
M. Dejam
Department of Horticultural Science, College of Agriculture, Shiraz University, Shiraz, Iran
M. Khosh-Khui
Department of Horticultural Science, College of Agriculture, Shiraz University, Shiraz, Iran
A. Shekafandeh
Department of Horticultural Science, College of Agriculture, Shiraz University, Shiraz, Iran
International Journal of Agricultural Research
Year: 2006 | Volume: 1 | Issue: 1 | Page No.: 14-19







ABSTRACT
Experiments were conducted to study the in vitro propagation of Bakrai using epicotyl explants. Explants were excised form in vitro grown seedlings, cultured on a basic MS medium supplemented with various concentrations of benzyladenine (BA) and naphthaleneacetic Acid (NAA). The highest percentage of explants producing normal adventitious shoots (83.3%) was obtained on a medium MS medium containing 2 mg L-1 BA. The maximum number of adventitious buds (20.33) produced in 4 mg L-1 BA, but most of these buds remained quiescent. Addition of 0.5 mg L-1 NAA increased shoot number and shoot length but higher NAA concentrations had an inhibitory effect on shoot regeneration, particularly in 1 mg L-1 BA concentration. The highest number of shoots (5.5) and longest shoots (9.68 mm) were obtained in concentrations of 2 and 1 mg L-1 BA plus 0.5 mg L-1 NAA, respectively. The shoots were rooted on a basal medium with half-strength MS salts and various concentrations of indolebutyric acid (IBA) (0.5-10 mg L-1 ). The highest percentage of rooting (91.67%) and the maximum number (1.71) and length (20.33 mm.) of roots were obtained with 10 mg L-1 IBA. The plantlets were transferred to pots containing an equal mixture (v/v) of sand and loamy soil and successfully acclimatized to natural environment with a survival rate of 80%.
PDF Abstract XML References
How to cite this article
M. Dejam, M. Khosh-Khui and A. Shekafandeh, 2006. Adventitious Bud Induction and Plant Regeneration in Epicotyl Segments of Bakrai (Citrus reticulata BlancoxC. limetta Swing.). International Journal of Agricultural Research, 1: 14-19.
DOI: 10.3923/ijar.2006.14.19
URL: https://scialert.net/abstract/?doi=ijar.2006.14.19
DOI: 10.3923/ijar.2006.14.19
URL: https://scialert.net/abstract/?doi=ijar.2006.14.19
INTRODUCTION
Citrus fruits are one of the most important fruit crops of the world planted in tropical and subtropical regions. The world production of citrus fruits in 1998 was over 100 million tones (Jackson and Looney, 1999). Most of citrus species and hybrids are highly polyembryonic due to natural occurrence of nucellar embryony and reproducing true-to-type plants by seeds. The main citrus rootstocks are therefore usually propagated by growing open-pollinated seeds (Wutscher, 1979). However, micropropagation and plant regeneration of citrus species are of interest because by in vitro techniques the number of plants produced would not be limited by seed supply and also more uniform plant populations might be produced (Moore, 1986). Furthermore, an efficient system of regeneration through organogenesis is the first step for genetic transformation of these species (Gmitter et al. 1992).
In vitro plant regeneration by direct organogenesis has been reported for different citrus species and from several tissues including roots (Sauton et al., 1982; Burger and Hackett, 1986; Bhat et al., 1992), stem segments (Barlass and Skene, 1982; Moore, 1986; Duran-Vila et al., 1989; Perez-Molphe-Balch and Ochao-Alejo,1997; Ghorbel et al., 1998; Kobayashi et al., 2003), epicotyls (Edriss and Burger,1984; Burger and Hackett, 1986; Sim et al., 1989; Maggon and Singh, 1995; Moreira-Dias et al., 2000; Bespalhok-Filho et al., 2001; Dejam et al., 2001; Almeida et al., 2002; Costa et al., 2004) and hypocotyls (Maggon and Singh, 1995; Normah et al., 1997). In most cases, benzyladenine (BA) has been required for shoot regeneration, although the optimum concentration of BA has been different for various species and explants. The rooting percentage of plantlets obtained by adventitious shoots has been different for various species and with different auxins and their concentrations. Bakrai which is a natural hybrid between mandarin and sweet lime is used as a citrus rootstock in Iran and some other countries. Bakrai is commonly propagated by seeds or hardwood cuttings. However, direct organogenesis can be used for both mass propagation and improvement of Bakrai. In this investigation, for the first time, successful plant regeneration from epicotyl segments of Bakrai is reported.
