
I. M. Daghman
Department of Land Management, Faculty of Agriculture, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
M. Sariah
Department of Land Management, Faculty of Agriculture, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
J. Kadir
Department of Land Management, Faculty of Agriculture, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
M. A. Zainal Abidin
Department of Land Management, Faculty of Agriculture, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
A. B. Rosenani
Department of Land Management, Faculty of Agriculture, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
International Journal of Agricultural Research
Year: 2006 | Volume: 1 | Issue: 6 | Page No.: 590-596







ABSTRACT
Preparation of Trichoderma harzianum (UPM40) with compost (OPTCD) as the food-base carrier was evaluated for the control of Rhizoctonia damping-off in leaf mustard (Brassica rapa L). Seeds sown in Rhizoctonia-infested soil treated with UPM40 dry preparation and fungicide (Brassicol®) gave significantly higher seedling emergence. The losses in emergence were only 2.83 and 3.71%, respectively, as compared to 21.99% in the untreated soil. The Disease Incidence (DI) expressed as the Area Under the Disease Progress Curve (AUDPC) was highest for the seedlings in the untreated soil and lowest for the seedlings in the UPM40- and fungicide-treated soils. The local isolate T. harzianum (UPM40) dry preparation was therefore effective in protecting the seeds and seedlings of B. rapa against R. solani pre- and post-emergence damping-off. No significant differences in the mean dry weights between the Trichoderma- and chemically-treated soils were observed in the Rhizoctonia-infested soils. However, in the non-infested soils, the mean dry weight of plants was significantly higher for the treatment with UPM40 (23.88 g) followed by that with fungicide (20.88 g) and then the control (16.78 g).
PDF Abstract XML References
How to cite this article
I. M. Daghman, M. Sariah, J. Kadir, M. A. Zainal Abidin and A. B. Rosenani, 2006. Dry Preparation of Trichoderma harzianum for Controlling Rhizoctonia damping-off in Brassica rapa. International Journal of Agricultural Research, 1: 590-596.
DOI: 10.3923/ijar.2006.590.596
URL: https://scialert.net/abstract/?doi=ijar.2006.590.596
DOI: 10.3923/ijar.2006.590.596
URL: https://scialert.net/abstract/?doi=ijar.2006.590.596
INTRODUCTION
In Peninsular Malaysia, Brassica rapa L. (leaf mustard), is the most popular leafy vegetable with 2,402 ha under it in 1999 (Department of Agriculture Malaysia, 1999). Intensive cultivation of leaf mustard makes it vulnerable to soil-borne pathogens such as Rhizoctonia that cause damping-off in many vegetable crops (Sokhi, 1994). Rhizoctonia solani Kuhn is an important seed- and soil-borne plant pathogen associated with seed rot and seedling decay in vegetables (Chan, 1999), thus reducing germination and causing pre- and post-emergence damping-off in seedlings, resulting in a poor plant stand. Practically all farmers rely intensively on chemicals. Indiscriminate and excessive use of chemicals for seed and soil treatment has caused the development of pathogen resistance to pesticides, pesticide contamination of produce, poisoning and a rise in the crop production costs. The use of biological control agents, however, is hampered by the lack of suitable formulations and application methods (Papavizas and Lewis, 1981). Soil amendments with an energy source, such as organic materials, to encourage beneficial microorganisms may also reduce or eliminate fungistasis (Lockwood, 1977; Papavizas and Lewis, 1983) and stimulate proliferation of the antagonists. Trichoderma harzianum (UPM40), an indigenous fungal isolate, has been shown to actively attack R. solani in vitro (Daghman, 2004). To have the same efficacy of the biocontrol agent in the field as in the laboratory, a proper formulation and good application techniques are needed. This study was to evaluate the efficacy of Trichoderma (UPM40) dry preparation, applied as an additive to the soil mixture, in controlling R. solani diseases in B. rapa. Mature compost prepared from Oil Palm Trunk and Chicken Dung (OPTCD) was incorporated as the food base-carrier in the formulation (Daghman, 2004).
MATERIALS AND METHODS
Preparation Fermentor Biomass of UPM40
T. harzianum (UPM40) was obtained from the Plant Protection Laboratory, Faculty of Agriculture, Universiti Putra Malaysia and confirmed to have antagonistic properties against R. solani (Daghman, 2004). The fungal culture was maintained on Potato Dextrose Agar (PDA) as the biocontrol candidate to be tested in this study. Production of the fungal biomass was carried out in 2 L bench-top fermentor flasks with a working volume of 1.5 L sterilized culture medium comprising a mixture of Rice Flour (RF) and molasses (ML) (1:4 RF:ML%). The medium was inoculated with 15 mL spore suspension (15x 108 spores mL-1) of UPM40 and maintained at 30°C in the incubator shaker (500 rpm) for 72 h. The biomass obtained (conidia and mycelia) were stored at -20°C for 24 h and then freeze-dried over 48 h, ground to fine powder and labeled as the dried-fermentor biomass of UPM40.
