
Jorge Rattin
Integrada I.N.T.A. Estaci6n Experimental Agropecuaria Balcarce, Provincia de Buenos Aires, Argentina
Adalberto Di Benedetto
Catedra de Floricultura (FAUBA), Av. San Martin 4453 (1417),
Buenos Aires, Argentina
Tomas Gornatti
Integrada I.N.T.A. Estaci6n Experimental Agropecuaria Balcarce, Provincia de Buenos Aires, Argentina
International Journal of Agricultural Research
Year: 2006 | Volume: 1 | Issue: 1 | Page No.: 58-67







ABSTRACT
The aim of this study was to determine the suitability of the transplant routine using plastic plug trays as an alternative of direct seeded and the effect of sowing date for different sweet maize mutants. Sweet maize mutants hybrids Canner (su1), Butter Sweet (sh2) and Cheyenne (se1) were sown in spring and summer by direct seeded or in plastic plug trays (128 cells tray-1) for transplant. Present results suggest that it was possible to obtain similar or highest yield from sweet maize mutant plants using a transplant method than direct-seeded under optimal environments. The disagreement with previous reports on the effects of the transplant routine would be associated with the use of plug plastic tray instead of the commonly polystyrene trays which would let avoid the damage to root system when seedlings were transplanted.
PDF Abstract XML References
How to cite this article
Jorge Rattin, Adalberto Di Benedetto and Tomas Gornatti, 2006. The Effects of Transplant in Sweet Maize (Zea mays L.). I. Growth and Yield. International Journal of Agricultural Research, 1: 58-67.
DOI: 10.3923/ijar.2006.58.67
URL: https://scialert.net/abstract/?doi=ijar.2006.58.67
DOI: 10.3923/ijar.2006.58.67
URL: https://scialert.net/abstract/?doi=ijar.2006.58.67
INTRODUCTION
There are several endosperm mutations in maize (Zea mays L.) which, when present in the homozygous recessive state, produce qualitative and quantitative differences in kernel carbohydrate metabolism that affect flavor and storability (Coe, 1993). They include traditional sweet corn, sugary1 (su1), shrunken-2 (sh2) and sugary enhancer (se1). These mutants all result in higher sugar and lower starch concentrations, although the magnitude of their effects varies (Evansen and Boyer, 1986). The new cultivars at the present time dominate the fresh market sweet corn for long-distance and wholesale markets.
Despite the desirable quality attributes of sh2, se1 and other endosperm genotypes, their utilization has been hindered by reduced field emergence, seedling vigor and stand uniformity, especially in cold soils (Douglass et al., 1993; Cline and Silvernail, 2002). Reduced stand uniformity results in heterogeneous ear maturity and reduced value and yield.
Field and sweet corn have been transplanted experimentally in an attempt to improve stands and hasten maturity (Khehra et al., 1990; Miller, 1972; Waters et al., 1990; Wyatt and Mullins, 1989). However, transplanting sweet corn remains a questionable practice because it increases production costs, often stunts plant development and decreases yield (Pendleton and Egli, 1969; Wellbaum et al., 2001). Transplant advantages include a better use of seed and space; avoid the environmental risks during the earlier low-rate phases of vegetative growth and gives earlier and more uniform flowering and yield.
There are marked differences in the survival rate of different vegetable species after transplanting, particularly if roots have been damaged, or weather conditions are unfavorable (Wien, 1997). In his classic work on transplanting, Loomis (1925) established three classes of transplanting ease. A commonly transplanted group, which usually survives the process well, includes tomato, lettuce, cauliflower and beets. An intermediate group contains celery, eggplant, onions and pepper. The third category comprises species that are difficult to transplant and includes beans, corn, cucumbers and melons.
The aim of this study was determine the suitability of the transplant routine using plastic plug trays as an alternative of direct seeded and the effect of sowing date for different sweet maize mutants.
