
Adalberto Di Benedetto
Catedra de Floricultura, Facultad de Agronomia, Universidad de Buenos Aires, Av. San Martin, 4453 (Cl417DSE), Buenos Aires, Argentina
Jorge Molinari
Catedra de Floricultura, Facultad de Agronomia, Universidad de Buenos Aires, Av. San Martin, 4453 (Cl417DSE), Buenos Aires, Argentina
Carlos Boschi
Catedra de Floricultura, Facultad de Agronomia, Universidad de Buenos Aires, Av. San Martin, 4453 (Cl417DSE), Buenos Aires, Argentina
Diego Benedicto
Catedra de Floricultura, Facultad de Agronomia, Universidad de Buenos Aires, Av. San Martin, 4453 (Cl417DSE), Buenos Aires, Argentina
Marcelo Cerrotta
Catedra de Floricultura, Facultad de Agronomia, Universidad de Buenos Aires, Av. San Martin, 4453 (Cl417DSE), Buenos Aires, Argentina
Guillermo Cerrotta
Catedra de Floricultura, Facultad de Agronomia, Universidad de Buenos Aires, Av. San Martin, 4453 (Cl417DSE), Buenos Aires, Argentina
International Journal of Agricultural Research
Year: 2006 | Volume: 1 | Issue: 6 | Page No.: 522-533







ABSTRACT
Saleability of tropical ornamental foliage plants and grower`s profit demand both high plant quality and productivity. The aim of this study has been to make observations during two season of the year: winter and summer, on various structural and photosynthetic parameters of five ornamental foliage plants (Aglaonema commutatum, Dieffenbachia maculata, Philodendron `Burgundy`, Philodendron peruvianum and Syngonium podophyllum) grown in a greenhouse at commercial facilities. The results from this study showed that the more a plant invests in leaf area, the higher the total carbon gain and the faster growth will be. Most foliage plants have their origins in the tropics and require relatively high night temperatures to sustain rapid growth; A significant change in growth rates would be associated to environmental seasonal influences. By other hand, estimation of leaf area through linear dimensions such as length and width of the leaf lamina and the close correlations between total leaf area and total dry weight with plant size would be an appropriate method and a useful tool for foliage ornamental productivity crop modelling.
PDF Abstract XML References
How to cite this article
Adalberto Di Benedetto, Jorge Molinari, Carlos Boschi, Diego Benedicto, Marcelo Cerrotta and Guillermo Cerrotta, 2006. Estimating Crop Productivity for Five Ornamental Foliage Plants. International Journal of Agricultural Research, 1: 522-533.
DOI: 10.3923/ijar.2006.522.533
URL: https://scialert.net/abstract/?doi=ijar.2006.522.533
DOI: 10.3923/ijar.2006.522.533
URL: https://scialert.net/abstract/?doi=ijar.2006.522.533
INTRODUCTION
Foliage plant production represents an important agricultural industry. The demand for tropical foliage plants for use in homes, apartments, hotels, business offices, airports and other public building continues to increase rapidly as individuals in modern society become further removed from natural environments.
The carbon gain is an important aspect of plant performance; there are strong links between resource availability, the ability of plants to use resources for carbon gain and productivity and, at the population level, parameters such as size, survival and reproductive output (Bazzaz et al., 1987).
Several attempts have been made to relate net photosynthetic rate (Pn) to plant productivity in ornamentals (Ottosen, 1990, 1994; Vidal et al., 1990), although it has been pointed out the difficulty of relating leaf Pn rates to yield parameters. Measurements of Pn rates of only parts of plants under laboratory conditions may not fully reflect field conditions, since the impact on plants is a combination of environmental and hereditary factors. As many of the examined species have been grown under field conditions, growth rate determinations based on short term measurements of photosynthesis have proved difficult (Zelitch, 1982). Leaf photosynthetic capacity is age dependent and, as the plant grows, canopy structure and other such changes modify the environment of a particular leaf. Allocation of carbon and nutrients to new leaves is an especially important “reinvestment” because it drives growth in an exponential fashion.
