
Shruti Mishra
Biochemistry, Faculty of Science, Banaras Hindu University, Varanasi-221 005, India
R. S. Dubey
Biochemistry, Faculty of Science, Banaras Hindu University, Varanasi-221 005, India
International Journal of Agricultural Research
Year: 2006 | Volume: 1 | Issue: 2 | Page No.: 122-141







ABSTRACT
Plants take up heavy metals from their surroundings via a number of different plasma membrane localized transporters. Inside the plant system toxicity of heavy metals is manifested by an array of physiological and metabolic disturbances if metal detoxification processes are delayed or not efficient. Immobilization of heavy metals by root exudates and in some cases mycorrhizal association constitute the extracellular mode of detoxification whereas chelation of heavy metals by phytochelatins, metallothioneins, organic acids and amino acids, induced synthesis of proline and synthesis of stress proteins are implicated in preventing metal accumulation in fully reactive form in the cytosol. Plant cells have developed sophisticated toxic metal sensing proteins that transmit signals to activate expression of metal detoxification and homeostasis genes in order to cope with the elevated heavy metal concentrations. In transducing signals of heavy metal origin involvement of second messengers viz., Ca2+, jasmonic acid and H2O2 as well as participation of ethylene and salicylic acid are likely. This review aims to give succinct overview of plant metal uptake and detoxification mechanisms.
PDF Abstract XML References
How to cite this article
Shruti Mishra and R. S. Dubey, 2006. Heavy Metal Uptake and Detoxification Mechanisms in Plants. International Journal of Agricultural Research, 1: 122-141.
DOI: 10.3923/ijar.2006.122.141
URL: https://scialert.net/abstract/?doi=ijar.2006.122.141
DOI: 10.3923/ijar.2006.122.141
URL: https://scialert.net/abstract/?doi=ijar.2006.122.141
Introduction
Soil contamination with heavy metals has become a world–wide problem particularly in areas with high anthropogenic pressures leading to losses in agricultural yield as well as hazardous health effects when these metals enter the food chain (Frankenberger, 2002; Jonak et al., 2004).
Plants require relatively small number of elements for their growth and survival. Natural soils however contain many other elements that are non-essential and in spite of the selectivity of root cell membranes, may be detected in plant tissues in trace amounts. Many of these elements, especially metals like Cd, Pb, As, Hg can be highly toxic. In addition, required micronutrients such as Cu, Ni and Zn can also be toxic to the plants when present in higher concentrations in the soil environment. Accumulation of heavy metals in the soil influences soil properties and adversely affects the establishment and growth of plants whose roots initially develop in the most contaminated layer (Bhattacharjee and Mukherjee, 2004; Jha and Dubey, 2004). Uptake of metals by plants occurs via the root and the extent of absorption mostly depends on the ionic potential of the element concerned. Sometimes metals can be absorbed actively or passively or undergo accumulation along with a macronutrient cation. There occurs a competition among the metals for chelation, movement to the root and uptake at the root-tip. Available data suggest that heavy metals if not detoxified rapidly enough, may trigger multiple deteriorative events in plants by altering major physiological and biochemical processes, ultimately leading to visible plant injuries and yield losses (Carrier et al., 2003; Jha and Dubey, 2004; Kukier and Chaney, 2004).
During the past two decades conserted research efforts have addressed the biology of heavy metal toxicity at the whole plant, cellular and subcellular levels. A detail understanding of the uptake of heavy metals by plants, components associated with metal tolerance and signal transduction pathway in presence of metal is essential in order to produce metal accumulating and metal tolerant plant species by using genetic engineering techniques. This article reviews our current status of knowledge related to the uptake of metals, the strategies adopted by plants in their detoxification as well as the possible involvement of signal molecules in metal detoxification processes mainly in relation to Cd, Pb, Hg, As, Ni and the metal Al. Although aluminium is not a heavy metal, but its mode of action and detoxification processes involved to reduce its toxicity are of particular concern due to potential phytotoxic nature of aluminium especially in acid soils.
Uptake of Heavy Metals
Root cell walls initially bind metal ions from the soil and thereafter via high affinity binding sites and plasma membrane localized transport systems; metal ions are taken up across plasma membrane. Generally uptake of metal ions occurs through secondary transporters such as channel proteins and/or H+-coupled carrier proteins. Uptake of cations through secondary transporters is facilitated by the membrane potential of the plasma membrane, which is negative on the inner side of the membrane (Hirsch et al., 1998). A number of different types of metal transporters are implicated in plants and often more than one transport system exists for one metal (Williams et al., 2000). Summary of putative Cd, As, Ni and Zn transporters identified in plants to date has been presented in Fig. 1.
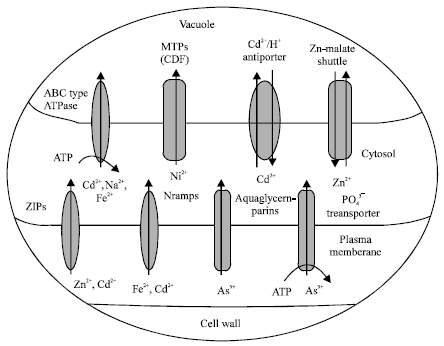 | |
| Fig. 1: | Transporters of heavy metals Cd, As(V), As(III), Ni and Zn |
In uptake and transport of Zn2+ and Cd2+, ZNT (zinc-iron permease [ZIP] gene homolog) proteins have been implicated. The histidyl area of the variable region of these transport proteins is involved in metal binding and their subsequent transport (Pence et al., 2000). The metal ion Cd2+ can also enter the plant cell by transporters for essential cations such as Fe2+ (Thomine et al., 2000). In Arabidopsis, AtNramps (Natural resistance associated macrophage proteins) genes encode the metal transporters, which transport both mineral nutrient Fe and toxic metal Cd (Thomine et al., 2000). Lead is taken up by plant roots primarily through passive absorption (Tung and Temple, 1996), although certain proteins have been identified that mediate cross membrane movement of Pb. In tobacco a protein NtCBP4 has been identified which is involved in metal uptake across plasma membrane. Expression of NtCBP4 in transgenic tobacco confers Pb-hypersensitivity (Arazi et al., 2000). Plants take up arsenate (AsV) via phosphate transporter. A competition occurs between phosphate and arsenate at the uptake level for the same transport system as evidenced from several studies in plant species like Oryza sativa (Abedin et al., 2002), Brassica juncea (Pickering et al., 2000). Arsenate accumulation follows Michaelis-Menten kinetics at a concentration range that coincides with the level of activity of the high affinity phosphate uptake system (Abedin et al., 2002). A class of multifunctional channels called aquaglyceroporins has been identified in plants, which transport neutral organic solutes such as urea and glycerol. Transport of arsenite (AsIII) via these aquaglyceroporins has been reported in plants (Meharg and Jardine, 2003).
No specific Ni-uptake system has been characterized in higher plants. However, two distinct high affinity systems have been described in prokaryotes- the single component Ni2+ permeases, which belong to the nickel/cobalt transporter (NiCoT) family and the Nik systems, which belong to ABC transporter family (Eitinger and Mandrand-Berthelot, 2000). The absorption of organic and inorganic Hg from soil by plants is low (Lodenius, 1990) and after uptake by plants, restricted transport of Hg is observed through phloem (Panda Kamal et al., 1992). However, air-borne mercury, in certain cases, is absorbed by plant leaves (Wojciechowska-Mazurek et al., 1995).