MATERIALS AND METHODS
The experiments were conducted at the INTA Balcarce Experimental Station, Argentina (37°45´S, 58° 18´W and altitude 130 m) during the 2002-2003 and 2003-2004 growing seasons on a typic argiudol soil with an organic matter of 5.6% in the first 25 cm depth. Water and nutrients were at non-limiting levels using an irrigation system which kept soil water above 50% of maximum soil available water in the first meter of depth. The experimental field was fertilized with 150 kg N ha-1 (18-46-0). Weeds and insects were adequately controlled.
In 2002, maize mutants hybrids Canner (su1), Butter Sweet (sh2) and Cheyenne (se1) provided by Semillería Basso (Argentina) were sown on 25 October and 30 December by direct seeded (D) or in plastic plug trays (128 cells tray-1) (T) using a river waste-base media. Transplanted plants were grown under greenhouse facilities from sowing to the two-leaf expanded stage (near 15 days from sowing). Final population density for both treatments was 80,000 plants ha-1. The experiment, repeated in 2003 beginning on 26 November and 2 January (2004).
The experiment design was a split plot, with hybrids as main plots and establishment routines (direct seeded and transplant) as subplots. The subplots consisted of four rows of 10 m (one border row on each side) and rows were 0.70 m apart.
Weather records (daily maximum-minimum air temperature and global solar radiation) were obtained from a meteorological station 500 m from the experimental site. Emergence dates were recorded.
Dry matter accumulation was determined by taking plant samples at stages V6, V9; VT and R3 for 2002-2003 experiments and V6, V7, V9, VT and R3 for 2003-2004 experiments according to Ritchie and Hanway (1982). The sample size was 10 plants per row, leaving appropriate border rows and border plans within the rows. Plants were cut at ground level, oven dried at 60°C for 10 days and weighed. Plant height (from ground level to basal tassel branches) and leaf number were determined at all sampled stages.
Data were processed by analysis of variance procedures and appropriate standard errors of the means were calculated.
RESULTS
Total leaf area was higher for ‘Canner’, ‘Butter Sweet’ and ‘Cheyenne’ mutants crops initiated by direct-seeded than by transplant (Fig. 1A-C). First sowing date showed higher leaf area than second sowing date too. ‘Canner’ maize mutant showed the highest differences in leaf area between sowing methods and sowing date at R3 stage.
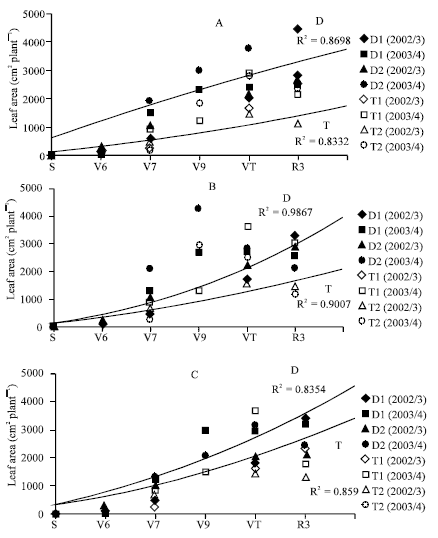 | |
| Fig. 1: | Leaf area increase through developmental stage for ‘Canner’ (A), ‘Butter Sweet’ (B) and ‘Cheyenne’ © sweet maize mutants under direct-seeded (D) or transplant (T) method and two sowing date (1, 2). |
Shoot dry weight was the highest for direct-seeded plants under optimal sowing date for su1 (Canner), sh2 (Butter Sweet) and se1 (Cheyenne) maize mutants hybrids (Fig. 2A-C). By other hand, first sowing date showed higher values than second sowing date.
There were no differences in kernel number per ear between crop initiation method and sowing dates (Table 1). This was the result of similar kernel per row and rows per ear. Lower kernel number from transplanted plants was only found for the se1 (Cheyenne) maize mutant during the first sowing date of 2003-2004 experiment.
