG were the most markedly affected. Specifically, monounsaturated 16:1 and 18:1 were substantially decreased, while 18:2 levels increased (Table 4).
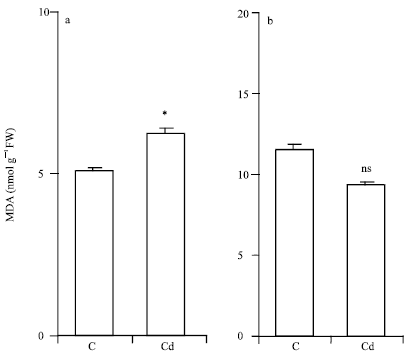 | |
| Fig. 1: | Malondialdehyde (MDA) accumulation in roots (a) and shoots (b) of maize (Zea mays L.) plants grown in hydroponics for 8 days and then treated with 100 μM Cd(NO3)2 for 4 days. The histograms presented mean values±SE (n = 5) of five separate experiments. *, Significance at the p≤0.05 according to the one-way ANOVA. ns, not significant |
| Table 4: | Total and polar lipid fatty acid composition in shoots of maize (Zea mays L.) plants grown in nutrient solutions for 8 days and then treated with 100 μM Cd(NO3)2 for 4 days |
 | |
| The data presented mean values±SE (n = 3) of three separate experiments. a significant difference compared to the control at p≤0.05 when applying one-way ANOVA. Tr: Trace | |
This was associated with marked changes in DBI. The total FA and individual phospholipids (particularly PG) showed a lower degree of unsaturation, as opposed to the glycolipid classes (Table 4).
Present results showed that Cd reduced the GL content by about 36%, but had no significant effect on total FA, PL and SL contents (Table 5). The SL/PL ratio was little increased from 1.44 to 1.92. As noted in Table 6, Cd treatment led to a 78 and 65% decrease in PG and MGDG contents, respectively, whereas levels of SQDG and DGDG were not affected.
| Table 5: | Composition of shoot lipids of maize (Zea mays L.) seedlings grown in nutrient solutions for 8 days and then treated with 100 μM Cd(NO3)2 for 4 days |
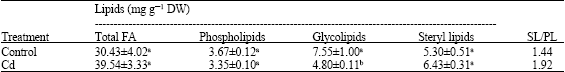 | |
| The data presented mean values±SE (n = 3) of three separate experiments. Different letters within the same column indicate significant differences(p≤0.05) according to the one-way ANOVA | |
| Table 6: | Polar glycerolipids in shoots of maize (Zea mays L.) plants grown in nutrient solutions for 8 days and then treated with 100 μM or Cd(NO3)2 for 4 days |
 | |
| The data presented mean values±SE (n = 3) of three separate experiments. Different letters within the same column indicate significant differences (p≤0.05) according to the one-way ANOVA | |
In addition, PC content was significantly increased (4.4-fold) compared to control plants. Consequently, the MGDG/DGDG ratio was decreased from 1.32 to 0.47 (Table 6). There was no significant change in MDA content under Cd stress (Fig. 1b).
DISCUSSION
The 100 μM of cadmium was chosen in this study so that it caused 50% reduction of root fresh weight. This concentration was selected after a plant growth experiment employing a series of increasing Cd concentrations (0-200 μM).
After cadmium exposure, the increase in DBI of total FA suggests that desaturases activity controlling the biosynthetic conversion of 18:0 to its unsaturated acyl groups was modulated via the desaturation pathways. Thus, we proposed that cadmium mediated an increase in the membrane fluidity. Previous studies demonstrated that the membrane unsaturation was closely related to heavy metal tolerance in a number of higher plants (Maksymiec, 1997). In agreement, aluminum stress resulted in an increase or a lower reduction in the membrane fluidity in an Al-tolerant sorghum cultivar (Peixoto et al., 2001) and an increase of that in an Al-resistant fungus, Lactarius piperatus (Zel et al., 1993a). In contrast, changes in the membrane fatty acid composition by decreasing the membrane fluidity have been reported in Thermoplasma acidophilum (Vierstra and Haug, 1978) and in an Al-sensitive fungus, Amanita muscaria (Zel et al., 1993b) under Al stress.