Extracellular Avoidance of Metal Build-up
At extracellular level plants respond to metals either by immobilizing them with their root exudates or some mycorrihzal association helps them in performing this task. In many plant species primary barrier against entry of heavy metals into plant roots operates at the root level. Cd can be immobilized by cell wall (Leita et al., 1996) and extracellular carbohydrates (Wagner, 1993). In roots and leaves of bush bean, Cd ions mostly seem to bind pectic sites and histidyl groups of the cell wall (Leita et al., 1996). Possible involvement of histidine and citrate has been suggested in root exudates that reduces Ni-uptake and helps in Ni-detoxification (Salt et al., 2000).
In several bean species differences in Al tolerance have been attributed to the capacity of roots to exude citric acid, a strong Al chelator (Ma, 2000). In cereals also better tolerance to Al toxicity is associated with root exudation of citric acid, malic acid, succinic acid, oxalic acid and other organic acids (Bertrand and Guary, 2002). Activation of organic acid efflux occurs rapidly without any measurable delay after exposure to Al in several plant species (Ryan et al., 2001). Efflux of citrate and malate from plant roots in response to Al occurs by activation of anion-channels located in the plasma membrane (Ryan et al., 2001). In lower plants, like algae, it has been shown that metals can be trapped by polysaccharides, alginates and other slimes secreted by the algae (Lee and Volesky, 1997).
There are accumulating evidences in support of mycorrhizal involvement in host plant metal detoxification more specially the ectomycorrhizal association (Blaudez et al., 2000; Jentscheke and Godbold, 2000). The fungi employ the same strategy for metal detoxification as adopted by higher plants i.e., binding to extracellular materials or sequestration in the vacuolar compartment (Blaudez et al., 2000).
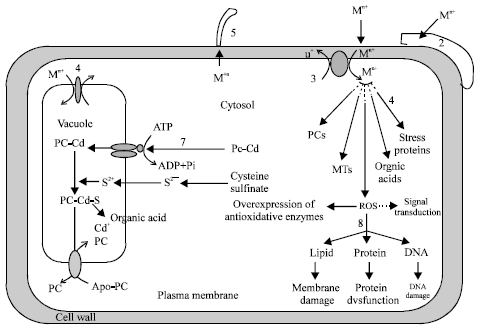 | |
| Fig. 2: | Cellular mechanisms proposed to be involved in metal uptake, sequestration and detoxification in plants. 1. Extracellular avoidance of metal buildup through immobilization by root exudates. 2. Ectomycorrhizal association restricts metal movement to roots. 3. Metal ions are taken up by plant roots through channel proteins and/or H+-coupled carrier proteins. 4. From cytosol metals are transported and accumulated in vacuoles, the events are aided by vacuolar electrogenic proton fluxes. 5. Glandular trichomes and epidermal structures (hydropotes) sequester metals in leaves. 6. Metal ions in cytosol can be detoxified via these routes. 7. Mechanisms involved in Cd chelation and compartmentalization in the vacuole. 8. Metal ions escaped from the complexation damage cellular macromolecules via the production of ROS |
Ectomycorrhizas restrict metal movement to host roots by exclusion processes (Jentsckeke and Godbold, 2000). Some mycorrhization stimulates the phenolic defense system in mycorrhizal symbiosis (Schützendübel and Polle, 2002). Cellular mechanisms proposed to be involved in metal uptake, sequestration and its detoxification in plants have been outlined in Fig. 2.
Intracellular Detoxification Pathways
Phytochelatins
Higher plants, algae and fungi can respond to exposure of several metals by the synthesis of thiol containing metal binding peptides called phytochelatins (PCS) (Cobbett, 2000). The immobilized metals in form of metal phytochelatin complexes are very stable and less toxic than free metal ions. PCS are considered to be an important component of the metal detoxifying mechanisms of higher plants (Zenk, 1996; Hartley-Whitaker et al., 2001; Küpper et al., 2004). PCS are derived from glutathione (GSH) and possess the general structure (γ-Glu-Cys)n-Gly where n = 2-11 (Zenk, 1996; Cobbett, 2000; Goldsbrough, 2000). Their biosynthesis occurs due to the trans-peptidation of γ-glutamyl-cysteinyl dipeptides from GSH by the action of constitutively present enzyme PC synthase that is activated due to heavy metal ions. When PCS are synthesized in sufficient amount to chelate metal ions, the enzyme is turned off and the reaction is terminated (Chen et al., 1997). The unusual bonding between glutamate and cysteine i.e., carboxyamide linkage in phytochelatin suggests that they are not synthesized on ribosomes and are thus not a direct gene product but are enzymically synthesized polypeptides (Harada et al., 2004). The cysteine residues of PCS ensure metal coordination via their thiol group and the high percentage of glutamic acid residues makes PCS extremely water soluble (Bertrand and Guary, 2002). The N-terminal and downstream γ-peptidyl bonds of PCS probably serve to protect thiol peptides from general protease action but not from the specific action of γ-glutamyl transpeptidases during the salvage process (Satofuka et al., 2001).
PCS have been classified as class III metallothioneins and are induced in all autotrophic plants analyzed so far. The biosynthesis of PCS is induced by many metals like Cd, Hg, Ag, Cu, Ni, Au, Pb and Zn; however, Cd is the strongest inducer (Zenk, 1996). Despite the fact that many metals are able to trigger PC synthesis, only some of them (i.e., Cd, Hg, Pb, Cu, Ag) are able to form complexes with PCS (Bertrand and Guary, 2002). Following Cd exposure, a rapid induction in PC level has been observed in Brassica napus (Carrier et al., 2003). Lead exhibits several coordination numbers when it binds with PCS; the number depends on the length of the PC chain; PCS with longer chain bind Pb more strongly than those with shorter chain length (Piechalak et al., 2002). Arsenate and arsenite both induce the synthesis of PCS (Pickering et al., 2000; Hartley-Whitaker et al., 2001). In Brassica juncea, an As-non-accumulator, As III-thiolate complex has been reported and the thiolate has probably its origin from glutathione or PCS (Pickering et al., 2000). Maitani et al. (1996) reported rapid induction of Hg-binding PCS in root cultures of Rubia tinctorum following metal exposure. In addition, a number of structural variants of PCS, such as (γ-Glu-Cys)n-β-Ala, (γ-Glu-Cys)n-Ser and (γ-Glu-Cys)n-Glu have been identified in certain plant species (Cobbett, 2000).
Considerable efforts have been made to study the genes encoding PC synthases as well as catalytic mechanism of PC synthases in higher plants. PC synthase like polypeptides are of widespread occurrence in plants and a catalytic homologue of this enzyme has been identified in prokaryotes (Tsuji et al., 2004). The enzyme PC synthase mediates a bisubstrate reaction in which the thiol group(s) of at least one of the substrate is blocked, usually, but not necessarily through the formation of metal thiolate. The chain extention proceeds along CπN with cleavage of the Cys-Gly peptide bond of the donor (Rea et al., 2004). It is proposed that the strongly conserved N-terminal half of the enzyme is responsible for catalysis and that activation arises from the binding of the metal ions to cysteine residues, within this domain (Ruotolo et al., 2004). This finding is in consistence with the observation that three most extreme Arabidopsis cad1 alleles have amino acid substitutions in this region (Ha et al., 1999). The C-terminal domain of PC synthase contributes stability to the full-length polypeptide and participates in defining the range of metals to which the enzyme is responsive (Ruotolo et al., 2004).
A lot of research efforts are underway to characterize the gene encoding PC synthase in higher plants. Clemens et al. (1999) and Ha et al. (1999) have successfully characterized structural genes of this enzyme in Arabidopsis, wheat and yeast. Two Cd-responsive novel genes coding for the proteins ATMEKK1 and a putative farnesylated protein, with two metal binding motifs have been reported in Arabidopsis that endow marked tolerance to the plants towards Cd (Suzuki et al., 2001).