The 2002-2003 experiment showed the highest final ear yield (g fresh weight plant-1) at R3 stage for the three maize mutant transplanted plants during the first sowing date (Table 2). This result was repeated in 2003-2004 experiment only for ‘Butter Sweet’; while there were no yield differences for Canner’, a ‘Cheyenne’ higher yield from direct-seeded plants was found.
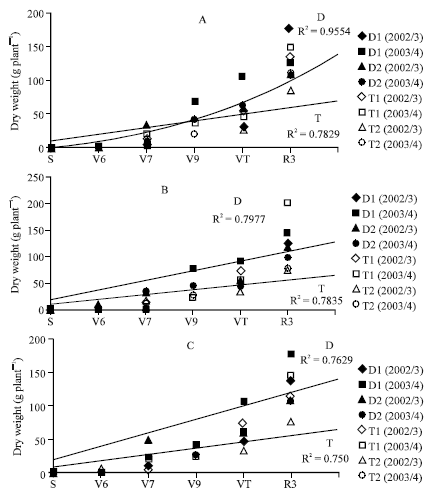 | |
| Fig. 2: | Dry weight accumulation for ‘Canner’ (A), ‘Butter Sweet’ (B) and ‘Cheyenne’ © sweet maize mutants under direct-seeded (D) or transplant (T) method and two sowing date (1, 2). |
| Table 1A: | Number of kernels per ear, row and number of rows per ear at R3 stage for three sweet maize hybrids, at direct seeded or transplant for two sowing date (2002-2003) |
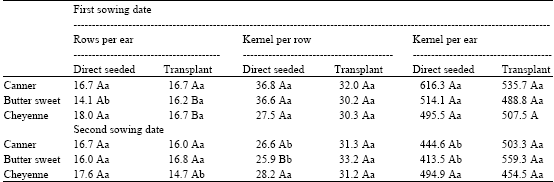 | |
| Capital letter (s) indicate statistically significant differences (p≤0.05) for hybrids between direct seeded and transplant within sowing dates and lower case letter (s) indicate statistically significant differences (p≤0.05) among sowing dates for each treatment | |
| Table 1B: | Number of kernels per ear, row and number of rows per ear at R3 stage for three sweet maize hybrids, at direct seeded or transplant for two sowing date (2003-2004) |
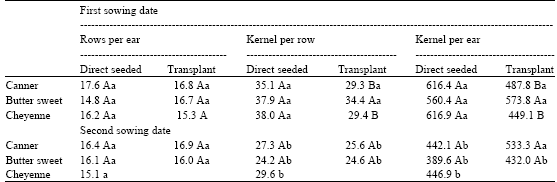 | |
| Capital letter (s) indicate statistically significant differences (p≤0.05) for hybrids between direct seeded and transplant within sowing dates and low-case letter (s) indicate statistically significant differences (p≤0.05) among sowing dates for each treatment | |
| Table 2: | Ear yield (g fresh weight plant-1) at R3 stage for three sweet maize hybrids at direct seeded or transplant for two sowing date during 2 years |
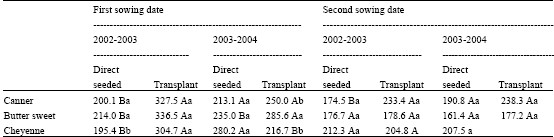 | |
| Capital letter (s) indicate statistically significant differences (p≤0.05) for hybrids between direct seeded and transplant within sowing dates and lower case letter (s) indicate statistically significant differences (p≤0.05) among years for the same treatment and sowing date | |
| Table 3: | Harvest Index at R3 stage for three sweet maize hybrids at direct seeded or transplant for two sowing date during 2 years |
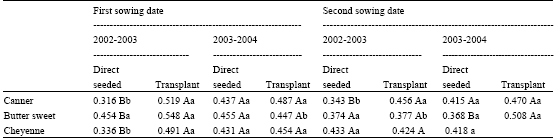 | |
| Capital letter (s) indicate statistically significant differences (p≤0.05) for hybrids between direct seeded and transplant within sowing dates and lower case letter (s) indicate statistically significant differences (p≤0.05) among years for the same treatment and sowing date | |
The second sowing date gives similar yield for the most initiation method, year and maize mutants.