Present study revealed that an increase in PC occurs concomitantly with the decrease in the SL/PL ratio, which can be considered as additional evidence that membrane fluidity is increased. This could be another aspect of altering membrane composition, or could be an indicator of regulatory mechanism that is required to maintain optimum membrane fluidity under cadmium toxicity. A lower SL/PL ratio has been associated with an increase in membrane fluidity (Yoshida and Uemura, 1990; Palta et al., 1993). Free sterols confer mechanical resistance on membrane bilayers by increasing the efficiency of phospholipids packing (Zhang et al., 1997), while an elevated abundance of PC produce less packed membranes, thereby increasing membrane fluidity (Shinitzky, 1984). Phospholipids play an important role in the regulation of activities of membrane-bound enzymes and in membrane fluidity and permeability (Quartacci et al., 2000) and their variation may be markedly influenced within species, the experimental plant material and under different conditions of stress. For example, exposure of maize to Al has been found to induce a marked increase in total PL (Chaffai et al., 2005). On the contrary, Al treatment decreased the PL in the microsomal membranes purified from 5 mm root tips of an Al-resistant (PT741) and Al-sensitive (Katepwa) cultivars of wheat (Zhang et al., 1996). These results differed from the response in plasma membranes from entire roots, where exposure to 20 μM Al did not affect the PL in both cultivars (Zhang et al., 1997).
The increase in PC/PE ratio observed in the present study indicated that the conversion of PE to PC could be disrupted. The PC/PE ratio has been shown to decrease in pepper roots under Cd stress, but this ratio was found to increase in root plasma membranes of Beta vulgaris in response to Al (Lindberg and Griffiths, 1993).
The increase in DGDG in roots may lead to alterations in cation permeability (Zhang et al., 1997) and membrane leakiness. A lower DGDG levels has been shown to limit the uptake of toxic Al species and prevent Al-induced increase in membrane leakiness (Sasaki et al., 1994).
Present findings provide circumstantial evidence that PG and MGDG, main polar lipids in shoots represent potential target of cadmium toxicity. Chloroplast metabolism and function are particularly sensitive to heavy metal pollution in a variety of lower and higher plant species (Krupa and Baszyñski, 1989). The polar lipid alterations have been found to be correlated with the environmental stimuli and their decline is regarded as a general response to inauspicious growth conditions (Sukenik et al., 1993). For example, in chloroplastic membranes of tomato plants, 100 μM Cd has been shown to cause 83 and 91% declines, respectively in GL and PL, due to significant decreases in polar acyl lipids MGDG, DGDG and PG (Djebali et al., 2005). Further, Maksymiec et al. (1994) have shown that copper induced degradation of MGDG in thylakoids of spinach plants. Water-deficit stress has been shown to cause a decrease in total and polar lipids and particularly in PE, PC, MGDG and DGDG in Carthamus tinctorius leaves (Hamrouni et al., 2001). The decline in the MGDG suggests that is the principal target of galactolipases enzymes or the activity of the lipolytic acyl hydrolase (LAH) (Skorzynska et al., 1991). Since MGDG plays an important role in maintaining the structure integrity of the PSII (Murata et al., 1990), it has been proposed that the extent of perturbation in the MGDG metabolism may result in disruption of the membrane organization (Platt et al., 1994). Moreover, the inhibition of the reverse CDP-choline phosphotransferase could give way to a less diacylglycerol (DAG) formation from PC, leading to a lower MGDG formation. Then, a decrease in the MGDG/DGDG ratio observed can be interpreted as a loss of PSII complex stability (Quartacci et al., 2000).