Plant age dependent differences in the complexation and speciation of heavy metals has also been observed. In Thlaspi caerulescens plants the nature of Cd binding changed depending on the tissue age (Küpper et al., 2004). In young leaves predominant Cd ligands appeared to be sulphur-ligands (e.g., phytochelatins, metallothioneins and other cysteine rich peptides) while in senescing leaves oxygen-ligands were predominantly involved.
Metals when get complexed with PCS are transported to vacuoles. In this way PC-metal complexes isolate the toxic metals from various metal sensitive enzymes in the plant cell cytoplasm and possibly form a more complex aggregation in the vacuole (Cobbett, 2000). The metal-PC complex is transported from the cytosol to the vacuole by either metal/H+ antiporters or ATP-dependent ABC transporters of tonoplast (Salt and Rauser, 1995; Rea et al., 1998). Involvement of a ABC-transporter YCF1 (yeast cadmium factor 1) has been reported in yeast which catalyzes Mg ATP-energized transport of bis(glutathionato)cadmium [Cd.GS2] into vacuole (Li et al., 1997). In S. pombe heavy metal tolerance factor 1 (HMT1) catalyzes the Mg ATP-energized uptake of Cd.PCs and apoPCs into the vacuoles (Ortiz et al., 1995). Similar Mg-ATP dependent transport of PC and Cd-PC complexes have been shown in vacuolar membrane vesicles of oat roots and this transport pathway appears to be an analog of HMT1 of S. pombe (Ortiz et al., 1995). In Cd-hyperaccumulator Thlaspi caerulescens plants about 90% of the total Cd present in mesophyll cells remains localized in vacuoles (Ma et al., 2005). Therefore in these plants vacuolar sequestration has been suggested as an important mechanism of internal detoxification of Cd in leaves (Ma et al., 2005). Vacuolar transportation of As-PC complexes has also been reported in several plant species (Meharg and Hartley Whitaker, 2002; Raab et al., 2004), however, thiol conjugated forms of As(III) have not been observed (Raab et al., 2004). In S. cerevisiae MRP (multidrug resistance-associated protein) homolog, Ycf1p has vacuolar localization and catalyzes sequestration of bis(glutathionato)arsenic [As(GS)3] in the vacuole (Ghosh et al., 1999).
PC-metal complexes not only serve as an important component of intracellular heavy metal detoxification mechanisms in plants by translocating metals across the tonoplast and sequestering in vacuoles but these complexes can also undergo long-distance transport from roots to shoots. A gene, TaPCS1 from wheat, when transgenically expressed in Arabidopsis roots, enhances long distance root-to-shoot Cd2+ transport and reduces Cd-accumulation in roots (Gong et al., 2003). In vacuoles inorganic sulfide and sulfite ions are incorporated in the PC-metal complexes making them more stable (Bertrand and Guary, 2002). Sulfide ions in the phytochelatin complex stabilize the pH at which dissociation of metal occurs (Steffens, 1990). Phytochelatin complexes containing labile sulfur ions are more stable and resistant to proteolytic degradation than those lacking these constituents (Steffens, 1990). In vacuoles, from PC-metal-sulfide complexes metals are liberated and the PC moiety is degraded (Grill et al., 1988). The metals are complexed with vacuolar organic acids while the individual amino acids can again enter into the cytosol (Zenk, 1996).
Besides detoxification of metals, PCS also appear to have certain other important roles in the cell like in maintaining homeostasis of heavy metals (Thomine et al., 2000), in sulfur metabolism (Rauser, 1995) as well as in antioxidative defense of the plant. PCS may have higher antioxidant activity than other well-known antioxidants (Giles et al., 2003). Certain studies have indicated that anthocyanins also have a role in metal sequestration. They can bind metal ions and with the help of ABC-type transporter proteins glutathione-conjugated anthocyanin-metal complexes are then transported to the vacuoles (Marrs, 1996).
Metallothioneins
Metallothioneins (MTs) are cysteine rich gene encoded proteins capable of sequestering metals by forming metal-thiolate clusters. They have molecular weight in the range 5-20 kDa and can be divided into two different classes on the basis of their cysteine content and structure. MTs are proteins typical of vertebrates but MT genes have also been identified in a wide range of plant species (Goldsbrough, 2000). The expression of MTs in plants growing in absence of metal excess has also been observed (Betrand and Guary, 2002).
Introduction of mammalian MT gene into B. compestris, B. napus and N. tabacum confers Cd tolerance by increasing complexation of metal with MT and minimizing translocation of Cd to the shoots (Maiti et al., 1989). A number of genes in plants have been found to encode MT-like proteins having sequence similarity to animal MT proteins. The history of plant MTs dates back to 1987, when the first plant metallothionein protein termed Ec protein (early cysteine-labled) was detected in wheat (Lane et al., 1987). Later, a number of MT-encoding genes were identified in Arabidopsis. A total of eight genes constitute MT-gene family in Arabidopsis (Zhou and Goldsbrough, 1994). MTs have highest complexing affinity for Cu and are induced by exposure to it (Murphy et al., 1997). On Cu treatment, strong induction in MT2 mRNA level has been observed in Arabidopsis. When genes for MT1 and MT2 from Arabidopsis were expressed in an MT-deficient yeast mutant, both genes complemented the mutation and provided a high level of resistance to Cu (Zhou and Goldsbrough, 1994). Similarly, in Cu-sensitive mutants of Arabidopsis it was shown that MT genes could be induced by Cu. A PsMTA gene identified in the roots of P. sativum regulates the synthesis of MT-like proteins (Robinson et al., 1992).
There appears to be similarity between MTs and PCS for complexation with metals as observed in plants such as datura and maize (Rivai et al., 1990). The role of MTs as antioxidants has also been advocated (Wong et al., 2004). A decrease in MT levels could result in increased level of free transition metal ions (Briat, 2002). According to Wong et al. (2004) a rice metallothionein protein OsMT2b appears to function as an ROS scavenger, which depicts the antioxidative role of plant metallothioneins. Similarly, NO-scavenging capacity and involvement in plasma membrane repair appear to be additional roles of MTs (Salt et al., 1998).
Stress Proteins
When subjected to heavy metal treatment, most of the plants trigger the synthesis of certain sets of novel proteins. Induction of protein synthesis under metal toxicity plays an important role in the maintenance of heavy metal homeostasis and/or detoxification by endowing plasma membranes with the capacity to serve as an initial barrier against the inflow of free heavy metal ions into the cells (Suzuki et al., 2002).
Some of these proteins belong to the family of heat shock proteins (HSPs). These proteins are referred to more commonly as ‘stress proteins’. HSPs act as molecular chaperones and help in normal protein folding and assembly, but may also function in the protection and repair of proteins under stressful conditions. It has been demonstrated that the DNA of Cd-stressed cells produces specific mRNA transcripts which regulate the synthesis of stress proteins (Edelman et al., 1988). In several plant species Cd-exposure induces the synthesis of a considerable number of stress proteins with molecular mass ranging from 10-70 kDa. In Oryza sativa, Cd supply induced the synthesis of new proteins with mol. wts. 70, 42, 26, 23, 15 and 11 kDa (Reddy and Prasad, 1995; Prasad, 1997). A 18 kDa Cd inducible protein with 4-SH groups per molecule was isolated by Shah and Dubey (1998a) from rice seedlings. Armeria maritima plants grown on Cu-rich soils showed increased expression of HSP17 (Neumann et al., 1995). Enhanced level of HSP17 was also observed in cell cultures of Silene vulgaris and Lycopersicon esculentum in response to a range of heavy metals (Wollgiehn and Neumann, 1999). Whereas, increased expression of HSP70 was observed in seaweed Entormorpha intestinalis after Cu-exposure (Lewis et al., 2001).
