
H. S. Jayasinghearachchi
Institute of Fundamental Studies, Hantan Road, Kandy, Sri Lanka
G. Seneviratne
Institute of Fundamental Studies, Hantan Road, Kandy, Sri Lanka
H.M. S.P.M. Weerasinghe
Institute of Fundamental Studies, Hantan Road, Kandy, Sri Lanka
International Journal of Agricultural Research
Year: 2006 | Volume: 1 | Issue: 1 | Page No.: 1-7







ABSTRACT
Effects of tannic acid on rhizobial performance and legume-rhizobial N2 fixing symbiosis were examined using soybean as the test plant. Two strains, Bradyrhizobium elkanii SEMIA 5019 and B. japonicum TAL 102 were grown in continuous culture media with different concentrations of tannic acid for 28 days to produce tannic acid-affected rhizobia. Soybean inoculated by the affected rhizobia in gravel pots was evaluated for plant growth, nodulation and N accumulation after 40 days. Two other sets of soybean plants inoculated by unaffected, normal rhizobia were treated with tannic acid separately at planting or two weeks after planting. This was to examine the effect of soil solution tannic acid on the symbiosis. Plant growth, nodulation and N accumulation of soybean were reduced when the rhizobial strains were affected mainly by ≥0.2 mM tannic acid. This was conspicuous in B. japonicum TAL 102. Tannic acid was responsible for biochemical changes of the rhizobial proteins, thus resulting in their alterations, especially in B. japonicum TAL 102. When grown in culture media with 0.4 mM tannic acid, an increased shoot N accumulation was observed in inoculated soybean. It is concluded from this study that soil tannins have both favorable as well as unfavorable effects on the legume-rhizobial N2 fixing symbiosis. They reduce rhizobial performance by direct interactions whereas they enhance N2 fixing activity possibly by removing soil mineral N in the proximity of legume root system at planting.
PDF Abstract XML References
How to cite this article
H. S. Jayasinghearachchi, G. Seneviratne and H.M. S.P.M. Weerasinghe, 2006. Tannin Interactions with Legume-rhizobial N2 Fixing Symbiosis. International Journal of Agricultural Research, 1: 1-7.
DOI: 10.3923/ijar.2006.1.7
URL: https://scialert.net/abstract/?doi=ijar.2006.1.7
DOI: 10.3923/ijar.2006.1.7
URL: https://scialert.net/abstract/?doi=ijar.2006.1.7
INTRODUCTION
Tannins are a naturally occurring chemically diverse group of water-soluble plant polyphenolics, which combine with proteins and other polymers to form stable, soluble or insoluble complexes (Hagerman et al., 1992; Konishi and Tanaka, 1999). They are biologically active chemicals with molecular masses between 500 Da and 3000 Da and are widely distributed in the soil. The relative abundance of these compounds varies widely among plant species and the tissue type (Kogel-Knabner, 2002). Tannins also have diverse effects on biological systems, as they are good chelators of ions (Mila and Scalbert, 1994), biological anti-oxidant and anti-microbial agents (Scalbert, 1991).
In natural and managed ecosystems, large amounts of tannins are released to the soil during litter turnover. They, being anti-microbial compounds affect the growth and activity of rhizobia in the soil (Mila and Scalbert, 1994). In addition, seed and root tannins of nodulating legumes have been observed to reduce rhizobial growth and nodulation (Wolff et al., 1993; Balasubramani et al., 1992). Thus, like many other environmental stress factors such as acidity, salinity, drought etc. that reduce rhizobial performance (Giller, 2001), tannins also play an important role in altering rhizobial effectiveness. However, this has not been adequately studied in terms of especially legume-rhizobial N2 fixing symbiosis. Present study investigates the effects of rhizobia affected by tannic acid, on legume-rhizobial N2 fixing symbiosis using soybean as the test plant. Bacteroids of the affected rhizobia in soybean root nodules are protein-fingerprinted to examine their biochemical changes. Further, the effect of tannic acid on the legume-rhizobial interaction in the symbiosis is also evaluated.
MATERIALS AND METHODS
Culturing Rhizobia
B. elkanii SEMIA 5019 and B. japonicum TAL 102, developed for soybean were used. Cultures were maintained in Yeast Manitol Broth (YMB) (Somasegaran and Hoben, 1994). They were incubated on a rotary shaker at 28°C for 6 days. Purity of the cultures was tested using yeast manitol agar with congo-red (CRYMA) plates and gram staining.