There are numerous well documented examples of size-related physiological changes in animals, but only contradictory results for plants (Di Benedetto et al., 2005; Soyza et al., 1996; Donovan and Ehleringer, 1991, 1992; Di Benedetto and Cogliatti, 1990b). Because total plant photosynthesis is increasing, relatively small changes in allocation to new leaves compounds the investment, leading to large changes in plant size; large differences in growth rates can be attributed to differences in allocation patterns (Pearcy et al., 1987).
The influence that plant size has on physiological responses was addressed under both constant and varying environmental conditions (Di Benedetto and Cogliatti, 1990a; Di Benedetto, 1991; Gerry and Wilson, 1995). Differences in leaf size that correlated with plant size also influenced physiological performance of the plant (Mendez and Obeso, 1993; Zotz, 1997).
Plant leaf area is a useful observation in physiological and agronomic research. Statistical regression and correlation models have been used to estimate leaf blade are from leaf blade dimensions such as length and width. It has been indicated that leaf length measurement were used to successfully estimate leaf area in several plant species. These relationships, derived from statistical regression and correlation models, between leaf blade area and leaf blade dimensions are specific for the plants and growth stage for which they were derived. They cannot be extrapolated to other related species with confidence (Goenaga and Singh, 1996).
The aim of this study has been to make observations during two season of the year: winter and summer, on various structural and photosynthetic parameters of five ornamental foliage plants grown in a greenhouse at commercial facilities.
MATERIALS AND METHODS
Plants of Aglaonema commutatum, Dieffenbachia maculata, Philodendron peruvianum, Philodendron ‘Burgundy’ and Syngonium podophyllum were collected from a commercial source and the experiment was carried out at the greenhouse facilities from Buenos Aires University campus, Argentina (34°28’S) from the beginning autumn 2005 to the end of summer 2005. Greenhouse was covered with black shade cloth (40% sunlight) during summer.
Plants were irrigated with a high quality tap water (pH: 6.64 and electrical conductivity of 0.486 dS m-1) and a weekly fertilization of 50 ppm N (1N:0.5P:1K:0.5Ca v/v) was used.
Leaf area were determined by drawing around the leaves on paper, cutting out the shapes, weighing them and then calculating the area from the known weight per unit area of the paper. The regression lines obtained from the measured values of w (leaf width) and l (leaf length) were used to estimate leaf area.
Plants were harvested at the end of summer and winter season respectively, dried at 80°C for 48 h and weighed to obtain the dry aerial and root biomass weight.
Net rate of CO2 uptake (Pn) was measured at ambient O2 and CO2 concentrations, on the last fully expanded leaf, using a portable LICOR LI-6200 system.
After measurement of Pn, four 0.9 cm diameter discs were removed from the central area (on either side of the midvein) of the last fully expanded leaf and chlorophyll was extracted with 4.0 cm3 of N, N-dimethylformamide at 4°C for 48 h (Moran, 1982). Absorptances at 647 and 664 nm were measured using a spectrophotometer.
Tissue from the middle region of the lamina was fixed in FAA, embedded in paraffin, sectioned at 10 or 20 μm on a rotary microtome and stained with safranin-crystal violet-fast green. Leaf thickness was determined from ten cross-sections per leaf with a microtome scale and a grid in one eyepiece of a microscope using a 10 x objective.
Changes during plant growth were estimated through a straight-line regression. The significance of differences was determined through the Test for zero slopes and Test for zero intercept from Kleinbaum and Kupper (1978).