A Cys-rich membrane protein with mol. wt. 16 kDa isolated from Arabidopsis, termed as AtPcrs plays an important role in Cd resistance of Arabidopsis plants and similar homologs have also been reported in Lycopersicon esculentum and Oryza sativa (Song et al., 2004). In Arabidopsis again, increased expression of another metal binding protein gene cdl19 has been reported after exposure of plants to Cd, Hg, Fe and Cu (Suzuki et al., 2002). In Hordeum vulgare seedlings increased synthesis in the amount of apoplastic polypeptides was observed in response to Zn treatment (Brune et al., 1994). Ni also induces synthesis of proteins in many plant species. It has been observed that Ni induces synthesis of proteins at the transcriptional or translational level (Blinda et al., 1997).
Expression of pathogenesis related proteins (PR proteins) has also been reported in certain plants under metal toxicity, but this response solely appears to be metal specific. Synthesis of a 16-kDa polypeptide related to PR-10 family pathogenesis related proteins is induced in Lupinus luteus plants under Cd, Zn and Cu treatment (Przymusińiski et al., 2004). The precise role of PR proteins in heavy metal tolerance is not known, however, it appears that these proteins have ribonuclease like function and are associated with the defense mechanism of plants during pathogen attack (Bantignies et al., 2000). It is believed that the defense reaction of cells might have much more general character, regardless of the kind of stress (Przymusińiski et al., 2004).
Germin and germin-like proteins (GLPs) constitute a ubiquitous family of plant proteins that seem to be involved in many developmental and stress-related processes. Increased expression of one of the wheat germin genes, namely gf-2.8 has been observed in certain plants exposed to the heavy metal ions Cd2+, Cu2+ and Co2+ (Berna and Bernier, 1999).
Chelation of Metals by Organic Acids and Amino Acids
In many plant species an alternative approach for detoxification of metals involves formation of metal-organic acid or metal-amino acid complexes. A wide range of metals, viz., Cd, Pb, Ni, Mn and Al are either partially or in certain cases completely detoxified by this way. Plants are protected from the toxic effects of Ni in their tissues by its complexation with the anions of hydrocarboxylic acids such as citrate, malate and malonate (Reeves 1992; Salt and Krämer, 2000) and the amino acid histidine (Krämer et al., 1996). Generally, the coordination complexes formed by heavy metals with exclusively oxygen-donor centers do not have especially high stability constants (Pollard et al., 2002) whereas histidine acts as a tridentate ligand, at typical cytoplasmic pH values, with its two nitrogen and one oxygen-donor centers and is therefore more efficient and selective chelator of heavy metal Ni than the hydroxycarboxylic acids (Pollard et al., 2002). Recently, complexation of Ni with another high affinity ligand, nicotinamine, has also been reported (Vacchina et al., 2003). In non-polluted soils ([Cd] < 0.3 μM), Cd usually gets complexed with vacuolar citrate and when Cd concentration rises PC plays a crucial role (Wagner, 1993). Cd and Zn resistance of Atriplex halimus L. is due to precipitation of metals in oxalate crystals (Lutts et al., 2004). In Oryza sativa synthesis of oxalate represents a significant response mechanism that enhances tolerance to Pb (Yang et al., 2000).
Organic acid mediated detoxification is an important strategy adopted by plants to reduce Al toxicity. Organic acids chelate with Al and reduce its toxic effects at the cellular level (Ma, 2000). The complexes of Al-citrate (1:1) in hydrangea and Al-oxalate (1:3) in buckwheat have been well characterized (Ma, 2000). Little is known about the way by which Al:organic acid complexes are transported across membranes but it is likely that specific transporters are involved. Chelation of metals with ligands such as histidine, citrate, etc. helps in routing them primarily to the xylem (Krämer et al., 1996; Salt and Krämer, 2000).
Proline
The amino acid proline is the most widespread metabolite that accumulates in many plant species under toxicity due to metals (Shah and Dubey, 1998b). In higher plants accumulating proline appears to be involved in the chelation of excess cytoplasmic metal ions, which show a preference for nitrogen or oxygen coordination (Farago and Mullen, 1979). Cd induced accumulation of proline has been observed in rice and lettuce plants (Shah and Dubey, 1998b; Bertrand and Guary, 2002). Rice plants grown for 15 days in sand cultures containing 160 μM Al3+ showed about 1.8 to 2.7 times increased level of proline compared to the plants growing without Al3+ (Sharma and Dubey, 2005). In several metal tolerant plant species constitutive proline levels are higher than the sensitive plants (Bertrand and Guary, 2002). There is no conclusive evidence for a direct role of proline in the cellular detoxification of heavy metals and there are different opinions regarding the mechanisms by which proline can alleviate metal toxicity. It has been shown that under heavy metal toxicity proline maintains a favourable water balance in plant tissues by serving as osmoprotectant (Paleg et al., 1984). It also acts as protein stabilizer (Sharma and Dubey, 2004), metal chelator (Farago and Mullen, 1979), inhibitor of lipid peroxidation (Mehat and Gaur, 1999), free radical scavenger (Alia et al., 2001), etc. Due to its zwitterionic and high hydrophilic character, proline appears to provide protection to the enzymes and biomolecules (Siripornadulsil et al., 2002). Under in vitro conditions addition of proline in the enzyme assay medium provides considerable protection to the enzymes, the activities of which are inhibited otherwise due to the metals Cd2+, Pb2+, Ni2+ and Al3+ (Sharma and Dubey, 2005). Proline accumulation in metal stressed plants possibly does not appear to be a direct effect of heavy metal stress but as a consequence of the water stress induced by metals (Bertrand and Guary, 2002).
Antioxidative Defense Mechanism
Build up of toxic concentrations of heavy metals within the plant tissues result, at some stage of stress exposure, in an increased formation of reactive oxygen species (ROS) (Shah et al., 2001; Verma and Dubey, 2003). In general these ROS (O2.¯, OH. and H2O2) are products of normal cellular metabolism, their production is under tight control due to cellular antioxidative defense system. Presence of ROS causes oxidative damage to biomolecules such as lipids, proteins, nucleic acids etc. (Shah et al., 2001; Blokhina et al., 2003). Induction in the activities of antioxidative enzymes as well as increase in the level of non-enzymic antioxidants are the strategies adopted by plants to scavenge ROS and to reduce oxidative damage caused due to ROS under heavy metal stress (Shah et al., 2001; Fecht-Christoffers et al., 2003; Verma and Dubey, 2003).
Recent reports suggest that though antioxidative defense system is not directly involved in heavy metal detoxification yet ROS play important role as intermediate signaling molecules to regulate the expression of genes for plant’s defense system (Orzco-Cárdenas et al., 2001; Vranova et al., 2002).
Heavy Metal Sequestration
In Vacuoles
Various metabolites and ions are stored inside the vacuoles. Vacuolar sequestration of a number of heavy metals like Cd, Ni, As, Zn is known which diverts metal ions from metabolically active compartments (cytosol, chloroplasts, mitochondria) and minimizes the harmful effects of metal ions to vital cellular processes. Transporters are present in internal membranes to allow regulation of stored metals in organelles. Active accumulation of most of the metal ions is driven by the electrochemical potential by electrogenic proton influxes via the vacuoler H+-ATPase (Kakinuma et al., 1993). Cd is transported across the tonoplast by a Cd2+/H+ antiport mechanism (Carrier et al., 2003). Both vacuolar (Bidwell et al., 2004) and extravacuolar localization of Ni ion occurs via a pH-gradient dependent manner in yeast (Nishimura et al., 1998) whereas in Ni-hyperaccumulator plant Thalspi goesingense vacuolar metal transport proteins termed as metal-tolerance proteins (TgMTPs) are involved in compartmentalization of Ni in vacuoles in shoots (Persans et al., 2001). Arsenic is also primarily stored in the vacuoles (Lombi et al., 2002). Transport of Zn to the vacuole is mediated by ‘Zn-malate shuttle’; malate being liberated in exchange for oxalate or citrate and is shuttled back to cytoplasm (Ernst et al., 1992) but the storage form of Zn in the vacuoles is not much studied. A gene (ZAT) closely related to animal ZnT (Zn transporter) gene has been isolated from Arabidopsis (Van der Zaal et al., 1999). Overexpression of ZAT gene in transgenic plants led to enhanced accumulation of Zn in the roots under high Zn treatment (Van der Zaal et al., 1999).