Preparation of Rhizoctonia Solani Inoculum
R. solani was isolated from infected leaf mustard and a pure culture maintained on PDA. Rhizoctonia inoculum was raised in a sterilized wheat grain medium (100 g/bag) incubated at room temperature (28±2°C) for 2 weeks. The medium was then mixed thoroughly in the soil mixture at 3% of the total soil weight. The artificially infested soil was incubated for four days for the pathogen to establish. The population of R. solani in the soil was evaluated initially (4 random samples per tray), that is, immediately after the 4 days’ incubation and at the end of the experiment 36 days later (Rodriguez-Kabana et al., 1980).
Efficacy of UPM40 Dry Formulation in the Control of Rhizoctonia Diseases
The freeze-dried fermentor biomass of UPM40 was incorporated in OPTCD in the ratio 1:10 w/w fungal biomass: compost. A soil mixture was prepared by thoroughly mixing topsoil, peat grow and sand in 3:2:1 v/v ratio. The soil mixture was placed in aluminum trays (90x 60x30 cm) with each tray containing 25 kg soil mixture. The formulated biomass was added to the soil mixture at 2% of the total soil weight. Random soil samples were taken from the aluminum trays (4 random samples from each tray) to determine the initial antagonist populations expressed as colony forming units (cfu) per g-1 air-dried soil and at 36 days after sowing (Papavizas and Lewis, 1981). B. rapa seeds were primed by soaking in distilled water for 24 h at room temperature to give them an advantage in moisture imbibition to enhance their germination. The treatments were soils treated with UPM40 dry preparation (T1) and fungicide (Brassicol® 200 mg L-1) (T2) and untreated soil (control, T3). The primed seeds were then sown in the soils.
The experimental design was factorial with the trays (replicates) arranged in a Completely Randomized Design on glasshouse benches. Each treatment was replicated four times with each replicate consisting of 30 seeds. The plants were watered daily and organic fertilizer (chicken manure) applied as recommended for B. rapa. The emerging seedlings were counted at day 4 after sowing and the percentage emergence calculated from the total number of seeds sown.
Seeds that failed to emerge or which had rotted were recorded as having pre-emergence damping-off. The incidence of Rhizoctonia post-emergence damping-off was recorded based on the symptoms of wilt, lesions, collapsed hypocotyls or dead seedlings. Re-isolation from the infected seeds and plant parts on Potato Dextrose Agar (PDA) was done to confirm the casual pathogen. The Disease Incidence (DI) was calculated based on the foliar-associated symptoms assessed every two days, according to the formula (modified from Campbell and Madden, 1990):
The reduction in DI, as a measure of the effectiveness of the treatments in suppressing the disease progress, was assessed using the data plotted as disease progress curves and the Area Under the Disease Progress Curve (AUDPC) calculated based on:
Whereby,
Slopes of the disease progress curves were obtained by transforming the DI data using a monomolecular model (Monit) by Campbell and Madden (1990). The effect of the dry formulation of Trichoderma on plant vigor was determined by the difference in dry weight increase of the seedlings and dry weight increase of the roots in infested vs. non-infested soils. The antagonist population on the roots of the seedlings was also assessed at 36 days after sowing in the Rhizoctonia-infested and non-infested soils.
RESULTS AND DISCUSSION
The UPM40 dry preparation and chemically-treated soils both suppressed the activity of R. solani in the Brassica rhizosphere and subsequently, the disease development. Brassica seeds sown in Rhizoctonia non-infested soil with (T1) and without UPM40 (T3) gave emergence of 88.33 and 83.33%, respectively, not significantly different from that of the chemically-treated soil (T2; 90.00%). However, when the seeds were sown in Rhizoctonia-infested soil, T1 and T2 gave significantly better emergence at four days after sowing than T3. Infected seeds had rotted and failed to germinate. The percentage reduction in emergence in the Rhizoctonia-infested soil from the control (T3) was 2.83 and 3.71%, respectively. This indicated that UPM40 dry preparation was effective in protecting the seeds against Rhizoctonia pre-emergence damping-off. As the emerged seedlings continued to grow, they were further subjected to Rhizoctonia infection resulting in post-emergence damping-off. Typical symptoms observed on the seedlings were water-soaked lesions on the hypocotyls, which subsequently collapsed, resulting in death of the seedlings. Re-isolation from the infected seeds and plant parts on PDA plates consistently produced pure cultures of the pathogen R. solani, thus confirming its pathogenicity on Brassica rapa.