RESULTS
The regression lines obtained from the measured values of w (leaf width) and l (leaf length) used to estimate single leaf area (Table 1) showed high regression coefficients for the five ornamental plants under study.
| Table 1: | Leaf Area (cm2 leaf-1) estimation used regression lines obtained from the measured values of w (leaf width) and l (leaf length) |
 | |
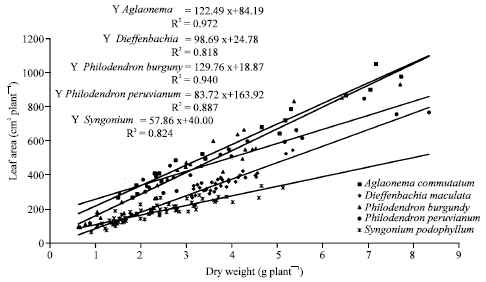 | |
| Fig. 1: | Leaf area changes according to total dry weight for Aglaonema commutatum, Dieffenbachia maculata, Philodendron ‘Burgundy’, Philodendron peruvianum and Syngonium podophyllum ornamental plants |
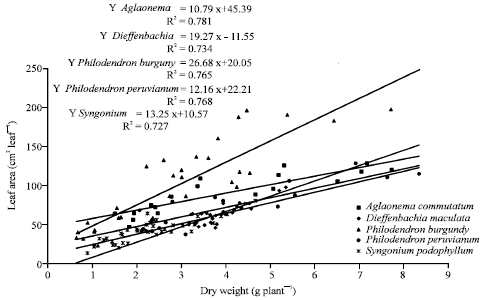 | |
| Fig. 2: | Single leaf area changes according to total dry weight for Aglaonema commutatum, Dieffenbachia maculata, Philodendron ‘Burgundy’, Philodendron peruvianum and Syngonium podophyllum ornamental plants |
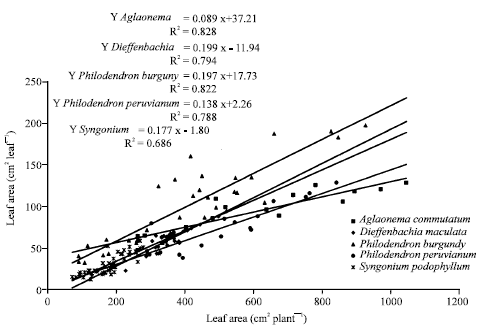 | |
| Fig. 3: | Single leaf area changes according to total leaf area for Aglaonema commutatum, Dieffenbachia maculata, Philodendron ‘Burgundy’, Philodendron peruvianum and Syngonium podophyllum ornamental plants |
Leaf area (cm2 plant-1) increased according dry weight do it; high correlation coefficients for A. commutatum, D. maculata, Ph. ‘Burgundy’, Ph. peruvianum and S. podophyllum were found (R2 = 0.972, 0.081, 0.940, 0.887 and 0.992 respectively) (Fig. 1).
Data for single leaf area (cm2 leaf-1) vs. total dry weight (g plant-1) plotted in Fig. 2 showed a close relationship between both (R2 = 0.781, 0.734, 0.765, 0.768 and 0.727) for the five ornamental plants tested. Regression lines for single leaf area (cm2 leaf-1) vs. total leaf area (cm2 plant-1) (Fig. 3) showed that previous leaf area determine single leaf size. Although plant size estimated as dry weight accumulated or leaf area expanded were the main factor which influences single leaf area, the quantitative relations differ for A. commutatum and the rest of species evaluated here.