MATERIALS AND METHODS
The ripe fruits were collected form open-pollinated trees of Bakrai in Darab, south of Iran in 2002 and brought to Tissue Culture Lab of Iranian Research Organization of Science and Technology, Shiraz Branch Iran. Seeds were extracted from fruits, washed by water and both seed coats were removed. Then surface sterilized by immersion in a 0.5% sodium hypochlorite solution for 10 min and rinsed 3 times with sterile distilled water. The Murashige and Skoog (1962) (MS) basal medium supplemented with 3% (w/v) sucrose was used in different experiments. The pH of media was adjusted to 5.7±0.1 before the addition of 27.5 g L-1 agar. The media were dispensed as 15 mL aliquots in 25x150 mm culture tubes, capped and sterilized by autoclaving for 30 min at 121°C and 1.5 kg cm-2 pressure. All cultures were maintained at 27±2°C and unless otherwise stated were incubated at 16 h light per day with 1000 lux intensity. Decoated seeds were cultured in basal medium in continuous darkness for 10 days and then transferred to light conditions for 20 days before use.
For shoot regeneration studies, the 0.5 cm long epicotyl explants were excised from in vitro seedlings and cultured horizontally in basal medium supplemented with concentrations of 1, 2 and 4 mg L-1 BA in combination with 0, 0.5 and 1 mg L-1 napthaleneacetic Acid (NAA). Cultures were incubated in darkness for 20 days and then transferred to light for 40 days. After 20 days, the number of responding explants and adventitious buds and at the end of incubation period, the number and length of produced shoots were recorded.
For rooting studies, the 8-12 mm shoots were placed vertically in a half-strength basal medium supplemented with IBA concentrations of 0.5, 1, 2, 5 and 10 mg L-1 . The cultures were incubated in darkness for 15 days and then transferred to light for 30 days. The percentage of rooting and number and length of roots per explant were recorded after 45 days. For acclimatization, the rooted plantlets were transferred to pots containing an equal mixture of sand and loam soil and irrigated with one-eight concentration of MS salts for 10 days. The pots were enclosed by clear polyethylene bags to minimize the moisture loss. The bags removed gradually to facilitate the conditioning of plantlets to natural environment.
A Complete randomized design with 10 replicates (3 explant/rep.) was used for all experiments. The data were analyzed and the means compared by Duncan’s new multiple range test at 5% probability using SAS software.
RESULTS
Adventitious Bud and Shoot Regeneration
Two weeks after culture, the adventitious buds were visible at the cut ends of epicotyl segments. In most cases, they were arose directly from explants, although, in some cases a small amount of white callus was also produced. Transferring of cultures to light conditions, enhanced shoot regeneration form these buds (Fig. 1A).
The percentage of responding explants was different among growth regulator treatments (Table 1). In basal medium, the response of explants was very low. The highest percentage of explants with adventitious buds was obtained in concentration of 2 mg L-1 BA.
With increasing BA concentrations, the number of adventitious buds increased and there were significant differences among treatments (Table 1). The maximum number of adventitious buds was produced in 4 mg L-1 BA concentration that showed a significant difference with other treatments.
Although the maximum number of buds produced with 4 mg L-1 BA, most of these buds had no growth. In lower BA concentrations, the frequency of shoot elongation was higher. The maximum number of adventitious shoots was obtained with concentration of 2 mg L-1 BA + 0.5 mg L-1 NAA (Table 1).
There were no significant differences in shoot length between 1 and 2 mg L-1 BA, but the length of shoots was comparatively shorter in concentration of 4 mg L-1 BA. The longest shoots were produced in medium with 1 mg L-1 BA plus 0.5 mg L-1 NAA (Table 1).