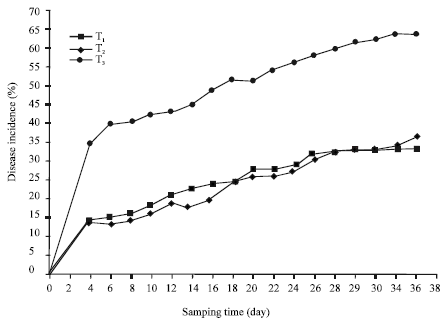 | |
| Fig. 1: | Effect of UPM40 dry formulation on the incidence of Rhizoctonia damping-off in B. rapa seedlings from Rhizoctonia-infested soil. (T1= UPM40-treated; T2 = fungicide-treated; T3 = control) |
The Disease Incidence (DI) was used to indicate the effectiveness of UPM40 dry formulation in suppressing the damping-off. The DI was rated based on symptoms that developed on the hypocotyls, stems and leaves. It was highest in T3 and lowest in T1 and T2 (Fig. 1). UPM40 dry preparation was able to control post-emergence damping-off based on it DI being significantly lower (30%) than that for the control (T3; 65%), 36 days after sowing. Brassicol®, applied as soil drench (T2; 25%), was as effective as UPM40, but the continuous application of fungicide will not only have an adverse effect on the environment but, more importantly, also on the produce. The disease development, as expressed by the AUDPC values of infected seedlings, at 36 days after sowing was 3616.7 units/square for T3 and 2347.7 units/square and 2433.9 units/square for T1 and T2, respectively. The disease developed faster in the seedlings in the non-treated soil with an epidemic rate (rm) of 0.871 unit/day as compared to T1 (rm = 0.655 unit/day) and T2 (rm = 0.718 unit/day) (Table 1). Since R. solani is pathogenic to many vegetable crops, the use of UPM40 dry formulation can be an alternative control strategy against damping-off to obtain a good plant stand and quality produce.
UPM40 has been proven to be strongly antagonistic against R. solani in in vitro dual culture and colony degradation tests with a percentage inhibition in radial growth (PIRG) of 82.72% (Daghman, 2004) and its potential as biocontrol agent against R. solani has been clarified in this study. The mode of antagonism is possibly through competition and mycoparasitism. Competition occurs when the antagonist reduces the pathogenic activity by consuming some/most of the limited resources, such as organic and inorganic nutrients, oxygen and space (Graham and Mitchell, 1999) and denying them to the pathogen. UPM40 overgrew a colony of R. solani within 7 days of co-incubation in the laboratory and antagonistic activity of T. harzianum, T. viride and G. virens has been reported previously against R. solani (Kim and Roh, 1987; Mathew and Gupta, 1998). T. harzianum had also been reported to suppress diseases by other soil-borne fungi (Mukherjee et al., 1995; Begum et al., 1998; Jinantana and Sariah, 1998).
| Table 1: | Epidemic rates and areas under the disease progress curve (AUDPC) for Rhizoctonia damping-off under the different soil treatments at 36 days after sowing |
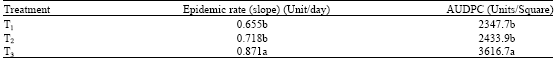 | |
| Means in the same column with the same letter are not significantly different by the LSD test (p = 0.05) on their transformed values. Each value is the mean of four replicates, T1 = UPM40-treated, T2 = fungicide-treated T3= control AUDPC= Area Under Disease Progress Curve | |
| Table 2: | Mean dry weights of plants from the different treatments at 36 days after sowing |
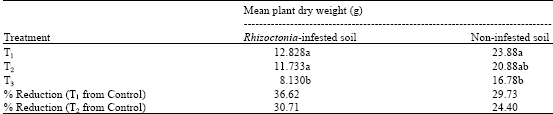 | |
| Means in the same column with the same letter are not significantly different by the LSD test (p≤0.05) on their transformed values. Each value is the mean of four replicates, T1 = UPM40-treated l, T2 = fungicide-treated, T3= control | |
| Table 3: | Average colony forming units (cfu) of UPM40 recovered from the rhizosphere and rhizoplane of B. rapa, 36 days after sowing in Rhizoctonia- infested and non-infested soils |
 | |
| Means within the same row with the same letter are not significantly different by the LSD test (p≤0.05). Each value is the mean of four replicates | |
The ability of Trichoderma to promote the growth of B. rapa was also assessed based on the total dry weight increase of the plants. In the Rhizoctonia-infested soils, there was no significant difference in the total dry weights between the UPM40- and fungicide-treated soils, suggesting that both treatments were equally effective in controlling the damping-off. However, the Rhizoctonia-infested and non-infested soils amended with the dry preparation of UPM40 (T1) produced significantly higher total dry weights than the untreated soil (T3) (Table 2). Their percentages reductions were 36.62 and 29.73% from their respective controls. This could have been due to strong competition by the pathogen for food and space which reduced the antagonist populations in the rhizosphere and rhizoplane of the seedlings. The results were similar to those of Chan (1999) who reported that T. virens significantly increased the plant stand and height, suggesting its possible role as a plant growth promoting fungus. Windham et al. (1986) also reported that T. harzianum produced a growth regulating factor that increased the rate of germination and stem dry weight increase in tomato. It has been postulated that in the biocontrol of pathogens in the rhizosphere, nutrient release from the soil and organic matter and enhanced plant hormone production may partially explain the Trichoderma-stimulated growth responses (Inbar et al., 1994).