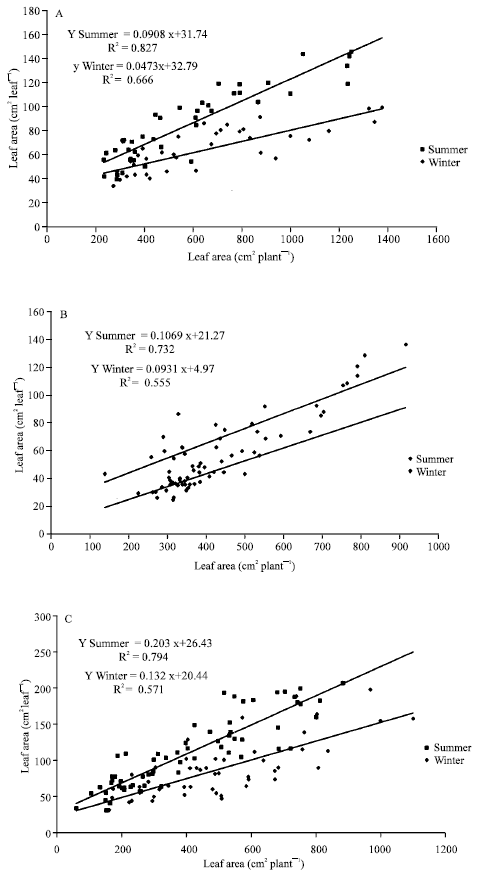
Figure 4 showed the changes in the regression lines related to the season; higher single leaf and an increase at the highest plant sizes during summer were found. Data from winter showed a correlation coefficients decrease for all the ornamental plants tested too. However, there were different responses related to plant tested; by example, A. commutatum (Fig. 4A), Ph. ‘Burgundy’ (Fig. 4C) and Ph. peruvianum (Fig. 4D) showed significant higher values as in the summer line slopes as in the x-axis interception than for winter. By the other hand, D. maculata (Fig. 4B) and S. podophyllum (Fig. 4E) only showed significant differences in x-axis interception values. There were significant differences in the higher single leaf area expanded by each species with the highest values from Ph. ‘Burgundy’ (Fig. 4C).
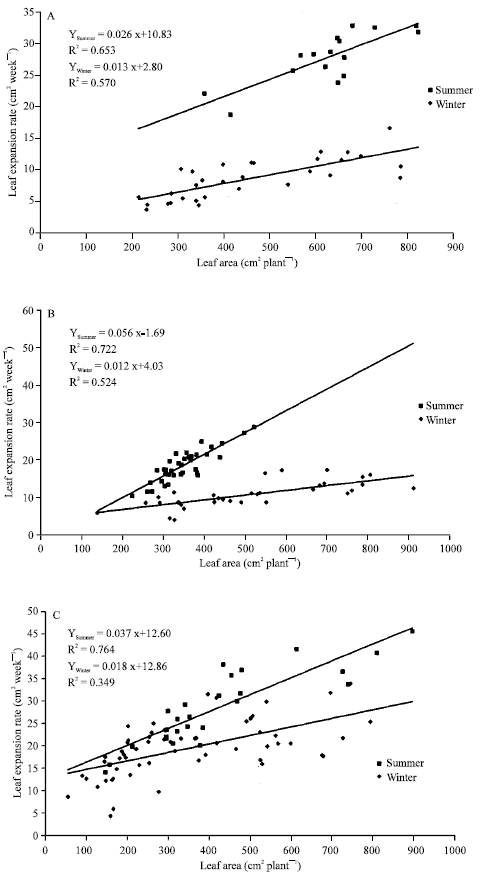
Leaf expansion rates (cm2 week-1) for the five Araceae plants tested increased according size plant do it (Fig. 5). Differences increased during summer and correlation coefficients were lower for the winter harvest but not in the same extension: A. commutatum (Fig. 5A), Ph. peruvianum (Fig. 5D) and S. podophyllum (Fig. 5E) showed a clear higher response to summer growth. A higher line slope but the same x-axis interception for Ph. ‘Burgundy’ (Fig. 5C) was found. By the other hand, Fig. 5B showed higher line slopes but lower x-axis interception for D. maculata under summer environmental facilities.