In Trichomes and Hydropotes
Apart from vacuolar sequestration, plants possess additional morphological features that are also involved in heavy metal sequestration and detoxification. Several reports have confirmed the involvement of glandular trichomes and epidermal structures (hydropotes) in the chelation, sequestration and detoxification of the metals.
Trichomes are epidermal hairs present at the surface of plant leaves and have diversified roles in exudation of various molecules, protection against wind and sunlight, storage of metals, etc. Retardation in growth and about two-fold increase in the number of trichomes was observed in Cd-exposed tobacco seedlings (Choi et al., 2001). A significant proportion of Ni has been found in trichomes of Alyssum lesbiacum plants (Krämer et al., 1997). At the bases of Arabidopsis halleri trichomes, elevated concentrations of Zn have been found. In these trichomes Zn remains tetrahedrally coordinated and complexed to carboxyl and/or hydroxyl functional groups of certain secondary organic species (Sarret et al., 2002). Further studies using variable-pressure scanning electron microscopy and energy dispersive X-ray analysis indicated that head cells of tobacco trichomes actively exclude toxic Cd by forming and excreting Cd/Ca-containing crystals (Choi et al., 2001). Specific overexpression of a gene coding for a metallothionein (MT2) has been reported in trichomes (Garcia-Hernandez et al., 1998), which suggests that trichomes constitute important sites for accumulation as well as detoxification of toxic metal ions.
In the semiaquatic and aquatic plants of the families Menyanthaceae and Nymphaceae; hydropotes located on the abaxial epidermis of the leaf laminae accumulate Cd (Lavid et al., 2001a). It is suggested that usual polymerization of polyphenols by peroxidases in hydropotes gets enhanced after uptake of heavy metals and thereby detoxification of metals occurs by their binding with polyphenols in these glands (Lavid et al., 2001b; 2001c).
Heavy Metal Detoxification and Signal Transduction Pathway
Though no unified signal transduction pathway exists in higher plants for detoxification of heavy metals, signal transduction cascades have been hypothesized which trigger the detoxification process. Likely involvement of the second messengers- calcium or fatty acid derivative jasmonic acid (JA) has been suggested (Maksymiec and Baszynski, 1999). In addition, activation of distinct mitogen activated protein kinase pathways has been observed in response to heavy metal stress (Jonak et al., 2004). It was observed by Xiang and Oliver (1998) that Arabidopsis plants treated with Cd or Cu responded by increasing transcription of the genes of glutathione (precursor of PC) synthesis and that jasmonic acid also activated same set of genes. However, it is not clear whether JA is involved in heavy metal induced expression of genes in Arabidopsis. Similarly, Cu2+ ions induced a marked increase in the content of jasmonic acid in rice (Rakwal et al., 1996).
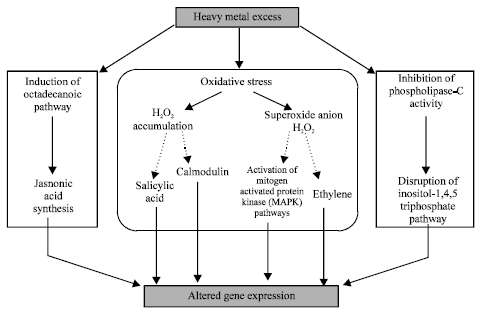 | |
| Fig. 3: | Potential routes by which heavy metals can influence gene expression. |
Both excess metal ions and JA or its derivative methyl jasmonate exert similar effects in plants (Maksymiec and Krupa, 2002). These observations suggest that possible pathways for the action of heavy metals and jasmonic acid appear to be similar. Maksymiec et al. (2002), suggested that heavy metals might induce octadecanoic pathways leading to synthesis of JA which would consequently act as elicitor and would generate metal toxicity symptoms. A generalized view of the potential routes through which heavy metals influence gene expression, has been presented in Fig. 3.
Most of the heavy metals enhance production of H2O2 and it is quite possible that accumulating H2O2 may somehow switch-on the signal transduction pathway meant for heavy metal detoxification. Role of H2O2 as a signaling molecule is increasingly being realized that activates expression of several genes (Orozco-Cárdenas et al., 2001).
Gene expression patterns change when plants encounter excessive amounts of heavy metals. Some of the heavy metal responsive genes code for signal transduction components, such as the Arabidopsis mitogen-activated protein kinase kinase kinase (MAPKKK), MEKK1, transcription factors, stress induced proteins etc. (Xiang and Oliver, 1998; Suzuki et al., 2001; Louie et al., 2003). In several plant species, mitogen activated protein kinase (MAPK) pathways are activated in response to heavy metal stress (Xiang and Oliver, 1998; Suzuki et al., 2001; Louie et al., 2003). Jonak et al. (2004) observed that exposure of alfalfa (Medicago sativa) seedlings to excess Cu or Cd ions activated four distinct MAPKs: SIMK, MMK2, MMK3 and SAMK. There exists possibility that MAPK activation by excess heavy metals is mediated by ROS, since heavy metals are known to disturb redox control of the cell. Several studies in different plant species support this view of MAPK activation by ROS (Nakagami et al., 2004; Rentel et al., 2004). A number of signal transduction components are involved in the detection of H2O2 and the activation of defense mechanisms in plants. These include: A two-component histidine kinase, a receptor like protein kinase, the MAPKK, AtANP1 (also the NtNPK1) and the MAPKs, AtMPK316 and Ntp46MAPK (Desikan et al., 2001; Vranova et al., 2002). In addition, calmodulin has been implicated in H2O2 signaling (Desikan et al., 2001). So far, the evidence for an involvement of MAPK pathway in heavy metal stress is scarce in plants and significance of MAPK signaling in response to heavy metal excess has yet to be fully understood.
Ethylene is a natural plant growth regulator involved in the control of a wide range of developmental responses (Kunkel and Brooks, 2002; Milone et al., 2003). Heavy metals induce ethylene biosynthesis (Milone et al., 2003) and it is well known that production of ethylene influences the plant defense response (Kunkel and Brooks, 2002).
Salicylic acid (SA) has been identified as an important signaling element involved in establishing the response of plants after abiotic stress (Alvarez, 2000). The significance of SA in signaling under heavy metal stress is supported by the fact that heavy metals induce an increase in SA content (Metwally et al., 2003). SA activates defense related genes either by H2O2 mediated signal transduction pathway or by directly affecting mechanisms of metal detoxification (Chen et al., 1993; Metwally et al., 2003). SA inhibits two major H2O2 scavenging enzymes catalase and ascorbate peroxidase which cause cellular H2O2 concentration to rise and this accumulating H2O2 subsequently acts as second messenger and activates defense related genes (Chen et al., 1993). Inositol-1,4,5,-triphosphate (IP3) also participates in transducing signals under metal stress conditions. Inhibition of phospholipase-C activity and subsequent disruption of the IP3 signal transduction pathway has been reported under aluminium excess (Jones and Kochian, 1995).