Enhancement of root growth was also observed in the plants grown in the UPM40-treated soil with a root dry weight of 9.12 g, followed by those of the chemically-treated (8.39 g) and untreated (5.72) soils which facilitated the uptake of water and nutrients. This resulted in a better crop health leading to a better stand, yield and, possibly, even better nutritive value of the produce. Cheah and Page (1997) found that Trichoderma effectively reduced club root and increased the fresh top weight of Chinese cabbage. Trichoderma also reduced root rot in diseased avocado seedlings and stimulated root regeneration in the treated plants (Sait, 2000).
The ability of UPM40 to proliferate and survive in the rhizosphere and rhizoplane of B. rapa was estimated based on the cfu’s of Trichoderma recovered using the Trichoderma selective media (TME). The population of UPM40 in the rhizosphere had decreased significantly by day 36 in the Rhizoctonia-infested and non-infested soils (Table 3). The decrease in propagules of UPM40 could have been due to competition by the indigenous microorganisms as the soil mixture used was not sterilized. Competition by indigenous microbes is likely to be a major determinant for successful establishment of introduced inoculants in the rhizosphere to a level sufficient for disease suppression. However, recovery of UPM40 was significantly higher from the roots than the rhizosphere, suggesting that it was migrating from the rhisosphere to the roots, colonizing the germinating roots and living on the root exudates. Plant roots are also a major source of carbon and energy for microorganisms in their production of exudates. Competition for the nutrients, primarily carbon, nitrogen and iron, may result in biological control of soil-borne plant pathogens (Sivan and Chet, 1989). For soil-borne diseases, a Trichoderma population in the soil mixture in the magnitude of 104 - 106 was reported to have a beneficial effect on plant growth and disease suppression (Papavizas, 1985).
In addition, the biomass of UPM40 used in this study comprised mainly of conidia and mycelium although the chlamydospores are thought more important in the survival and proliferation of the biocontrol candidate. Therefore, using a biomass of chlamydospores may enhance the effectiveness of UPM40 in controlling Rhizoctonia damping-off in Brassica. Studies on the distribution of UPM 40 as root colonists with time and space are needed to describe and predict the root colonization process. This would enable more effective management of the rhizosphere population for better biological control of Rhizoctonia diseases and enhanced plant growth. It should be followed up with locality trials under a range of conditions since this study is a pioneer research in the use of local isolate of T. harzianum for the control of Rhizoctonia damping-off in Brassica.
REFERENCES
- Campbell, C.L. and L.V. Madden, 1990. Introduction to Plant Disease Epidemiology. Wiley, Hoboken, New Jersey, ISBN-13: 9780471832362, Pages: 560.
Direct Link - Azcon-Aguilar, C. and J.M. Barea, 1997. Arbuscular mycorrhizas and biological control of soil-borne plant pathogens-an overview of the mechanisms involved. Mycorrhiza, 6: 457-464.
CrossRefDirect Link - Inbar, J., M. Abramsky, D. Cohen and I. Chet, 1994. Plant growth enhancement and disease control by Trichoderma harzianum in vegetable seedlings grown under commercial conditions. Eur. J. Plant Pathol., 100: 337-346.
CrossRefDirect Link - Sivan, A. and I. Chet, 1989. The possible role of competition between Trichoderma harzianum and Fusarium oxysporum on rhizosphere colonization. Phytopatholgy, 79: 198-203.
Direct Link - Windham, M.T., Y. Elad and R. Baker, 1986. A mechanism for increased plant growth induced by Trichoderma spp. Phytopathology, 76: 518-521.
Direct Link - Papavizas, G.C., 1985. Trichoderma and Gliocladium: Biology, ecology and potential for biocontrol. Annu. Rev. Phytopathol., 23: 23-54.
CrossRefDirect Link