 | |
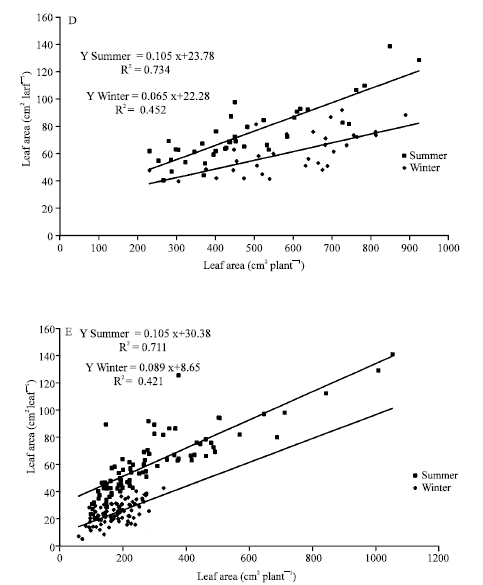 | |
| Fig. 4: | Single leaf area vs. total leaf area for Aglaonema commutatum (A), Dieffenbachia maculata (B), Philodendron ‘Burgundy’ (C), Philodendron peruvianum (D) and Syngonium podophyllum (D) ornamental plants grown under two contrasting seasons |
| Table 2: | Root: Shoot fitted lines and correlations coefficients for five ornamental shade plants |
 | |
 | |
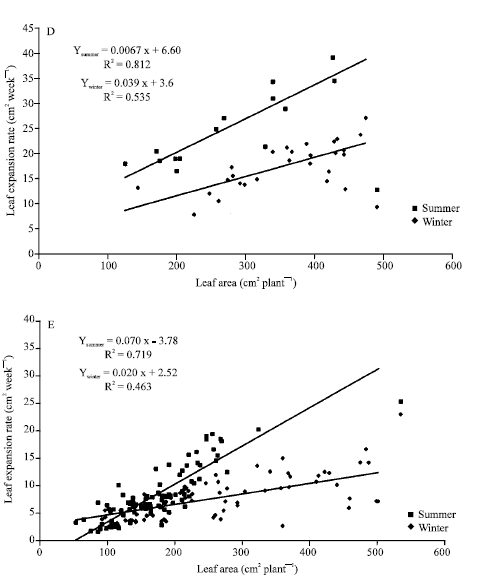 | |
| Fig. 5: | Leaf expansion rate vs. total leaf area for Aglaonema commutatum (A), Dieffenbachia maculata (B), Philodendron ‘Burgundy’ (C), Philodendron peruvianum (D) and Syngonium podophyllum (D) ornamental plants grown under two contrasting seasons |
There was only a small correlation between the root:shoot ratio and total dry weight (g plant-1) for Ph. peruvianum, while the correlation coefficients for A. commutatum, D. maculata, Ph. ‘Burgundy’ and S. podophyllum are quiet low (Table 1).
Extremely lower correlation coefficients for photosynthetic rate (μmoL CO2 m-2 sec-1) and leaf thickness (μm leaf-1) vs. single leaf area (cm2 leaf-1) or photosynthetic rate (μmoL CO2 m-2 sec-1) vs. chlorophyll concentration (mg m-2) for A. commutatum, Ph. ‘Burgundy’ and Ph. peruvianum harvested at the end of the summer or winter season were found (Table 2).
DISCUSSION
Foliage plants comprise a group of plants where only the vegetative growth and plant morphogenesis are of interest. Since flowering can be excluded, the success of producing saleable plants in short time depends on the maintenance of high photosynthetic rates throughout the cultivation period.
One of the problems inherent in making photosynthetic comparisons, which are concurrent with changes in leaf morphology, is in determining a valid basis of expression of photosynthetic rate. Araus et al. (1986) have indicated that differences in maximum Pn between species are even more pronounced when they are expressed on the basis of chlorophyll rather than leaf area; there is a poor correlation between chlorophyll content and maximum Pn, per unit leaf area, for different sun and shade species. Positive correlation between maximum Pn, determined at normal CO2 pressure and leaf thickness is often found among leaves of the same species. However, the present results showed a poor correlation between single leaf area, photosynthetic rate, chlorophyll content and leaf thickness for A. commutatum, Ph. ‘Burgundy’ and Ph. peruvianum (Table 3) grown under optimal (summer) or suboptimal (winter) environments. One possible explanation of the negative relationship between leaf size and photosynthetic capacity is that genotypes with large leaves, such as the Araceae ornamental tested, may have more severe mutual shading which would reduce their photosynthetic capacity.