When Harvest Index (HI) on a fresh weight base at R3 stage was analyzed (Table 3), results are similar to obtained for ear yield, this is, highest or the same values for transplanted plants in 2002-2003 and 2003-2004 experiments, respectively for su1 (Canner), sh2 (Butter Sweet) and se1 (Cheyenne) maize mutants hybrids. Second sowing date showed no differences for direct-seeded vs. transplant, except for ‘Canner’ in the 2002-2003 experiment.
| Table 4: | Daily solar radiation, maximum and minimum temperature. Monthly mean for 2002-2003 and 2003-2004 experiments |
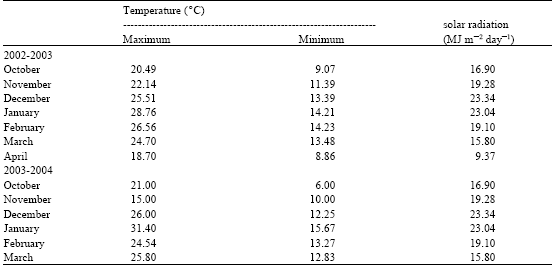 | |
Solar radiation and air temperature during the experiments (Table 4) did not seem to have limited potential maize crop growth for the first sowing date. However, weather conditions during the second sowing date decreased, mainly for the kernel-filling stage.
DISCUSSION
The quality of sweet maize has been enhanced in recent years by the introduction of mutant genes affecting carbohydrate metabolism in the endosperm in place of the standard sugary (su1) gene. The commonly indicated less establishment capacity for the new maize mutants could not be confirmed in our experiments because there were only minor statistical significant differences in total leaf area (Fig. 1), total dry weight (Fig. 2), leaf number and plant height (data not shown) until V7 stage for the direct-seeding routine under optimal sowing date.
Maize grain yield is related to light interception, to radiation-use efficiency and to photoassimilate partitioning. Radiation interception varies from seedling emergence to crop harvest and depends largely on the canopy leaf area (Watiki et al., 1993). Leaf area development is described as a function of air temperature. Since temperature affects many plant processes, including nutrient uptake, water absorption, photosynthesis, respiration and translocation of photosynthate, it is not surprising that temperature is considered the most important environmental factor governing plant development. Mean air temperature was above the thresholds that may affect radiation-use efficiency (16°C) or biomass partitioning to grains (19°C) (Wilson et al., 1995) and solar radiation was highest the levels registered in a cool and cloudy high latitude environment of northern Europe (Otegui and Bonhomme, 1998) for the first sowing date (Table 4). During the second sowing date solar radiation and mean air temperature significantly decreased for the kernel-filling stage.
The most important result of this work was that it was possible to obtain high yield from sweet maize mutant plants using a transplant method and highest than direct-seeded under optimal environments (Table 2). These results are not in agreement with previous reports (Pendleton and Egli, 1969; Wellbaum et al., 2001). The primary objective for transplanting sweet corn is easily maturity; if growers can produce sh2 mutants before the competition in a particular market does, the extra crop value may justify the greater production costs of transplanting.
Harvest index, or grain weight divided by total shoot weight, is considered stable for maize grown in the absence of environmental constraint. Nevertheless, modifications introduced by genotype and environment could be expected, as those induced in dry matter partitioning by temperature during early stages of growth as a ratio of grain to biomass yield, the harvest index of cereal crops can be affected by any factor which influences the components of yield to different extents. Increased plant population density, by favoring biomass production over grain, also tends to result in lower harvest index, but within the normal range of commercial crop densities, such reductions rarely exceed 10% of the optimum value (Hay, 1995). Present results are not in agreement because we found an increase in harvest index using a transplant routine (Table 3). However, it takes into account that harvest index for sweet maize included the ear at a R3 stage.