Evaluation of Growth, Nodulation and N Accumulation of Soybean When Inoculated with Tannic Acid-affected Rhizobia
The two bradyrhizobial strains were grown in tannic acid treated continuous cultures for 28 days. Commercial tannic acid (Vel, Geldenasksebaan 464, H.R/R.C, Leuven 6172, Belgium) was used for this. First, filter-sterilized tannic acid was added to an autoclaved YMB in order to give final concentrations of 0.05, 0.1, 0.2 and 0.4 mM. This range of concentrations was within the range of tannins found in soil (Rice and Pancholy, 1973). Three replicates were included for each concentration. Control cultures of each strain were maintained in the media without tannic acid. All the cultures were incubated at 28°C on a rotary shaker.
Soybean seeds were surface sterilized, pre-germinated at 20°C and sown (3 seedlings per pot) in regiform pots filled with a heat sterilized gravel medium and McKnight solution, a sterilized N-free plant nutrient medium (McKnight, 1949). Plants were inoculated separately with 3 mL of the tannic acid-affected cultures above and the control cultures, containing ~109 cells mL-1. Six replicates were maintained and pots were arranged in a Completely Randomized Design (CRD) in a greenhouse. Plants were grown with a daily minimum-maximum temperature range of 25-30°C. They were watered daily and the McKnight solution was supplied once a week. Plants were harvested after 40 days of growth (at flowering). Shoot, root and nodule dry weights were recorded. Total nitrogen concentrations of dried and ground shoot and root samples were estimated using Kjeldahl method (Bremner and Tabatabai, 1972).
Extraction of Proteins from Bradyrhizobial Bacteroids Isolated from Soybean Nodules
Only two concentrations (0.2 and 0.4 mM) of tannic acid were considered for the extraction of tannic acid-affected rhizobial proteins. Bacteroids of SEMIA 5019 and TAL 102 were isolated from fresh soybean nodules using a technique described by Day et al. (1989), with some modifications. Nodules were crushed in 20 mM potassium phosphate buffer containing 0.1 mM sodium ascorbate and soluble polyvinylpyrrolidone K30 (pH 7.4) (Eady, 1980). The macerate was squeezed through four layers of cheesecloth to remove large debris. The filtrate was centrifuged at 78 x g for 10 min to remove plant cell debris. Pellet was discarded and supernatant was collected. Supernatant was re-centrifuged at 15,339 x g for 15 min. Supernatant was discarded and pellet was re-suspended in sodium phosphate buffer and re-centrifuged. Supernatant was discarded and the pellet was re-suspended in sample treatment buffer containing DTT and 20% SDS. Bacteroid suspension was sonicated for 2 min at 65% power and heated in a water bath at 95°C for 10 min precisely. After heating, placing them in an ice bath immediately cooled down the tubes and then they were centrifuged at 78 x g for 5 min and the supernatant (protein extract) was collected.
Gel Electrophoresis of Bacteroid Proteins
One-dimensional SDS polyacrylamide gel electrophoresis (1D SDS PAGE) was carried out for the proteins of the bacteroids (Botha and Jooste, 1992; Dewettink et al., 1992). Gels were stained with coomassie brilliant blue to visualize the proteins (Sambrook et al., 1989). They were analyzed using cluster analysis of Gel Compar (1993) computer software. The unweighed pair group method with arithmetic means (UPGMA; Sneath and Sokal, 1973) was applied for the analysis.
Evaluation of Growth, Nodulation and N Accumulation When Inoculated Soybean Plants Were Affected by Tannic Acid
Tannic acid (0, 0.05, 0.2 and 0.4 mM) was applied to two separate sets of gravel pot plant cultures containing McKnight solution; application to the first set at planting and the other at two weeks after planting. Seedlings were inoculated at planting with 3 mL broth of unaffected SEMIA 5019, containing ~109 cells mL-1. Plants were maintained and treated in a similar manner, as described above.
Data analysis
All data were analyzed by GLM procedure of SAS software (SAS, 1987). Means were separated using Tukey’s HSD test.