Conclusions
The increasing release of heavy metals Cd, Pb, As, Hg, Ni into the environment has caused concern for human and plant health. These heavy metals serve as major pollutants of the environment and drastically affect plant growth and its metabolism ultimately leading to severe losses in yield. Metal ions reach within the plant tissues through a number of membrane transporters localized in the root. A number of metal transporters and their gene families have now been identified but still their metal ion specificities need to be established and there are still certain heavy metals for whom no specific membrane transporters are known in higher plants. Once inside the cell these metal ions interact with biomolecules and cellular organelles, various metabolic processes are adversely affected. Plants have developed defense mechanisms to combat heavy metal induced injury. Extracellularly plants immobilize metals near root region either with the help of root exudates or with mycorrhizal association. Among the intracellular detoxification mechanisms adopted by plants the most promising one involves sequestration of metal ions by the formation of complexes between metal cation and the peptides phytochelatins synthesized within the plant tissues. Formation of metal complexes with organic acids or amino acids appears to be an alternate approach to detoxify metals within the plant tissues. Synthesis of stress specific proteins, amino acid proline and increased production of reactive oxygen species are observed under toxicity with metals Cd, Pb, Cu, etc. More than one mechanism may be involved in mitigating the toxicity of particular metal. Much work is needed to fully unveil the mechanisms associated with toxicity of heavy metals in plants as well as the mechanisms associated with plant metal-tolerance. Tolerance of plants to certain metals is associated with increased synthesis of phytochelatins, proline and elevated levels of antioxidant enzymes. Overexpression of animal metallothionein gene in higher plants and cyanobacteria as well as overproduction of phytochelatins in transgenic plants confer heavy metal tolerance in the transformants. On encountering with excessive amounts of heavy metals, gene expression patterns in plants change. In Arabidopsis it has been shown that heavy metal responsive genes code for signal transduction components however, in higher plants pathways through which metal stress stimuli are transduced await further investigation.
The concept of phytoremediation of contaminated soils has been increasingly supported by research in recent years. The understanding of heavy metal detoxification processes afforded by genetic and molecular investigations, would allow us to explore the mechanisms by which some species are capable of higher-accumulation of heavy metals and how they may be best used for phytoremediation. The understanding of cellular signaling induced by heavy metal stress will be a vital tool in further improving this technology. With these few examples it appears that once the salient components associated with metal tolerance are well-established, future trials of genetic engineering may provide successful transformants with improved tolerance to heavy metals.
ACKNOWLEDGMENTS
One of the authors (SM) is grateful to University Grants Commission, New Delhi for the award of a Senior Research Fellowship.
REFERENCES
- Abedin, M.J., J. Feldmann and A.A. Meharg, 2002. Uptake kinetics of arsenic species in rice plants. Plant Physiol., 128: 1120-1128.
CrossRef - Alverez, M.E., 2000. Salicylic acid in the machinery of hypersensitive cell death and disease resistance. Plant Mol. Biol., 44: 429-442.
PubMedDirect Link - Arazi, T., B. Kaplan, R. Sunkar and H. Fromm, 2000. Cyclic-nucleotide- and Ca2+/calmodulin-regulated channels in plants: Targets for manipulating heavy-metal tolerance and possible physiological roles. Biochem. Soc. Trans., 28: 471-475.
PubMedDirect Link - Bantignies, B., J. Seguin, I.M. Muzac, F. Dedaldechamp, P. Gulick and R. Ibrahim, 2000. Direct evidence for ribonucleolytic activity of a PR-10-like protein from white lupin roots. Plant Mol. Biol., 42: 871-881.
PubMedDirect Link - Berna, A. and F. Bernier, 1999. Regulation by biotic and abiotic stress of a wheat Germin gene encoding oxalate oxidase, a H2O2-producing enzyme. Plant Mol. Biol., 39: 539-549.
Direct Link - Blaudez, D., B. Botton and M. Chalot, 2000. Cadmium uptake and subcellular compartmentation in the ectomycorrhizal fungus Paxillus involutus. Microbiology, 146: 1109-1117.
CrossRefDirect Link - Blokhina, O., E. Virolainen and K.V. Fagerstedt, 2003. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot., 91: 179-194.
CrossRefPubMedDirect Link - Choir, Y.E., E. Harada, M. Wada, I.M. Tsubo, Y. Morita, T. Kusano and H. Sano, 2001. Detoxification of cadmium in tobacco plants: Formation and active secretion of crystals containing cadmium and calcium through trichomes. Plant, 213: 45-50.
Direct Link - Garcia-Hernandez, M., A. Murphy and L. Taiz, 1998. Metallothioneins 1 and 2 have distinct but overlapping expression patterns in Arabidopsis. Plant Physiol., 118: 387-397.
Direct Link - Giles, N.M., G.I. Giles and C. Jacob, 2003. Multiple roles of cysteine residue in biocatalysis. Biochem. Biophys. Res. Commun., 300: 1-4.
Direct Link - Jentschke, G. and D.L. Godbold, 2000. Metal toxicity and ectomycorrhizas. Physiologia Plantarum, 109: 107-116.
CrossRefDirect Link - Kramer, U., J.D. Cotter-Howells, J.M. Charnock, A.J.M. Baker and J.A.C. Smith, 1996. Free histidine as a metal chelator in plants that accumulate nickel. Nature, 379: 635-638.
Direct Link - Kunkel, B.N. and D.M. Brooks, 2002. Cross talk between signaling pathways in pathogen defense. Curr. Opin. Plant Biol., 5: 325-331.
CrossRefPubMedDirect Link - Kupper, H., A. Mijovilovich, W. Meyer-Klaucke and P.M.H. Kroneck, 2004. Tissue-and age-dependent differences in the complexation of cadmium and zinc in the cadmium/zinc hyperaccumulator Thlaspi caerulescens (Ganges ecotype) revealed by X-ray absorption spectroscopy1[w]. Plant Physiol., 134: 748-757.
- Lee, H.S. and B. Volesky, 1997. Interaction of light metals and protons with seaweed biosorbent. Water Res., 31: 3082-3088.
Direct Link - Leita, L., M. Dc Nobili, S. Cesco and C. Mondini, 1996. Analysis of intercellular cadmium forms in roots and leaves of bush bean. J. Plant Nutr., 19: 527-533.
Direct Link - Lewis, S., M.E. Donkin and M.H. Depledge, 2001. Hsp70 expression in Enteromorpha intestinalis (Chlorophyta) exposed to environmental stressors. Aquatic Toxicol., 51: 277-291.
CrossRef - Li, Z.S., Y.P. Lu, D.J. Thiele and P.A. Rea, 1997. A new pathway for vacuolar cadmium sequestration in Saccharomyces cerevisiae: YCF1- mediated transport of BIS (glutathionato) cadmium. Proc. Natl. Acad. Sci. USA., 94: 42-47.
Direct Link - Lombi, E., F.J. Zhao, M. Fuhrman, L. Ma and S.P. McGrath, 2002. Arsenic distribution and speciation in the fronds of the hyperaccumulator Pteris vittata. New Phytol., 156: 195-203.
Direct Link - Louie, M., N. Kondor and J.G. de Witt, 2003. Gene expression in cadmium-tolerant Datura innoxia: Detection and characterization of cDNAs induced in response to Cd2+. Plant Mol. Biol., 52: 81-89.
PubMedDirect Link - Lutts, S., I. Lefevre, C. Delperee, S. Kivits, C. Dechamps, A. Robledo and E. Correal, 2004. Heavy metal accumulation by the halophyte species Mediterranean saltbush. J. Environ. Qual., 33: 1271-1279.
CrossRefDirect Link - Ma, J.F., 2000. Role of organic acids in detoxification of aluminium in higher plants. Plant Cell Physiol., 41: 383-390.