Biomass accumulation and partitioning to reproductive structures are critical aspects of plant fitness (Andrade et al., 1999). In many plant species, dry matter partitioning to reproductive organs is a stable and highly heritable trait (Sadras et al., 1997). The mechanisms underlying reproductive partitioning are, however, poorly understood. In maize, plant density has important effects on partitioning of dry matter between vegetative and reproductive structures. The effect of an increase in plant density radiation on dry matter partitioning between vegetative and reproductive structures are expected (Andrade et al., 1993). Present results showed that a root restriction (transplant treatment) for sweet maize mutants would change photoassimilate partitioning at behalf to reproductive structures (Table 3) even at higher plant densities (8 plants m-2) that the usually suggested for this crop (4-6 plants m-2).
Pendleton and Egli (1969) proposed that one explanation for the transplants yielding no more than the early seedlings may lie in their shorter plant height and less leaf surface. It has been indicated (Loomis, 1925; Waters et al., 1990; Wellbaum et al., 2001) that maize does not transplant well because pruned roots do not branch and root replacement is generally poor compared with crops such as cabbage or tomato. The inability of maize roots to regenerate after transplanting resulted in stunted plants. The root system of a maize seedling has seminal roots that consist of the radicle or primary root and a variable number of lateral roots that arise adventitiously at the base of the first internode of the stem, just above the scutellar node. The seminal root initials are present in the embryo and are the most important for early growth and establishment.
Wellbaum et al. (2001) has shown that seminal roots were broken during transplanting as they were pulled from transplant growing in polystyrene trays, indicating that this material was not optimal for maize transplant production. However, similar results [less total leaf area (Fig. 1)] were found in plants growing from plastic plug trays which showed intact root systems at the transplant stage. At transplant, sweet corn seedlings showed a well developed root system with white roots and without damage but the plug cell base determined horizontal root growth around it (picture not shown). Root restriction stress related to a small plug cell volume could be limited biomass accumulation during the vegetative stages. This effect has been indicated for other plants under intensive propagation systems (Di Benedetto and Klasman, 2004). Although physiological mechanisms are unclear, both synthesis and translocation of hormones such as auxins and citokinins from root apices would be involved and related to a change in root verticality as a restriction from the plug cell base.
The productivity of maize is constrained by the relative plasticity of the response to environmental stresses. Plant plasticity to abiotic stresses has recently been described as being either morphological or phenotypic in character (Vyn and Hooker, 2002). The understanding of the response of maize to particular short- and long-term stresses is limited even further by interactions with other stresses in the crop such as the root restriction imposed by small plug cell trays when a transplant routine was used.
Plants were harvested at specific stages of vegetative development as measured by mature leaf number. Since leaf appearance rate changed with temperature, the time between sowing and each of the harvests and hence the duration of growth, also varied. Delayed sowings are generally accompanied by increased temperatures during the growing season, which accelerate crop development and decrease accumulated solar radiation (Cirilo and Andrade, 1994; Otegui et al., 1995) resulting in less biomass production. Our results are in agreement with previous reports because R3 stage for both direct-seeding and transplant plants was achieved 7-13 days later, respectively for the second vs. first sowing date (data not shown).
Variation in sowing date in maize modifies the radiative and thermal conditions during growth. The amount of incident radiation and the proportion of this radiation that is intercepted by the crop directly determine crop growth rate. Temperature affects the duration of crop growth and consequently the time during which incident radiation can be intercepted and transformed to dry matter. Temperature also affects final leaf number (Tollenaar and Hunter, 1983) and leaf canopy development (Hesketh and Warrington, 1989) which define crop leaf area index. Therefore, these factors determine the proportion of the incident radiation that is intercepted by the crop. Present results are in agreement with Cirilo and Andrade (1994) which have indicated that significant differences in crop growth and development would be found when sowing date is changed. Late sowing date would be limited by the assimilation leaf source (Fig. 1).