RESULTS
Effect of Tannic Acid-affected Rhizobia on N2 Fixing Symbiosis of Soybean
Nodule dry weight of soybean decreased significantly with inoculation of SEMIA 5019 affected by 0.2 or 0.4 mM tannic acid (p≤0.001; Table 1). With TAL 102, a significant reduction in nodule dry weight was observed when it was affected only by 0.4 mM tannic acid (p≤0.001). Shoot and root dry weights of soybean decreased significantly compared to the control, when inoculated with SEMIA 5019 or TAL 102 that was affected by 0.4 mM tannic acid (p≤0.001). Root dry weight decreased significantly when inoculated with SEMIA 5019, affected by all three concentrations of tannic acid (p≤0.001). However, shoot dry weight did not show significant differences when inoculated with 0.05 or 0.2 mM tannic acid affected SEMIA 5019 (Table 1). Shoot N accumulation of the plants inoculated with 0.4 mM tannic acid affected SEMIA 5019, was significantly lower than that of the control (p≤0.01). Root N accumulation of the plants inoculated with 0.2 or 0.4 mM tannic acid affected cultures was significantly lower than that of the control (p≤0.001). Shoot N accumulations of the plants decreased significantly under the inoculation with 0.4 mM tannic acid affected TAL 102 (p≤0.01). The two concentrations of tannic acid separated the standard bacteroid protein profile of SEMIA 5019 by 23-27% (Fig. 1). The corresponding separation of TAL 102 was 60-93%.
| Table 1: | Effect of tannic acid-affected Bradyrhizobium elkanii SEMIA 5019 and B. japonicum TAL 102 on nodulation, plant growth and N accumulation of soybean |
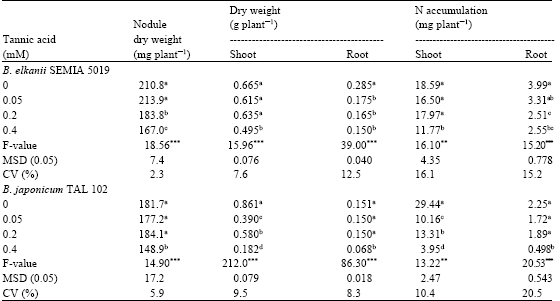 | |
| Values in the same column followed by different letter (s) indicate significant differences at p≤0.05 by Tukey’s HSD test. ***significant at p≤0.001, **significant at p≤0.01. Each value is a mean of six replicates. MSD minimum significant difference. CV Coefficient of Variation | |
| Table 2: | Effect of tannic acid on Bradyrhizobium elkanii SEMIA 5019-soybean N2 fixing symbiosis when different concentrations of tannic acid were applied to the culture medium at planting or two weeks after planting |
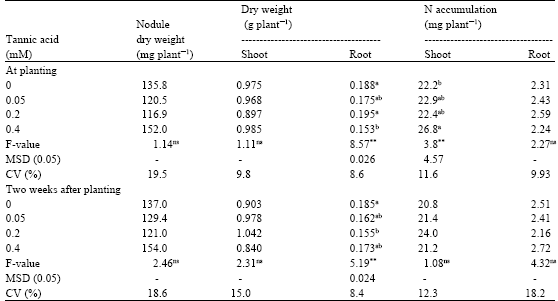 | |
| Values in the same column followed by different letter(s) indicate significant differences at p≤0.05 by Tukey’s HSD test. **significant at p≤0.01. nsnot significant at p≤0.05. Each value is a mean of six replicates. MSD minimum significant difference. CV coefficient of variation | |
Effect of Tannic Acid on Soybean-rhizobial N2 Fixing Symbiosis
At planting or two weeks after planting, tannic acid application to soybean inoculated with SEMIA 5019 did not change nodule and shoot dry weights, compared to the control (Table 2).
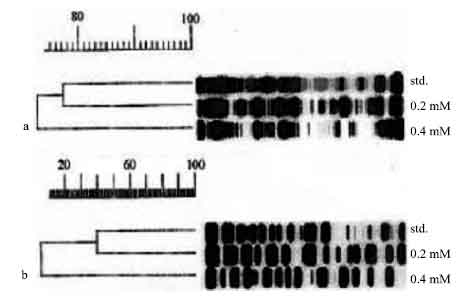 | |
| Fig. 1: | Dendrograms showing bacteroid protein profiles of soybean nodules. Proteins of (a) Bradyrhizobium elkanii SEMIA 5019 and (b) Bradyrhizobium japonicum TAL 102, as affected by two concentrations (0.2 and 0.4 mM) of tannic acid. std.-standard protein profiles of the strains in the absence of tannic acid interactions |
Root dry weight decreased whereas shoot N accumulation increased significantly, compared to the control when 0.4 mM tannic acid was applied at planting (p≤0.01), but there was no such an effect when tannic acid was applied 2 weeks after planting. Root N accumulation was not affected by tannic acid applications.