PubMedDirect Link - Ma, J.F., D. Ueno, F.J. Zhao and S.P. McGrath, 2005. Subcellular localization of Cd and Zn in the leaves of a Cd-hyperaccumulating ecotype of Thlaspi caerulescens. Planta, 220: 731-736.
PubMedDirect Link - Maiti, I.B., G.J. Wagner, R. Yeargen and A.G. Hunt, 1989. Inheritance and expression of the mouse metallothionein gene in tobacco. Impact on Cd tolerance and tissue Cd distribution in seedlings. Plant Physiol., 91: 1020-1024.
Direct Link - Maksymiec, W. and T. Baszyski, 1999. The role of Ca2+ ions in modulating changes induced in bean plants by an excess of Cu2+ ions. Chlorophyll fluorescence measurements. Physiol. Plant., 105: 562-568.
Direct Link - Maksymiec, W. and Z. Krupa, 2002. Jasmonic acid and heavy metals in Arabidopsis plants: A similar physiological response to both stressors? J. Plant Physiol., 159: 509-515.
CrossRef - Meharg, A.A. and J. Hartley-Whitaker, 2002. Arsenic uptake and metabolism in arsenic resistant and nonresistant plant species. New Phytol., 154: 29-43.
CrossRefDirect Link - Meharg, A.A. and L. Jardine, 2003. Arsenite transport into paddy rice (Oryza sativa) roots. New Phytol., 157: 39-44.
Direct Link - Mehta, S.K. and J.P. Gaur, 1999. Heavy-metal-induced proline accumulation and its role in ameliorating metal toxicity in Chlorella vulgaris. New Phytol., 143: 253-259.
Direct Link - Metwally, A., I. Finkemeier, M. Georgi and K.J. Dietz, 2003. Salicylic acid alleviates the cadmium toxicity in barley seedlings. Plant Physiol., 132: 272-281.
CrossRefDirect Link - Milone, M.T., C. Sgherri, H. Clijsters and F. Navari-Izzo, 2003. Antioxidative responses of wheat treated with realistic concentration of cadmium. Environ. Exp. Bot., 50: 265-276.
Direct Link - Murphy, A., J.M. Zhou, P.B. Goldsbrough and L. Taiz, 1997. Purification and immunological identification of metallothioneins 1 and 2 from A. thaliana. Plant Physiol., 113: 1293-1301.
PubMedDirect Link - Nakagami, H., S. Kiegeri and H. Hirt, 2004. OMTK1, a novel MAPKKK, channels oxidative stress signaling through direct MAPK interaction. J. Biol. Chem., 279: 26959-26966.
Direct Link - Neumann, D., U.Z. Nieden, O. Lichtenberger and I. Leopold, 1995. How does Armeria maritima tolerate high heavy metal concentrations? J. Plant Physiol., 146: 704-717.
Direct Link - Nishimura, K., K. Igarashi and Y. Kakinuma, 1998. Proton gradient driven nickel uptake by vacuolar membrane vesicles of Saccharomyces cerevisiae. J. Bacteriol., 180: 1962-1964.
PubMedDirect Link - Orozco-Cardenas, M.L., J. Narvaez-Vasquez and C.A. Ryan, 2001. Hydrogen peroxide acts as a second messenger for the induction of defense genes in tomato plants in response to wounding, systemin and methyl jasmonate. Plant Cell, 13: 179-191.
PubMed - Paleg, L.G., G.R. Stewart and J.W. Bradbeer, 1984. Proline and glycine-betaine influence on protein solvation. Plant Physiol., 75: 974-978.
PubMedDirect Link - Panda, K.K., L. Maheswar and B.P. Brahma, 1992. Monitoring and assessment of mercury pollution in the vicinity of a chloralkali plant. II plant availability, tissue concentration and genotoxicity of mercury from agricultural soil contaminated with solid waste assessed in barley (Hordeum vulgare L.). Environ. Pollut., 76: 33-42.
PubMedDirect Link - Pence, N.S., P.B. Larsen, S.D. Ebbs, D.L.D. Letharn and M.M. Lasat et al., 2000. The molecular physiology of heavy metal transport in the Zn/Cd hyperaccumulator Thlaspi caerulescens. Proc. Natl. Acad. Sci. USA., 97: 4956-4960.
CrossRefDirect Link - Persans, A.W., K. Nieman and D.E. Salt, 2001. Functional activity and role of cation-efflux family members in Ni hyperaccumulation in Thlaspi goesingense. Proc. Natl. Acad. Sci. USA., 98: 9995-10000.
Direct Link - Pickering, I.J., R.C. Prince, M.J. George, R.D. Smith, G.N. George and D.E. Salt, 2000. Reduction and coordination of arsenic in Indian mustard. Plant Physiol., 122: 1171-1178.
Direct Link - Piechalak, A., B. Tomaszewska, D. Baralkiewicz and A. Malecka, 2002. Accumulation and detoxification of lead ions in legumes. Phytochemistry, 60: 153-162.
CrossRefDirect Link - Pollard, A.J., K.D. Powell, F.A. Harper and J.A.C. Smith, 2002. The genetic basis of metal hyperaccumulation in plants. Curr. Rev. Plant Sci., 21: 539-566.
CrossRef - Przymusiski, R., R. Ruciska and E.A. Gwoidi, 2004. Increased accumulation of pathogenesis-related proteins in response of lupine roots to various abiotic stresses. Environ. Exp. Bot., 52: 53-61.
Direct Link - Raab, A., J. Feldmann and A.A. Meharg, 2004. The nature of arsenic-phytochelatin complexes in Holcus lanatus and Pteris cretica. Plant Physiol., 134: 1113-1122.
Direct Link - Rauser, W.E., 1995. Phytochelatins and related peptides. Structure, biosynthesis and function. Plant Physiol., 109: 1141-1149.
PubMedDirect Link - Rea, P.A., Z.S. Li, Y.P. Lu and Y.M. Drozdowicz, 1998. From vacuolar GS-X pumps to multispecific ABC transporters. Annu. Rev. Plant Physiol. Plant Mol. Biol., 49: 727-760.
CrossRefDirect Link - Rea, P.A., O.K. Vatamaniuk and D.J. Rigden, 2004. Weeds, worms and more. Papain's long-lost cousin, phytochelatin synthase. Plant Physiol., 136: 2463-2474.
Direct Link - Reddy, G. and M.N.V. Prasad, 1995. Cadmium-induced protein phosphorylation changes in rice (Oryza sativa L.) seedlings. J. Plant Physiol., 145: 67-70.
Direct Link - Rentel, M.C., D. Lecourieux, F. Ouaked, S.L. Usher and L. Petersen et al., 2004. OXI1 kinase is necessary for oxidative burst-mediated signaling in Arabidopsis. Nature, 427: 858-861.
CrossRefDirect Link - Robinson, N.J., I.M. Evans, J. Mulcrone, J. Bryden and A.M. Tommey, 1992. Genes with similarity to metallothionein genes and copper, zinc ligands in Pisum sativum L. Plant Soil, 146: 291-298.
CrossRef - Ruotolo, R., A. Peracchi, A. Bolchi, G. Infusini, A. Amoresano and S. Ottonello, 2004. Domain organization of phytochelatin synthase: Functional properties of truncated enzyme species identified by limited proteolysis. J. Biol. Chem., 279: 14686-14693.
Direct Link - Ryan, P.R. and E. Delhaize, 2001. Function and mechanism of organic anion exudation from plant roots. Annu. Rev. Plant Physiol. Plant Mol. Biol., 52: 527-560.