Recently, process-oriented studies of leaf development have shown that under non-drought conditions, each leaf has a well-defined growth rate, which is a function of temperature and a maximum leaf area, which is a function of position on the plant. Thus, in the absence of water stress, leaf area development can be described by relatively simple temperature functions (Dale et al., 1980). However, the ‘root restriction’ stress generated by plug cell determines changes in leaf growth. On account of leaf area decreased (Fig. 1) but not leaf number (data not shown), the individual leaves were smaller for the transplant.
Clipping of early leaves has been shown to increase Harvest Index (HI) of maize Zea mays L. In a study with sweet maize, Crockett and Crookston (1980) measured an average 20% increase in HI following early leaf clipping. The increase was found to result from a greater reduction in stove mass than in ear mass (Crockett and Crookston, 1980, 1981). Results from Table 3 are in agreement although the effect of transplant would be related with another growth restriction such as sowing date.
Clipping reduces stove because photosynthetic leaf area is removed and vegetative development is retarded, it is possible that early clipping must induce a change in the microclimate of the corn plant, specifically in the temperature of the growing point region. Jepson and Crookston (1986) showed that the growing point temperature of maize was significantly increased by early leaf clipping. We also observed that early growing point heating was followed by an alteration in the partitioning of dry matter that favored the ear rather than stove.
We have not recorded growing point temperature in our experiments by which we have not sure if it was related to a change in leaf growth for the transplanted plants, but although transplant determine a decrease in biomass accumulation, mechanism would be different for clipping plants because this treatment commonly decreases yield (Eghareyba et al., 1976) and transplant increases it (Fig. 1). By other hand, the use of transplanted plants would be suggest for basic physiological experiments related to the study of source-sink relationship mechanisms because transplant change the source (leaf area) for intact plants (unclipped) growing under the same abiotical environment.
The aim of this reserch was to study the effect of a change in the establishment routine (transplant) on yield of different sweet maize mutants. Due to our results showed similar yield on unstressed environment (first sowing date) (Table 2) it is clear that transplant would be an alternative to direct-seeding when plastic plug cell trays was used. Although transplant would increase establishment costs the change in plant architecture associated to smaller leaf area accumulation rate let would increase plant density (in fact our experiments were developed at a higher plant density than usually suggested) but this matter require another experimental design and is the base for the following research.
ACKNOWLEDGMENTS
This work was partially supported by the UBACyT 2004-2005 Program (G 031).
The authors gratefully acknowledge the provision of experimental maize hybrids by Martín Garbarino (Semillería Basso) (Buenos Aires, Argentine) and the contribution of Fernando Andrade in providing field facilities and his invaluable suggestions in experiment design.
REFERENCES
- Andrade, F.H., S.A. Uhart and M.I. Frugone, 1993. Intercepted radiation at flowering and kernel number in maize: Shade versus plant density effects. Crop Sci., 33: 482-485.
CrossRefDirect Link - Andrade, F.H., C. Vega, S. Uhart, A. Cirilo, M. Cantarero and O. Valentinuz, 1999. Kernel number determination in maize. Crop Sci., 39: 453-459.
Direct Link - Cirilo, A.G. and F.H. Andrade, 1994. Sowing date and maize productivity I: Crop growth and dry matter partitioning. Crop Sci., 34: 1039-1043.
Direct Link - Cline, G.R. and A.F. Silvernail, 2002. Effects of cover crops, nitrogen and tillage on sweet corn. Hortechnology, 12: 118-125.
Direct Link - Crockett, R.P. and R.K. Crookston, 1980. Tillering of sweet corn reduced by clipping of early leaves. J. Am. Soc. Hortic. Sci., 105: 565-567.