Rooting and Acclimatization
Root initiation usually occurred near the cut surface of shoot explant’s basal ends (Fig. 1B). They were visible 15-20 days after culture. Increasing IBA concentrations increased the percentage of rooting and the number and length of roots.
| Table 1: | Effects of plant growth regulator treatments on adventitious shoot formation in epicotyl cultures of Bakrai |
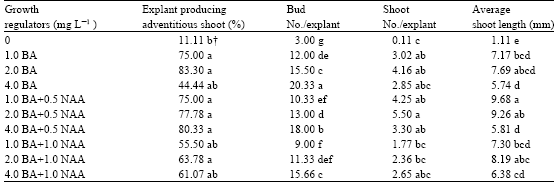 | |
| †Means in each column with similar letter(s) are not significantly different at 5% level of probability using DMRT | |
| Table 2: | Effects of auxin concentrations on rooting of Bakrai shoots |
 | |
| †Means in each column with similar letter(s) are not significantly different at 5% level of probability using DMRT | |
 | |
| Fig. 1: | Stages in the development of Bakrai plantlets obtained from epicotyl segments. (A) Adventitious shoots produced on MS+BA (2 mg L-1) + NAA (0.5 mg L-1), (B) Rooting of adventitious shoots on 1/2MS+BA (10 mg L-1), © Young Bakrai plantlets transferred to soil |
In the concentration of 10 mg L-1 IBA, the mentioned characters were significantly higher than those in lower IBA concentrations (Table 2).
The plantlets were acclimatized at 27±2°C with 16 h photoperiod at 4000 lux light intensity for 4 weeks. The gradual removal of plastic bags caused hardening and adaptation of plantlets (Fig. 1C). About 20% of plantlets were lost during acclimatization.
DISCUSSION
In this investigation, the response of epicotyl explants of Bakrai enhanced with addition of BA to basal medium and maximum response was obtained with 2 mg L-1 BA concentration. This is in agreement with results obtained with epicotyl and hypocotyl explants of sweet orange (Maggon and Singh, 1995). In epicotyl explants of Citrus mitis, the highest response has been reported with concentrations of 0.5 or 1 mg L-1 BA and higher concentrations showed inhibitory effects (Sim et al., 1989). On the other hand, in internodal segments of Mexician lime and mandarin, the optimum concentration of BA was 7.5 mg L-1 (Perez-Molphe-Balch and Ochao-Alejo, 1997). These differences may be due to genotypic variations, type and age of explants and even the position of explants on medium.
Although the maximum number of buds produced with 4 mg L-1 BA, most of these buds remained quiescent. It seems that high concentrations of BA had an inhibitory effect on shoot elongation (Dejam et al., 2002). Addition of 0.5 mg L-1 NAA, increased the number and length of adventitious shoots in all BA concentrations, but 1 mg L-1 NAA decreased the number and length of shoots especially in 1 mg L-1 BA. Edriss and Burger (1984) found that 0.1 to 1 mg L-1 NAA was essential for shoot regeneration in epicotyl segments of ‘Troyer’ citrange, while Moore (1986) reported an inhibition in shoot formation by NAA at these concentrations, when BA concentration was low. In another investigation, the stimulatory effects of low NAA concentrations (0.08 to 0.4 mg L-1) and inhibitory effect of higher NAA concentration (1 mg L-1) on shoot formation in epicotyl explants of ‘Troyer’ citrange has been reported Moreira-Dias et al. (2000).
For rooting the Bakrai microshoots, the best results was obtained with IBA. Although the various concentrations of NAA was also tried, NAA caused poor rooting and callus production in basal ends of shoots was noticed. Rooting was achieved, when IBA was added to the medium in spite of various reports of better results with NAA in other citrus species (Barlass and Skene, 1982; Kitto and Young, 1981; Normah et al. 1997). However, recently low rooting percentage with NAA has been reported in lemon (Kotsias and Roussos, 2001) and sweet orange (Dejam et al., 2002) microshoots. Increasing of IBA concentrations up to 10 mg L-1 increased rooting percentage, root number and length in Bakrai. Increases in rooting percentage and root number have been reported with increasing auxin concentrations in lemon (Kotsias and Roussos, 2001) and ‘Troyer’ citrange (Moreira-Dias et al., 2000) shoots, but the length of roots has been decreased in these cases. On the other hand, in mandarin and lemon shoots, it has been reported that with increasing auxin concentrations, both the percentage of rooting and root number and length have been decreased (Singh et al., 1994). These differences may be due to genotypic variations, different shoot regeneration pathways and/or differences in the sensitively of microshoots to auxins. In conclusion, the best results can be obtained with culturing epicotyl explants of Bakrai on media with 1 and 2 mg L-1 BA plus 0.5 mg L-1 NAA.