CrossRefDirect Link - Salt, D.E. and W.E. Rauser, 1995. Mg ATP-dependent transport of phytochelatins across the tonoplast of oat roots. Plant Physiol., 107: 1293-1301.
PubMed - Salt, D.E., N. Kato, U. Kramer, R.D. Smith and I. Raskin, 2000. The Role of Root Exudates in Nickel Hyperaccumulation and Tolerance in Accumulator and Nonaccumulator Species of Thlaspi. In: Phytoremediation of Contaminated Soil and Water, Terry, E. and G. Banuelos (Eds.). CRC Press, UK., pp: 189-200.
- Sarret, G., P. Saumitou-Laprade, V. Bert, O. Proux and J.L. Hazemann et al., 2002. Forms of zinc accumulated in the hyperaccumulator Arabidopsis halleri. Plant Physiol., 130: 1815-1826.
Direct Link - Satofuka, H., T. Fukui, M. Takagi, H. Atomi and T. Imanaka, 2001. Metal-binding properties of phytochelatin-related peptides. J. Inorg. Biochem., 86: 595-602.
CrossRef - Schutzendubel, A. and A. Polle, 2002. Plant responses to abiotic stresses: Heavy metal-induced oxidative stress and protection by mycorrhization. J. Exp. Bot., 53: 1351-1365.
CrossRefPubMedDirect Link - Shah, K. and R.S. Dubey, 1998. A 18 kDa Cd inducible protein complex: Its isolation and characterization from rice (Oryza sativa L.) seedlings. J. Plant Physiol., 152: 448-454.
Direct Link - Shah, K. and R.S. Dubey, 1997. Effect of cadmium on proline accumulation and ribonuclease activity in rice seedlings: Role of proline as a possible enzyme protectant. Biol. Plant., 40: 121-130.
CrossRef - Shah, K., R.G. Kumar, S. Verma and R.S. Dubey, 2001. Effect of cadmium on lipid peroxidation, superoxide anion generation and activities of antioxidant enzymes in growing rice seedlings. Plant Sci., 161: 1135-1144.
CrossRefDirect Link - Sharma, P. and R.S. Dubey, 2004. Ascorbate peroxidase from rice seedlings: Properties of enzyme isoforms, effects of stresses and protective roles of osmolytes. Plant Sci., 167: 541-550.
CrossRef - Sharma, P. and R.S. Dubey, 2005. Modulation of nitrate reductase activity in rice seedlings under aluminium toxicity and water stress: Role of osmolytes as enzyme protectant. J. Plant Physiol., 162: 854-864.
Direct Link - Siripornadulsil, S., S. Traina, D.P.S. Verma and R.T. Sayre, 2002. Molecular mechanisms of proline-mediated tolerance to toxic heavy metals in transgenic microalgae. Plant Cell., 14: 2837-2847.
Direct Link - Song, W.Y., E. Martinoia, J. Lee, D.Y. Kim and E. Vogt et al., 2004. A novel family of cys-rich membrane proteins mediates cadmium resistance in Arabidopsis. Plant Physiol, 135: 1027-1039.
Direct Link - Steffens, J.C., 1990. The heavy metal-binding peptides of plants. Ann. Rev. Plant Physiol. Plant Mol. Biol., 41: 553-575.
CrossRef - Suzuki, N., N. Koizumi and H. Sano, 2001. Screening of cadmium-responsive genes in Arabidopsis thaliana. Plant Cell and Environ., 24: 1177-1188.
CrossRef - Suzuki, N., Y. Yamaguchi, N. Koizumi and H. Sano, 2002. Functional characterization of a heavy metal binding protein Cdl19 from Arabidopsis. Plant J., 32: 165-173.
PubMedDirect Link - Tsuji, N., S. Nishikori, O. Iwabe, K. Shiraki and H. Miyasaka et al., 2004. Characterization of phytochelatin synthase-like protein encoded by alr0975 from a prokaryote, Nostoc sp. PCC 7120. Biochem. Biophy. Res. Commun., 315: 751-755.
CrossRef - Thomine, S., R. Wang, J.M. Ward, N.M. Crawford and J.I. Schroeder, 2000. Cadmium and iron transport by members of a plant metal transporter family in Arabidopsis with homology to Nramp genes. Proc. Natl. Acad. Sci. USA., 97: 4991-4996.
Direct Link - Tung, G. and P.J. Temple, 1996. Uptake and localization of lead in corn (Zea mays L.) seedlings, a study by histochemical and electron microscopy. Sci. Total Environ., 188: 71-85.
CrossRefDirect Link - Vacchina, V., S. Mari, P. Czernic, L. Marques and K. Pianelli et al., 2003. Speciation of nickel in a hyperaccumulating plant by high-performance liquid chromatography-inductively coupled plasma mass spectroscopy and electrospray MS/MS assisted by cloning using yeast complementation. Anal. Chem., 75: 2740-2745.
CrossRef - Van der Zaal, B.J., L.W. Neuteboom, J.E. Pinas, A.N. Chardonnens and H. Schat et al., 1999. Overexpression of a novel Arabidopsis gene related to putative zinc-transporter genes from animals can lead to enhanced zinc resistance and accumulation. Plant Physiol., 119: 1047-1055.
PubMedDirect Link - Verma, S. and R.S. Dubey, 2003. Lead toxicity induces lipid peroxidation and alters the activities of antioxidant enzymes in growing rice plants. Plant Sci., 164: 645-655.
Direct Link - Vranova, E., D. Inze and F. Van Breusegem, 2002. Signal transduction during oxidative stress. J. Exp. Bot., 53: 1227-1236.
CrossRefDirect Link - Wagner, G.J., 1993. Accumulation of cadmium in crop plants and its consequences to human health. Adv. Agron., 51: 173-212.
Direct Link - Williams, L.E., J.K. Pittman and J.L. Hall, 2000. Emerging mechanisms for heavy metal transport in plants. Biochim. Biophys. Acta, 1465: 104-126.
PubMedDirect Link - Wojciechowska-Mazurek, M., T. Zawadzka, K. Karlowski, K. Cwiek-Ludwicka and E. Brulinska-Ostrowska, 1995. Content of lead, cadmium, mercury, zinc and copper in fruit from various regions of Poland. Rocz. Panst. Zakl. Hig., 46: 223-238.
PubMedDirect Link - Wollgiehn, R. and D. Neumann, 1999. Metal stress response and tolerance of cultured cells from Silene vulgaris and Lycopersicon peruvianum: Role of heat stress proteins. J. Plant Physiol., 154: 547-553.
Direct Link - Wong, H.L., T. Sakamoto, T. Kawasaki, K. Umemura and K. Shimamoto, 2004. Down-regulation of metallothionein, a reactive oxygen scavenger, by the small GTPase OsRac1 in rice. Plant Physiol., 135: 1447-1456.
Direct Link - Xiang, C. and D.J. Oliver, 1998. Glutathione metabolic genes coordinately respond to heavy metals and jasmonic acid in Arabidopsis. Plant Cell, 10: 1539-1550.
Direct Link - Yang, Y.Y., J.Y. Jung, W.Y. Song, H.S. Suh and Y. Lee, 2000. Identification of rice varieties with high tolerance or sensitivity to lead and characterization of the mechanism of tolerance. Plant Physiol., 124: 1019-1026.
PubMedDirect Link - Zenk, M.H., 1996. Heavy metal detoxification in higher plants-a review. Gene, 179: 21-30.
CrossRefDirect Link - Zhou, J. and P.B. Goldsbrough, 1994. Functional homologs of fungal metallothionein genes from Arabidopsis. Plant Cell, 6: 875-884.
PubMedDirect Link - Marrs, K.A., 1996. The functions and regulation of glutathione S-transferases in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol., 47: 127-158.
CrossRefDirect Link