Alteration in the fatty acid unsaturation of PG may represent primary stress response to cadmium toxicity. PG plays main role in protein-to-lipid interactions and has been shown to be a key component in membranes dynamics (Szalontai et al., 2003) and is considered to be associated with increased cold tolerance in several plant species (Murata, 1983). Moon et al. (1995) have implicated the unsaturation level of PG as being important in the removal and replacement of damaged water splitting D1 proteins within PSII in plants. The cadmium-induced changes in PG unsaturation may therefore induce destabilization of such proteins, resulting in alteration of the photosynthetic membranes. As previously reported for maize shoots, the unsaturation level of PG was significantly altered under Al stress (Chaffai et al., 2005). The effect of Al resulted in a reduction of the abundance of 16:0 and 16:1 fatty acids of PG and a pronounced increase of 18:3, concomitantly with the increase in DBI (Chaffai et al., 2005). The leaves of tomato plants changes its membrane fatty acid composition by decreasing the level of unsaturated fatty acids (18:2, 18:3) and increasing the level of saturated lipids (16:0) with increasing copper or cadmium stress (Ouariti et al., 1997).
The increase in the MDA content in roots after cadmium treatment is consistent with the contention that heavy metal stress strongly affected lipid metabolism and leads to disruption or destabilization of membrane structure and this effect is likely due to a significant production of reactive oxygen species. Cadmium can lead to oxidative stress by displacement of essential heavy metal ions in reaction centers of proteins, resulting in the loss of their biological function and the release of free ions (Hall, 2002). It is also possible that cadmium induced an oxidative stress by the production of free radicals, inhibiting the antioxidant enzyme activity or also by reducing the non enzymatic oxidants (Shaw, 1995). A possible explanation for the absence of MDA change in shoots could be an alleviation of Cd-induced oxidative stress by changing the antioxidant status responses. This includes an increase in the capacity of reactive oxygen species scavenging by increasing the activity of antioxidant enzymes or the levels of the non enzymatic antioxidants.
ACKNOWLEDGMENTS
We are grateful to to Dr. Sami Ben Jamaa for providing extremely valuable comments. We are also indebted to Mrs. Najeh Galai for her technical assistance and excellent help in preparing the manuscript.
REFERENCES
- Bligh, E.G. and W.J. Dyer, 1959. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol., 37: 911-917.
CrossRefPubMedDirect Link - Chaffai, R., B. Marzouk and E.E. Ferjani, 2005. Aluminum mediates compositional alterations of polar lipid classes in maize seedlings. Phytochemistry, 66: 1903-1912.
PubMedDirect Link - Djebali, W., M. Zarrouk, R. Brouquisse, S. El Kahoui, F. Limam, M.H. Ghorbel and W. Chaibi, 2005. Ultrastructure and lipid alterations induced by cadmium in tomato (Lycopersicon esculentum) chloroplast membranes. Plant Biol., 7: 358-368.
PubMedDirect Link - Folch, J., M. Lees and G.H.S. Stanley, 1957. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem., 226: 497-509.
CrossRefPubMedDirect Link - Gallego, S.M., M.P. Benavides and M.L. Tomaro, 1996. Effect of heavy metal ion excess on sunflower leaves: Evidence for involvement of oxidative stress. Plant Sci., 121: 151-159.
CrossRefDirect Link - Hall, J.L., 2002. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot., 53: 1-11.
CrossRefDirect Link - Hamrouni, I., H. Ben Salah and B. Marzouk, 2001. Effects of water-deficit on lipids of safflower aerial parts. Phytochemistry, 58: 277-280.
CrossRefDirect Link - Heath, R.L. and L. Packer, 1968. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys., 125: 189-198.
CrossRefPubMedDirect Link - Huang, T.C., C.P. Chen, V. Wefler and A. Raftery, 1961. A stable reagent for the Liebermann-Burchard reaction. Application to rapid serum cholesterol determination. Anal. Chem., 33: 1405-1407.