DISCUSSION
Plant growth, nodulation and N accumulation of soybean were reduced when the rhizobial strains were affected by tannic acid. Profound effects were observed with TAL 102, which were also reflected from its high alteration of bacteroid protein profiles. This indicated that the reduced performance of tannic acid-affected rhizobia could be attributed to their changes in protein biochemistry, possibly through protein-tannin complexation (Konishi and Tanaka, 1999). Among other environmental factors, this seems to be an important factor responsible for the loss of effectiveness of introduced rhizobia in a long run that has been observed in many studies (Vargas et al., 2000; Hungria et al., 2003). This makes the need for reinoculation of rhizobial inoculants for better results.
Increased shoot N accumulation of inoculated soybean under the relatively high tannic acid concentration in the culture medium at planting could have been caused by the removal of any mineral N present as impurities in the medium by tannic acid through polymerization (Northup et al., 1995), favoring nitrogenase activity.
It is thus concluded from this study that soil tannins have both favorable as well as unfavorable effects on the legume-rhizobial N2 fixing symbiosis. They reduce rhizobial performance by direct interactions whereas they enhance N2 fixing activity possibly by removing soil mineral N in the proximity of legume root system at planting.
ACKNOWLEDGMENTS
Studies on rhizobiology in this project were initiated during a Sri Lanka-Belgium collaboration on biological nitrogen fixation (1991-1997). Ms. A.P. Morawaka Arachchi, R.R. Ratnayaka and J.S. Zavahir of our project and Mr. K. Wijesekara of the institute helped with laboratory preparations and in finalizing the manuscript.
REFERENCES
- Balasubramani, G., S. Krishnaveni and S. Kannaiyan, 1992. Inhibition of Azorhizobium and Rhizobium by seed leachates. World J. Microbiol. Biotechnol., 8: 542-543.
Direct Link - Botha, W.C. and P.J. Jooste, 1992. Evaluation of normalization and compensation procedures used in the analysis of electrophoretic protein profiles for bacterial taxonomy. S. Afr. J. Sci., 88: 167-172.
Direct Link - Bremner, J.M. and M.A. Tabatabai, 1972. Use of an ammonia electrode for determination of ammonium in Kjeldhal analysis of soils. Commun. Soil Sci. Plant Anal., 3: 159-165.
Direct Link - Hagerman, A.E., C.T. Robbins, Y. Weerasuriya, T.C. Wilson and C. Mcarthur, 1992. Tannin chemistry in relation to digestion. J. Range. Manage., 45: 57-62.
Direct Link - Hungria, M., R.J. Campo and I.C. Mendes, 2003. Benefits of inoculation of the common bean (Phaseolus vulgaris) crop with efficient and competitive Rhizobium tropici strains. Biol. Fert. Soils, 39: 88-93.
Direct Link - Kogel-Knabner, I., 2002. The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter. Soil Biol. Biochem., 34: 139-162.
CrossRefDirect Link - Konishi, K. and T. Tanaka, 1999. Inhibitory effects of tannins on the NADH dehydrogenase activity of bovine heart mitochondrial complex I. Biol. Pharmocol. Bull., 22: 240-243.
Direct Link - Mila, I. and A. Scalbert, 1994. Tannin antimicrobial properties through iron deprivation: A new hypothesis. Acta Hort., 381: 749-755.
Direct Link - Northup, R.R., Z. Yu, R.A. Dahlgren and K.A. Vogt, 1995. Polyphenol controls of nitrogen release from pine litter. Nature, 377: 227-229.
Direct Link - Rice E.L. and S.K. Pancholy, 1973. Inhibition of nitrification by climax ecosystems. II: Additional evidence and possible role of tannins. Am. J. Bot., 80: 691-702.
Direct Link - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA., ISBN-13: 9780879695774, Pages: 397.
Direct Link - Vargas, M.A.T., I.C. Mendes and M. Hungria, 2000. Response of field-grown bean (Phaseolus vulgaris L.) to Rhizobium inoculation and nitrogen fertilization in two Cerrados soils. Biol. Fert. Soils, 32: 228-233.
Direct Link - Wolff, A.B., P.W. Singleton, M. Sidirelli and B.B. Bohlool, 1993. Influence of acid soil on nodulation and interstrain competitiveness in relation to tannin concentrations in seeds and roots of Phaseolus vulgaris . Soil Biol. Biochem., 25: 715-721.
Direct Link