Direct Link - Dale, R.F., D.T. Coelho and K.P. Gallo, 1980. Prediction of daily green leaf area index for corn. Agron. J., 72: 999-1005.
CrossRefDirect Link - Di Benedetto, A. and R. Klasman, 2004. The effect of plug cell volume on the post-transplant growth for Impatiens walleriana pot plant. Eur. J. Hortic. Sci., 69: 82-86.
Direct Link - Douglass, S.K., J.A. Juvik and W.E. Splittstoesser, 1993. Sweet corn seedling emergence and variation in kernel carbohydrate reserves. Soil Sci. Technol., 21: 433-445.
Direct Link - Eghareyba, P.N., R.D. Horrocks and M.S. Zuber, 1976. Dry matter accumulation in maize response to defoliation. Agron. J., 68: 40-43.
Direct Link - Hay, R.K.M., 1995. Harvest index: A review of its use in plant breeding and crop physiology. Ann. Applied Biol., 126: 197-216.
CrossRefDirect Link - Hesketh, J.D. and I.J. Warrington, 1989. Corn growth response to temperature: Rate and duration of leaf emergence. Agron. J., 81: 696-701.
Direct Link - Jeppson, R.G. and R.K. Crookston, 1986. Effect of elevated growing-point temperature on maize growth and yield. Crop Sci., 26: 595-598.
Direct Link - Khehra, A.S., H.S. Brar, R.K. Sharma, B.S. Dhillon and V.V. Malhotra, 1990. Transplanting of maize during the winter in India. Agron. J., 82: 41-47.
Direct Link - Otegui, M.E. and R. Bonhomme, 1998. Grain yield components in maize: I. Ear growth and kernel set. Field Crop Res., 56: 247-256.
Direct Link - Otegui, M.E., M.G. Nicolini, R.A. Ruiz and P.A. Dodds, 1995. Sowing date effects on grain yield components for different maize genotypes. Agron. J., 87: 29-33.
CrossRefDirect Link - Pendleton, J.W. and D.B. Egli, 1969. Potential yield of corn as affected by planting date. Agron. J., 61: 70-71.
Direct Link - Sadras, V.O., M.P. Bange and S.P. Milroy, 1997. Reproductive allocation of cotton in response to plant and environmental factors. Ann. Bot., 80: 75-81.
Direct Link - Tollenaar, M. and T.B. Hunter, 1983. A photoperiod and temperature sensitive period for leaf number of maize. Crop Sci., 23: 457-460.
Direct Link - Vyn, T.J. and D.C. Hooker, 2002. Assessment of multiple- and single-factor stress impacts on corn. Field Crop Res., 75: 123-137.
Direct Link - Waters, L.J., R.L. Burrows, M.A. Bennett and J. Schoenecker, 1990. Seed moisture and transplant management techniques influence sweet corn stand establishment, growth, development and yield. J. Am. Soc. Hortic. Sci., 115: 888-892.
Direct Link - Watiki, J.M., S. Fukai, J.A. Banda and B.A. Keating, 1993. Radiation interception and growth of maize/cowpea intercrop as affected by maize plant density and cowpea cultivar. Field Crop Res., 35: 123-133.
Direct Link - Wellbaum, G.E., J.M. Frantz, M.K. Gunatilaka and Z. Shen, 2001. A comparison of the growth, establishment and maturity of direct-seeded and transplanted sh2 sweet corn. HortScience, 36: 687-690.
Direct Link - Wilson, D.R., R.C. Muchow and C.J. Murgatroyd, 1995. Model analysis of temperature and solar radiation limitations to maize potential productivity on a cool climate. Field Crop Res., 43: 1-18.
Direct Link - Wyatt, J.E. and J.A. Mullins, 1989. Production of sweet corn from transplants. HortScience, 24: 1039-1039.
Direct Link