Direct Link - Lindberg, S. and G. Griffiths, 1993. Aluminium effects on ATPase activity and lipid composition of plasma membranes in sugar beet roots. J. Exp. Bot., 44: 1543-1550.
CrossRefDirect Link - Maksymiec, W., 1998. Effect of copper on cellular processes in higher plants. Photosynthetica, 34: 321-342.
CrossRefDirect Link - Moon, B.Y., S.I. Higashi, Z. Gombos and N. Murata, 1995. Unsaturation of the membrane lipids of chloroplasts stabilizes the photosynthetic machinery against low-temperature photoinhibition in transgenic tobacco plants. Proc. Natl. Acad. Sci. USA., 92: 6219-6223.
Direct Link - Ouariti, O., N. Boussama, M. Zarrouk, A. Cherif and M.H. Ghorbal, 1997. Cadmium- and copper-induced changes in tomato membrane lipids. Phytochemistry, 45: 1343-1350.
CrossRefPubMedDirect Link - Palta, J.P., B.D. Whitaker and L.S. Weiss, 1993. Plasma membrane lipids associated with genetic variability in freezing tolerance and cold acclimation of Solanum species. Plant Physiol., 103: 793-803.
PubMed - Peixoto, P.H.P., J. Cambraia, R.S. Anna, P.R. Mosquim and M.A. Moreira, 2001. Aluminium effects on fatty acid composition and lipid peroxidation of a purified plasma membrane fraction of root apices of two sorghum cultivars. J. Plant Nutr., 24: 1061-1070.
Direct Link - Quartacci, M.F., C. Pinzino, C.L.M. Sgherri, F. Dalla Vecchia and F. Navari-Izzo, 2000. Growth in excess copper induces changes in the lipid composition and fluidity of PSII-enriched membranes in wheat. Physiol. Plant., 108: 87-93.
CrossRefDirect Link - Di Toppi, L.S., M. Lambardi, L. Pazzagli, G. Cappugi, M. Durante and R. Gabbrielli, 1999. Response to cadmium in carrot in vitro plants and cell suspension cultures. Plant Sci., 137: 119-129.
CrossRef - Schutzendubel, A. and A. Polle, 2002. Plant responses to abiotic stresses: Heavy metal-induced oxidative stress and protection by mycorrhization. J. Exp. Bot., 53: 1351-1365.
CrossRefPubMedDirect Link - Shaw, B.P., 1995. Effects of mercury and cadmium on the activities of antioxidative enzymes in the seedlings of Phaseolus aureus. Biol. Plant., 37: 587-596.
CrossRefDirect Link - Szalontai, B., Z. Kota, H. Nonaka and N. Murata, 2003. Structural consequences of genetically engineered saturation of the fatty acids of phosphatidylglycerol in tobacco thylakoid membranes. An FTIR study. Biochemistry, 42: 4292-4299.
PubMedDirect Link - Zhang, G., J.J. Slaski, D.J. Archambault and G.J. Taylor, 1996. Aluminum-induced alteration in lipid composition of microsomal membrane from an aluminum-resistant and aluminum sensitive cultivar of Triticum aestivum. Physiol. Plant., 96: 683-691.
Direct Link - Zhang, G., J.J. Slaski, D.J. Archambault and G.J. Taylor, 1997. Alteration of plasma membrane lipids in aluminum-resistant and aluminum-sensitive wheat genotypes in response to aluminum stress. Physiol. Plant., 99: 302-308.
Direct Link - Zhang, M., R. Barg, M. Yin, Y. Gueta-Dahan and A. Leikin-Frenkel et al., 2005. Modulated fatty acid desaturation via overexpression of two distinct ω-3 desaturases differentially alters tolerance to various abiotic stresses in transgenic tobacco Cells. Plant J., 44: 361-371.
CrossRefDirect Link