Only few studies have focused on the relationship between photosynthetic rates (Pn) and plant productivity or growth (Zelitch, 1982; Ottosen, 1990). Results from Table 3 are in agreement with the apparent paradox in lack of correlation between measurement of single-leaf photosynthesis and growth rate. Short-term measurements of photosynthesis may not be a good indicator of the long-term growth effects. Over time, acclimatization to a changing environment takes place in plants and short-term measurements of photosynthesis will often overestimating the effect of, for example, increasing light levels during summer. Photosynthetic measurements as a means of gaining insight into the optimal growing conditions for plants should therefore be used with care.
Accurate estimation of leaf area is vital for evaluation of plant productivity. Destructive harvesting of plants followed by sampling of leaves is the most accurate means to measure area, but is not suitable for applications requiring repeated measurements on the same plant. In such case, the areas of intact leaves can be estimated based on empirically-derived relationships between area and measurements of leaf size and shape (Goenaga and Singh, 1996). The regression lines obtained from the measured values of w (leaf width) and l (leaf length) used to estimate leaf area from A. commutatum, D. maculata, Ph. ‘Burgundy’, Ph. peruvianum and S. podophyllum showed high correlation coefficients (0.98 to 0.99) as was indicated in Table 1.
| Table 3: | Fitted lines and correlations coefficients for three ornamental shade plants grown under two contrasting seasons |
 | |
Plant growth and development is sustained through photosynthetic carbon fixation and the subsequent translocation of photoassimilate to growing tissues or storage organs. Mature leaves are a source tissue because there is a net export of photoassimilate, generally sucrose, from them to the sink organs, which include roots and actively growing meristems. The value of a leaf, to the plant that bears it, is the contribution that it makes to the fitness of the plant (Di Benedetto and García, 1992). The cost of producing a leaf is the investment of resources of carbon, nitrogen, etc.; the interest that is gained from this investment is the carbon fixed by the leaf in photosynthesis. Results plotted in Fig. 2 and 3 are in agreement with this general view; however, the returns for Ph. ‘Burgundy’ would be greater than for A. commutatum, D. maculata, S. podophyllum and its related specie Ph. peruvianum according as plant size increased.
During the monopodial phase, as the leaf size increases from one leaf to the next, the apparent phyllochron decreases. The growth rate is always controlled by the source-sink balance within the plant. The source consists of photosynthetically active leaves and stored reserves and the sink is made of growing organs. The decrease in the apparent phyllochron means that the increase in the assimilate source is greater than the increase in the sink (Dufour and Guerin, 2003). The leaf expansion rates for all ornamental Araceae plants tested are in agreement with earlier studies but the response are related to plant material used (Philodendron ‘Burgundy’ showed the higher increased in reinvestments) and season (Fig. 5).
Differences in competitive abilities may be due to traits such as leaf area, plant height and mass. In particular, initial plant size, expressed as total plant mass, may influence competitive ability (Di Benedetto et al., 2005). One factor that may contribute to variation in competitive ability is growth rate (Gerry and Wilson, 1995). According to the model of Samson and Werk (1986), reproductive allocation is expected to increase allometrically with increasing plant size (Mendez and Obeso, 1993). Figure 1 and 2 are in agreement with these authors, but, although it is expected that plastic responses in resource allocation should be common in species inhabiting heterogeneous environments, the five species used are shade plants which have evolved under homogeneous tropical and subtropical rainforests.