REFERENCES
- Almeida, W.A.B., F.A.A. Mourao-Filho, B.M.J. Mendes and A.P.M. Rodrigues, 2002. In vitro organogenesis optimization and plantlet regeneration in Citrus sinensis and C. limonia. Scientia Agricola, 59: 35-40.
Direct Link - Barlass, M. and K.G.M. Skene, 1982. In vitro plantlet formation from Citrus species and hybrids. Scientia Hort., 17: 333-341.
Direct Link - Bespalhok-Filho, J.C., A.K. Kobayashi, L.F.P. Pereira, Z. Hissano and L.G.E. Vieira, 2001. In vitro adventitious shoot regeneration from sweet orange using epicotyl sections. Crop Breed. Applied Biotechnol., 1: 27-34.
Direct Link - Bhat, S.R., I. Chitralekha and K.P.S. Chandel, 1992. Regeneration of plants from long-term root culture of lime, Citrus aurantifolia (Christm.) swing. Plant Cell Tissue Organ Cult., 29: 19-25.
Direct Link - Costa, M.G.C., V.S. Alves, E.R.G. Lani, P.R. Mosquim, C.R. Carvalho and W.C. Otoni, 2004. Morphogenic gradients of adventitious bud and shoot regeneration in epicotyl explants of citrus. Scientia Hortic., 100: 63-74.
Direct Link - Duran-Vila, N., V. Ortega and L. Navarro, 1989. Morphogenesis and tissue culture of three citrus species. Plant Cell Tissue Organ Cult., 16: 123-133.
Direct Link - Edriss, M.H. and D.W. Burger, 1984. In vitro propagation of Troyer citrange from epicotyl segments. Scientia. Horticulturae, 23: 159-162.
Direct Link - Ghorbel, R., L. Navarro and N. Duran-Vila, 1998. Morphogenesis and regeneration of whole plants of grapefruit (Citrus paradisi), sour orange (C. aurantium) and alemow (C. macrophylla). J. Hortic. Sci. Biotechnol., 73: 323-327.
Direct Link - Kobayashi, A.K., J.C. Bespalhok L.F.P. Pereira and L.G.E. Vieira, 2003. Plant regeneration of sweet orange (Citrus sinensis) from thin sections of mature stem segments. Plant Cell Tissue Organ Cult., 74: 99-102.
Direct Link - Kotsias, D. and P.A. Roussos, 2001. An investigation on the effect of different plant growth regulating compounds in in vitro shoot tip and node culture of lemon seedlings. Scientia Horticulturae, 89: 115-128.
CrossRefDirect Link - Maggon, R. and B.D. Singh, 1995. Promotion of adventitious bud regeneration by ABA in combination with BAP inn epicotyl and hypocotyl explants of sweet orange (Citrus sinensis L. Osbeck). Scientia Hortic., 63: 123-128.
Direct Link - Moreira-Dias, J.M., R.V. Molina, Y. Bordon, J.L. Guardiola and A. Garcia-luis, 2000. Direct and indirect shoot organogenic pathways in epicotyl cuttings of Troyer citrange differ in hormone requirement and in their response to light. Ann. Bot., 85: 103-110.
Direct Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Normah, M., N.S. Hamidah and F.D. Ghani, 1997. Micropropagation of Citrus halimii-an endangered species of South-East Asia. Plant Cell Tissue Org. Cult., 50: 225-227.
Direct Link - Perez-Molphe-Balch, E. and N. Ochao-Alejo, 1997. In vitro plant regeneration of Mexican lime and mandarin by direct organogenesis. HortScience, 32: 931-934.
Direct Link - Singh, S., B.K. Ray, S. Bhattacharyya and P.C. Deka, 1994. In vitro propagation of Citrus reticulata Blanco and Citrus limon Burm.f. HortScience, 29: 214-216.
Direct Link