The distribution of biomass among plant organs is not fixed. It is affected by the environment, habit of the plant, life span of the plant and, competitive interactions. Although investment in root system was low: 11.3; 12.5; 33.1; 17.5 and 13.2% of the total dry weight (data not shown) for A. commutatum, D. maculata, Ph. ‘Burgundy’, Ph. peruvianum and S. podophyllum respectively, root:shoot ratio (Table 2) showed an erratic response according plant size (estimated as total dry weight) increased. The distribution of assimilate within the plant is regulated by source-sink interactions. Sources are exporters of assimilates and sinks are net importers of assimilate. The major sources in vegetative plants are the mature leaves as showed the present results. While the acquisition of resources by a plant provides the necessary substrates for growth (primarily C and N), it is the allocation of these resources at the whole-plant level that ultimately determines growth rate and final size.
One of the few integrating principles related to allocation is the hypothesis of balanced root and shoot activity. It has been indicated (Poorter and Remkes, 1990) that fast-growing species are more oriented to maximize shoot functioning, whereas the slow growers tend to maximize root functioning. The results plotted in Table 2 are not in agreement with these suggestions perhaps because water and nutrient supply would not be limited growth factor under greenhouse commercial facilities such as happened in the primitive wild environment.
By other hand, it has been indicated that the carbon partitioning between leaf area expansion and growth of non-photosynthesizing tissues becomes increasingly evident with the age of the individual. Growth rates are dependent on leaf area index, but as leaf area increases, the growth rate reaches a maximum value and thereafter, it may decline (Schulze, 1982). There would be not the case for A. commutatum, D. maculata, Ph. ‘Burgundy’, Ph. peruvianum and S. podophyllum under greenhouse facilities (Table 2 and Fig. 5).
In summary, saleability of tropical ornamental foliage plants and grower’s profit demands high plant quality and productivity. The results from this study showed that the more a plant invests in leaf area, the higher the total carbon gain and the faster growth will be. Most foliage plants have their origins in the tropics and require relatively high night temperatures to sustain rapid growth (Poole and Conover, 1981); a significant change in growth rates would be associated to environmental seasonal influences. By other hand, estimation of leaf area through linear dimensions such as length and width of the leaf lamina (Table 1) and the close correlations between total leaf area and total dry weight (Chanda et al., 1985) with plant size (Fig. 1 and 2) would be an appropriate method and a useful tool for foliage ornamental productivity crop modelling. The knowledge of the whole plant physiology is necessary in order to devise a greenhouse management program which is close to optimal with respect to growth.
ACKNOWLEDGEMENT
This study was partially supported by the UBACyT 2004-2007 Program (G 031).
REFERENCES
- Bazzaz, F.A., N.R. Chiariello, P.D. Coley and L.F. Pitelka, 1987. Allocating resources to reproduction and defense. BioScience, 37: 58-67.
Direct Link - Di Benedetto, A. and A.F. Garcia, 1992. Adaptation of ornamental aroids to their light environments. I: Spectral and anatomical characteristics. J. Hortic. Sci., 67: 179-188.
Direct Link - Moran, R., 1982. Formulae for determination of chlorophyllous pigments extracted with N,N-dimethylformamide. Plant Physiol., 69: 1376-1381.
CrossRefDirect Link - Ottosen, C.O., 1990. Growth versus net photosynthesis in clones of Ficus benjamina. HortScience, 25: 956-957.
Direct Link - Poorter, H. and C. Remkes, 1990. Leaf area ratio and net assimilation rate of 24 wild species differing in relative growth rate. Oecologia, 83: 553-559.
CrossRefDirect Link - Soyza, A.G., A.C. Franco, R.A. Virginia, J.F. Reynolds and W.G. Whitford, 1996. Effects of plant size on photosynthesis and water relations in the desert shrub Prosopis glandulosa Fabaceae. Am. J. Bot., 83: 99-105.
Direct Link - Vidal, D., E. Griera, P. Marin and J. Sabido, 1990. Anatomical and physiological acclimation of Fatsia japonica leaves to irradiance. Am. J. Bot., 77: 1149-1158.
Direct Link